Full Length Research Paper

ABSTRACT
Soybean is an important crop in the Democratic Republic of Congo, a country faced with high levels of war induced malnutrition but its productivity is limited by poor soil fertility coupled with low access to nitrogen mineral fertilizers. Commercial rhizobia strains introduced in 2010 failed to adapt and increase soybeans yields at desired level. We studied the performance of six indigenous rhizobia strains in enhancing soybean productivity compared to two commercial strains USDA110 and SEMIA5019. The study was carried out in the greenhouse and field of Kalambo station of International Institute of Tropical Agriculture (IITA), D. R. Congo during 2016/2017 cropping season. The treatments included: (1) N-, control without inoculation and N-fertilizer; (2) N+, non-inoculated control with 80 kg of N ha−1; and inoculated with (3) commercial strain Bradyrhizobium diazoefficiens USDA110; (4) commercial strain Bradyrhizobium elkanii SEMIA5019; (5) local strains Bradyrhizobium japonicum NAC17; (6) NAC22; (7) NAC37, (8) NAC42 (9) NAC 46 and (10) NAC78. Greenhouse and field experiments were laid out as completely randomized design and randomized complete block design respectively. The best inoculation treatments across all experiments were the indigenous strains NAC46 and NAC17 which nodulated equally or better than the commercial strain USDA 110. In the field NAC46 and NAC17 increased soybean grain yield from 2.4 to 3.3 t ha-1 and 3.4 t ha-1; indicating the increase of 68.7 and 70.8% respectively, over the commercial strain USDA110. The results demonstrated that indigenous rhizobia NAC46 and NAC17 would thus be the silver bullet to enhanced BNF and soybean yields in South Kivu province of Democratic Republic of Congo.
Key words: Inoculation, local rhizobia; soil fertility, USDA110.
INTRODUCTION
Soybean, Glycine max (L.) Merr. is an important crop worldwide and is becoming even more important and popular in South Kivu due to its potential to curb high malnutrition (Hartman et al., 2011). Soybean was introduced in Africa from Asia in the 19th century (Khojely et al., 2018) to address the need for cropping systems diversification dominated by maize (Giller et al., 2011). In these systems, soybeans enhance soil fertility through their ability to fix nitrogen (N) from atmosphere in symbiotic relationship with rhizobia bacteria through the iological Nitrogen Fixation (BNF) process (Collino et al., 2015). At the same time, this crop provides smallholders farmers the opportunity to increase their households’ income while fighting malnutrition issues because of their important nutritional value in terms of protein, amino acid and micronutrient (Arslanoglu, 2011; Xu et al., 2015).
Since the last decades, in South Kivu province of Democratic Republic of Congo, where the economy depends largely on agriculture (JeníÄek and Grófová, 2016; Maass et al., 2012), there is an increase of soybean demand due to the presence of market created by the development of livestock (Rudel et al., 2015) and industry of soybean processing (Bisimwa et al., 2012). The most common soybean based formula consumed in South Kivu province includes soy infant formula (Bahwere et al., 2016; Owino et al., 2011), soymilk, soy oil, soybean flowers, soybean biscuits, soybean spices, soybean meat, soybean bread and cakes and soybean waste industry used as animal feed (Shurtleff and Aoyagi, 2009). This crop is essentially cultivated by smallholders’ farmers and maintained by women to improve nutrition and generate income for their households, and by youth to pay costs of their education (CIALCA, 2010).
In South Kivu, farmers generally plant legumes, including the soybean without adding mineral fertilizers because they are neither available nor affordable and less economic to them (Lambrecht et al., 2016; Pypers et al., 2011a; Vanlauwe et al., 2010). Therefore, soils have been depleted due to a continuous cropping without soil replenishment as consequence of population pressure (Bashagaluke, 2015). In that case, crop yield depends upon N fixation by native rhizobia that are not always effective (Ojo et al., 2015) thus obtaining low yields, estimated at 0.5 t ha-1 (FAO, 2018).
Inoculation of soybean with appropriate, highly effective, adapted and compatible rhizobia has been stated as the most economic (Chianu et al., 2011), productive (Saturno et al., 2017) and environment friendly (Collino et al., 2015) mean to improve crop yield. Two main approaches have been pursued by research international organizations to improve soybean yield: first, promiscuous soybean cultivars were developed to nodulate freely with native rhizobia (Tefera, 2011); second, inoculation with highly effective rhizobia strains has been promoted (van Heerwaarden et al., 2018). In that line, commercial inoculants, Biofix Legume inoculants, containing Bradyrhizobium diazoefficiens USDA110 strain, was introduced among South Kivu farmers by N2 Africa program since 2010 (www.n2africa.org) and disseminated among farmers by agricultural extensions services and humanitarian organization.
From trials and farmer’s fields results, the commercial inoculants increased legume yield from 500 to 1343 kg ha-1 (van Heerwaarden et al., 2018), but still not at desired levels in certain farms and with no increase in other farmers, while the potential soybean yield is above 5000 kg ha-1 (Salvagiotti et al., 2008; Zanon et al., 2016). That low improvement was attributed to the effect of environmental and edaphic conditions on the introduced commercial strains in addition to the failure to overcome the competition barriers opposed by native rhizobia (van Heerwaarden et al., 2018).
Numerous studies in Africa have shown the presence of effective rhizobia strains among indigenous rhizobia populations (Chibeba et al., 2017; de Almeida Ribeiro et al., 2015; Musiyiwa et al., 2005). In addition, indigenous rhizobia have been described by many studies as being persistent, well adapted to local conditions and therefore can compete successfully at the expense of exotic strains for nodule occupancy and N fixation (Fening and Danso, 2002). From past results on genetic diversity on indigenous rhizobia nodulating soybeans in Africa and in South Kivu, some indigenous rhizobia clustered together with the commercial strains USDA110 (bootstrap value: 99%) showing possible relatedness of indigenous strains with this commercial strain. There is need to test these indigenous strains related to the commercial strain and identify indigenous strains suitable for South Kivu environment and edaphic conditions. We tested six indigenous rhizobia strains for their competitiveness ability to improve soybean nodulation and yield compared to the commercial strains USDA110 and SEMIA5019 in order to characterize and isolate indigenous strains with potential to be included in soybean inoculants.
MATERIALS AND METHODS
Study area
The study was carried out in South Kivu province of Democratic Republic of Congo, in the greenhouse and station field of International Institute of Tropical Agriculture (IITA), Kalambo station. South Kivu is one of the 25 provinces of Democratic Republic of Congo, located in Eastern between 1°36’ - 5° South and 26°49’- 29°20’ East and the surface is estimated to be 69,130 km2 with 3.8 million people of population with the estimated density of 91 people per km2 (Pypers et al., 2011). It is recognized as a high humid forest zone depicted by high vegetation diversity (Potapov et al., 2012) and highlands. Soils are mostly Dystric, Humic Nitisols and Humic Ferralsols (van Engelen et al., 2006; Eswaran et al., 1997). This region has a tropical climate, the Aw3 type according to Koppen classification with an average annual rainfall of 1500 mm and mean temperature of 18°C (Nash and Endfield, 2002).
Agriculture is the main activity contributing more than 50% to the GDP of this province (CIALCA, 2010). Most of the farms have the characteristics of smallholders farming systems characterized by the farm size ranging from 0.5-1 ha (Pypers et al., 2011). The main cultivated crops include cassava, common beans, maize and banana (Maass et al., 2012). Soybean crop has been promoted since 1990 to deal with high malnutrition caused by repetitive wars (Kismul et al., 2015) and since then its cultivation is increasing (FAO, 2018). This province has been the site of repetitive armed conflicts and rapes (Bartels et al., 2013).
Soils sampling and analysis
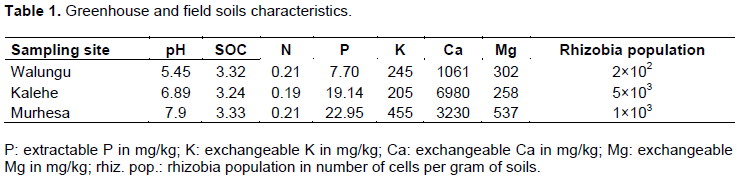
Selected field had no history of neither rhizobia inoculation nor soybean cultivation. Two weeks before sowing, twenty composite soil samples were collected from 0-20 cm depth along the field diagonal (Carter and Gregorich, 2008). Soil samples were pretreated (air dried and sieved through a 2 mm perforation size) prior for analysis. Total soil organic carbon was determined by the modified Walkley-Black (Okalebo et al., 2002), total N and available phosphorus were determined by the Kjeldahl and Olsen methods, respectively (Anderson and Ingram, 1993). Exchangeable potassium, Magnesium and Calcium were determined by Mehlich 3 method (Okalebo et al., 2002). Soil pH was determined using the 1:10 water method and measured by the seven compact, S210 Metler Toledo pH meter, after 60 min after agitation. The population of native rhizobia was determined by the plant infection technique (Somasegaran and Hoben, 1994) (Table 1).
Rhizobia culture and inoculant preparation
Six indigenous rhizobia and two commercial strains used in this study were obtained from IITA/Kalambo station rhizobiology laboratory selected based on their high effectiveness index (Ndusha, 2014) and high genetic similarity compared to the commercial strain USDA110 (Ndusha et al., in press). These rhizobia strains were streaked on Yeast Extract Mannitol broth (YMB) (Somasegaran and Hoben, 1994), incubated at 25°C until turbid until the concentration attained 109 cells ml-1. Inoculants were prepared from indigenous rhizobia cultures using sterilized peat as carrier material, incubated for two weeks and applied at the rate of 10 g kg seed-1 with 20% sugar-water (w/v) used as adhesive following the two-step inoculation method of Woomer (2011).
Trial management and experimental design
Three experiments were carried out to compare six outperformed strains from our past study (Ndusha, 2014) with two commercial strains USDA 110 and SEMIA5019: 1) Effectiveness testing in potted field soils in the greenhouse using two types of soils (Table 1), 2) on station field testing, and 3) finally assessing the growth rate and ability of utilizing different carbon sources on YMA media.
Indigenous rhizobia testing in controlled conditions
Greenhouse experiment was established in the greenhouse at Kalambo station of IITA; temperature in the greenhouse varied from 22 to 38°C. Two site soils; from Walungu and Kalehe village, were used as substrate in 3 L pot containers. The two villages were selected because they are all soybean production zones and their soils conditions are very different (Table 1). Sterilized 3 L capacity PVC pots were filled with 2.5 kg of soil and covered with a sterile plastic plate with limited access to limit contamination. Soybean seeds were surface sterilized using the pre described procedure (Somasegaran and Hoben, 1994), pre-germinated in agar plates; and 3 seeds per pot were sewed, thinned to 2 after emergence for appropriate spacing. Seeds were inoculated with 1 ml of broth pre cultured (described in section 2.3). For the mineral N control, urea was applied at a rate of 80 kg ha-1 (Pypers et al., 2011). Watering was done regularly at the frequency of 3 per week adjusted according to plant needs. After 7 weeks, at early flowering, plants were harvested; nodules counted, weighted and shoot weight determined by weighing after oven dried at 70°C for 48 h. A Completely Randomized Design consisting of 10 treatments including 6 indigenous rhizobia and 2 commercial strains (SEMIA5019 and USDA110), and non-inoculated pot with (N+) and without mineral N (N-), with 3 replicates was established. Promiscuous soybean (SB24) was used as the test crop, selected for their high adoption among farmers (Walangululu et al., 2014).
Indigenous rhizobia testing in field condition
A field experiment was established in the station field of IITA Kalambo located in Murhesa during 2015-2016 long rains (September to January). Soils characteristics of field were determined (Table 1). Six indigenous rhizobia strains were compared to 2 commercial strains USDA110 and SEMIA5019 on promiscuous soybean variety SB24. The experiment was laid out as Randomized Complete Block design with 3 replicates. The treatments included: (1) N-, control without inoculation and N-fertilizer; (2) N+, non-inoculated control with 80 kg of N ha−1; and inoculated with (3) commercial strain B. diazoefficiens USDA110; (4) commercial strain Bradyrhizobium elkanii SEMIA5019; (5) local strains Bradyrhizobium japonicum NAC17; (6) NAC22; (7) NAC37, (8) NAC42 (9) NAC 46 and (10) NAC78. Each plot measured 6 m x 4 m, seeds were planted in rows 45 cm apart and at 5 cm intervals. To avoid cross contamination, plots were separated by four non-inoculated lines. Legume inoculants were prepared from isolates subsequently described in the paper.
The trial management was done according to known farmer’s practice; weeding as per need before the canopy closure. The intensity of green color in leaves was measured at different growth stages using a chlorophyll meter (Dey et al., 2016). Plants were assessed for nodulation at flowering stage, seven weeks after planting. Plants were carefully uprooted, roots washed, nodules counted, oven-dried for 48 h at 70°C and dry weight recorded. Soybean grains were harvested at maturity (4 months), dried and dry weight recorded.
Data analysis
Data were subjected to analysis of variance (ANOVA) using the software R version 3.5.1. When differences between treatments were detected, Tukey test was used to compare means at p >0.05 level of significance. Relative effectiveness (RE) was determined by dividing the shoot dry weight of treatment over that of the N+ treatment, in the same block (Chibeba et al., 2018).
RESULTS
Phylogenetic relationship between indigenous strains and commercial strains
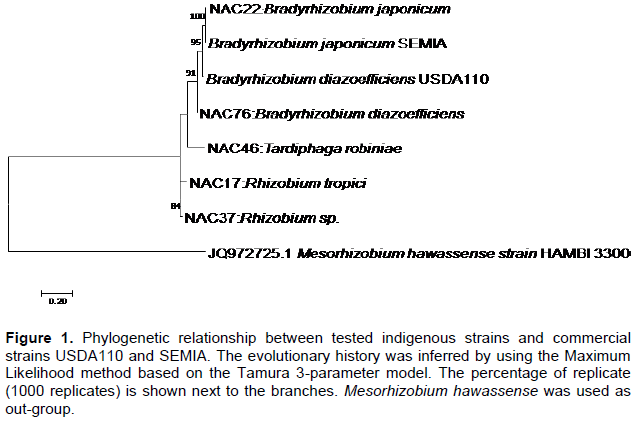
Indigenous rhizobia isolated from Congo phylogeny was determined based on the housekeeping gene gnlII. The tested isolates and commercial strain USDA 110 tended to cluster together with bootstrap value of over 90% (Figure 1).
Nodulation and shoot dry weight of indigenous rhizobia recorded in the greenhouse
Nodules number reported in the greenhouse varied significantly among strains (p=0.0001) and between the two soils types (p=0.0203). The recorded nodules number varied from 3 nodules to 21 nodules per plant in Walungu soil while it varied between 1 to 36 nodules per plant in Kalehe soil. The highest nodules number in Walungu was recorded by the indigenous strain NAC46 and NAC76 (±21 nodules per plant), followed by commercial strains (USDA110 and SEMIA 5019) and indigenous strains (NAC22, NAC17 and NAC37). The lowest number of nodules in Walungu site soils was recorded by the treatment without inoculation but with mineral N fertilizer (N+), where recorded nodules number averaged only 3 nodules per plant (Table 2). The highest number of nodules in Kalehe soil was recorded with the same indigenous strains NAC46 and the control (N-) (average 34 nodules per plant), followed by NAC76 (21 nodules per plant) (Table 2). The lowest nodules number per plant was also reported with the treatment N+, where the nodule number averaged only 1.
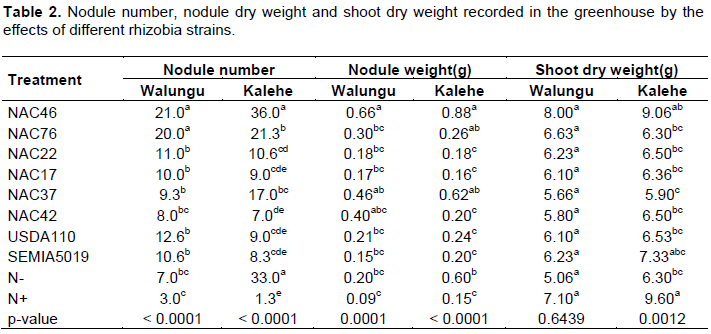
The nodules dry weight (DW) recorded in the greenhouse experiment also varied between site soils and among strains (p=0.0001). The highest nodules dry weight was recorded in Kalehe soil (average 0.373 g per plant DW) while the lowest nodules weight was recorded in Walungu soil (average 0.284 g per plant DW). The inoculation with indigenous strains NAC46 produced the highest nodules weight followed by NAC37 and NAC76 in Walungu soil (Table 2). The lowest nodules weight was recorded with N+ control for both soils.
Shoot dry weight variation (p=0.0012) was recorded only in the Kalehe soils while in Walungu no difference was recorded among the inoculated strains. In Kalehe soils, the highest shoot weight was recorded by N+ control (9.6 g plant DW), followed by indigenous strain NAC46, and by the commercial strain SEMIA5019. The lowest shoot dry weight in Kalehe soils was recorded by the indigenous strain NAC37 (5.9 g plant DW) (Table 2).
Nodule number, nodule dry weight, shoot dry weight, leaf greenness, plant height and crop yield recorded in the field study
In the field, all treatments produced nodules but their number varied greatly across treatments (p< 0.0001). The nodules number varied from 30 to 69 nodules per plant. Even the non-inoculated and not fertilized (N- and N+) control plants nodulated abundantly (average 40 nodules per plant). The highest nodule number was recorded by the treatments of both indigenous and commercial strains, which did not differ among them, except for NAC42, with a lower number of nodules (Table 3).
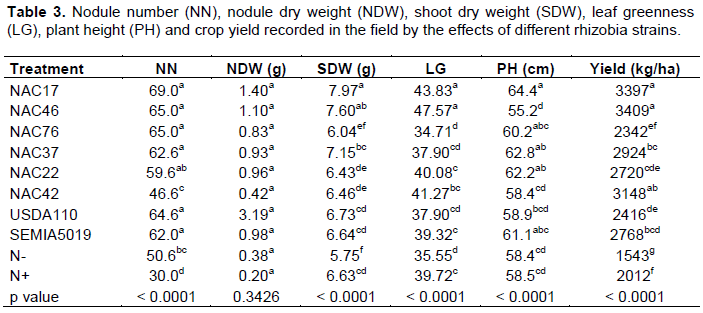
Nodules dry weight did not vary with the treatments but shoot dry weight significantly varied among rhizobia strains (p< 0.0001). The highest biomass was recorded with the indigenous strain NAC17 (8.0 g plant DW), followed by NAC46, even higher than the commercial strains USDA110 and SEMIA5019. The lowest shoot dry weight was recorded by the treatment N- control (5.8 g plant DW) (Table 3).
The plant leaf greenness also varied among treatments (p< 0.0001). The highest intensity of green color measured on leaves was recorded by the indigenous strains NAC17 and NAC46 (Table 3). The lowest green color intensity was recorded by the control N- and the indigenous strain NAC76 (about 55) (Table 3).
Plant height also varied significantly across treatments (p< 0.0001). The highest plant height was recorded by the indigenous rhizobia strain NAC17 (64 cm), followed NAC22 and NAC37. The commercial strain SEMIA5019 took the third place while USDA 110 took the fourth position. The lowest treatment in terms of plant height was the treatment NAC46 (55 cm) (Table 3). Grain yield also varied across treatments. Yield improvement was recorded by the indigenous strains NAC17 and NAC46 that yielded 1.4 fold than the commercial strain USDA110 and 1.6 fold than the application of N (N+). The lowest grain yield was recorded by the treatment N-, followed by N+ (Table 4).
Relative effectiveness of indigenous rhizobia strains compare to the reference strains
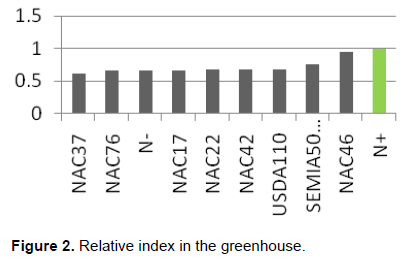
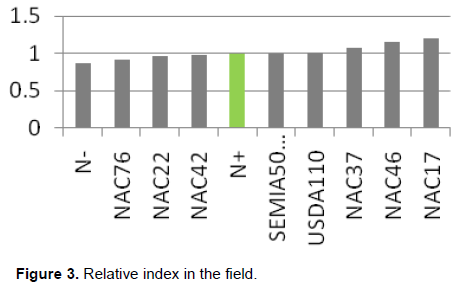
In the greenhouse, the effectiveness of all tested isolates did not exceed the reference treatment N+ but the indigenous rhizobia NAC46 had a relative index higher than the commercial strains USDA110 and SEMIA 5019 (Figure 2). However, in the field, the relative effective index by the 3 indigenous rhizobia strains (NAC17, NAC46 and NAC37) exceeded both commercial strains and control with N (N+) (Figure 3.)
DISCUSSION
Effectiveness of indigenous strains under controlled environment
In the greenhouse (controlled environment), there was significant differences in nodulation among treatments in the two soils highlighting the need to inoculate the soils of the two sites. These differences may be explained by the low abundance or less effectiveness of native rhizobia population in the Walungu and Kalehe soils (Table 1). Sanginga and Okogun (2003) and Osunde et al. (2003) stated that inoculation responses are more likely to occur when there are less than 103 cells of indigenous or naturalized rhizobia per gram of soil or when the native rhizobia are less effective. The same results were found by Koskey et al. (2017) in their study on potential of native rhizobia in enhancing N fixation and crop yield of climbing beans in contrasting environments of Kenya, the yield improvement and nodulation of introduced rhizobia strains depended largely on the number of viable native rhizobia in the soil.
There was significant difference in nodulation between the two sites soils, higher nodules number was produced in Kalehe soils (Table 2). This is because of the differences in soils conditions (Table 1). Nodulation depends on a number of soils factors, especially the soil pH (Lapinskas, 2007), P availability and the indigenous rhizobia abundance and effectiveness (Sanginga et al., 1996; Singleton and Tavares, 1986; Slattery, 2004; Thies et al., 1991). Kalehe soil has better conditions for growth and survival of rhizobia bacteria namely the neutral pH and higher P content compared to Walungu soil (Table 1). This result is in concordance with other authors, for example Gyogluu et al. (2016) who assessed the symbiotic response of soybeans to inoculation by different B. japonicum strains at 3 experimental sites in Mozambique. They found response variation depending on different sites and suggested that there are specific effects of sites on nodulation and dry matter improvement by rhizobia. This observation is also in agreement with Boucho et al. (2019); they found that the response to inoculation is highly affected by soils conditions and for their case phosphorus availability in the soil promote nodulation and biomass. Therefore, this study suggests the improvement of soils conditions prior to inoculation such as liming where soil pH is low and phosphorus application should be considered for maximization of inoculation response.
A significant difference in shoot dry weight (biomass) improvement in the greenhouse was observed only in Kalehe soils. The non response in Walungu soils may be explained in terms of the low pH and low P levels, which limited the process of nitrogen fixation despite the presence of nodules. The same observation was done by a study conducted in Argentine by Collino et al. (2015); they observed variation of BNF depending on crop, soil and meteorological factors. This study also revealed differences among strains in both nodulation and shoot dry weight improvement which is primarily due to their genetic differences. Other studies conducted in Africa have similarly reported consistent variation in symbiotic effectiveness among indigenous rhizobia strains (Abaidoo et al., 2007; Chibeba et al., 2017, 2018) and consequently this study further contributes to the evidence that effective rhizobia do occur in African soils.
Effectiveness of indigenous strains under field conditions
In the field, nodules were observed on all treatments including the non inoculated control without nitrogen N-; the non inoculated control produced even higher nodules number compare to the inoculated plots. This is due to the presence in soils of natives and compatible rhizobia. Many studies have demonstrated that effective rhizobia can be presented in some soils but they are less efficient (Abaidoo et al., 2007; Gyogluu et al., 2016; Jaiswal and Dakora, 2019; Wongphatcharachai et al., 2015). These less efficient indigenous rhizobia strains can be even more competitive than the inoculants and occupy a significant portion of the nodules, reducing the impact of the introduced inoculant strain on improving BNF (Batista et al., 2015). The same observations were made by Irisarri et al. (2019) who observed a higher nodules occupation by native rhizobia compared to introduced strains. This has important practical implication for agriculture to the effect that selection of rhizobia for improving BNF and legume productivity must be done among native population, and be provided in higher concentration through inoculants. Furthermore, the host plant show preference on native rhizobia compared to exotic strains (Osunde et al., 2003).
Only the N+ control produced very few nodules. This is attributable to the fact that the presence of mineral nitrogen inhibits the biological nitrogen fixation by inhibiting the nodules formation and nitrogenase activity. This result is in agreement with a study conducted by Ulzen et al. (2016) who reported very few nodules with the application of 100 kg of nitrogen. In addition, many authors have stated that N doses as low as 20-40 kg of N/ha may highly decrease nodulation and BNF, with no benefits to yield (Hungria et al., 2005, 2006; Saturno et al., 2017).
The differences in nodulation among tested strains in response to inoculation may be attributed to the fact that natives or naturalized rhizobia were less effective in one hand and on the other hand introduced strains were highly effective. This results are in line with those of Osunde et al. (2003) who tested the nodulation of two soybean promiscuous varieties by introduced elite rhizobia and indigenous rhizobia. The introduced rhizobia through inoculants were less competitive but highly effective compared to native rhizobia. The differences in nodulation may be ascribed to their genetic makeup expressed through symbiotic efficiency, competitiveness for nodule occupancy, compatibility with the host plant and adaptive ability to soil stress conditions (Tas et al., 1996). The indigenous rhizobia NAC17 and NAC46 produced higher number of nodules compared to the commercial strain USDA110 suggesting that these strains had higher symbiotic efficiency and higher adaptation ability to local conditions compared to the commercial strains.
The increase of leaves green color noted in the native strains compared to the control may be because of nitrogen being the major constituent of chlorophyl that confers green color to the plants. Inoculation improves nitrogen content in leaves and thus promotes the formation of chlorophyll which is also important for photosynthesis (Sinclair, 2004; Hakeem et al., 2012). The same results were observed by Abaidoo et al. (2007) who classified rhizobia isolates tested into four symbiotic phenotypic groups based on green color intensity. The less effective group was composed by isolates that recorded lower green color intensity on leaves of soybean genotypes.
In the field, the significant differences of shoot dry weight were observed among treatments as result of enhanced nodulation. Nitrogen is the component responsible for vegetative development; it has been demonstrated that up to 80% of the above-ground N accumulation in soybean is due to N fixation by rhizobia (Hungria et al., 2006). The N- control produced a higher number of nodules but lower shoot dry weight. This is as a result of the native rhizobia being less effective (Osunde et al., 2003). The same observation has been made by Chibeba et al. (2018) who observed that an appreciable proportion of rhizobia population in Mozambican soils was composed of ineffective rhizobia. The N+ control produced high shoot dry weight; this is mainly as a result of the mineral N being absorbed by the plant at early stages compared to the fixed N and thus improved vegetative formation (Saturno et al., 2017). This is in agreement with the findings of Kinugasa et al. (2012) who found that higher biomass production did not result in grain yield increase.
This study demonstrated significant differences in yields among treatments. There was yield improvement with inoculated plots compare to the plots where N had been applied and N-control. The indigenous strains NAC46 and NAC17 increased yields by 1.7 and 1.6 fold respectively compared to the N+ control, 2.2 folds compared to the N-control and 1.4 and 1.3 fold compared to the commercial strain USDA110.These yield gains are within the 3.2–14.5% interval of inoculation yield benefit reported in Brazil (Hungria et al., 2006) and in Mozambique (Chibeba et al., 2018). This study has further confirmed the findings of Chibeba et al. (2017, 2018) and Hungria et al., (2005, 2006) that BNF is the most efficient way of improving soybean productivity.
CONCLUSION
The best strains across all the experiments are NAC17 and NAC46 with average yields gain 60-70% over commercial strains and controls. These results suggest that these indigenous strains hold the best potential as commercial inoculants in South Kivu soils conditions. USDA 110 and SEMIA 5019 are also effective but it is preferable to use adapted and competitive strains. Therefore, the native strains are likely to adapt well not only in South Kivu, but also in other countries with similar agro-climatic conditions.
ACKNOWLEDGEMENT
This study is a part of PhD at University of Nairobi undertaken by the first author and was funded by RUFORUM (grant number: RU/2016/GTA/DRG/004). The N2 Africa project is acknowledged for rhizobia strains collection and maintenance. The Organization of Women for Science in Developing World (OWSD) is thanked for scholarship to the first author. The Université Evangelique en Afrique Through the partnership with Pain pour le Monde Organization is appreciated for support to the first author.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abaidoo RC, Keyser, Singleton HH, Dashiell, Sanginga KE (2007). Population size, distribution, and symbiotic characteristics of indigenous Bradyrhizobium spp. that nodulate TGx soybean genotypes in Africa. Applied Soil Ecology 35:57-67. Crossref |
||||
|
Arslanoglu F (2011). Effect of genotype and environment interaction on oil and protein content of soybean (Glycine max (L.) Merrill) seed. African Journal Biotechnology 10(80):8409-18417. Crossref |
||||
|
Bahwere P, Balaluka B, Wells JC, Mbiribindi CN, Sadler K, Akomo P, Dramaix-Wilmet M, Collins S (2016). Cereals and pulse-based ready-to-use therapeutic food as an alternative to the standard milk- and peanut paste-based formulation for treating severe acute malnutrition: a non inferiority, individually randomized controlled efficacy clinical trial. American Journal of Clinical Nutrition 103:1145-1161. Crossref |
||||
|
Bartels S, Kelly J, Scott J, Leaning J, Mukwege D, Joyce N, VanRooyen M (2013). Militarized Sexual Violence in South Kivu, Democratic Republic of Congo. Journal of Interpersonal Violence 28(2)340-358. Crossref |
||||
| Bashagaluke BJ (2015). Application of infrared technique in soil properties' characterization in South Kivu province of DR Congo. African Journal of Food Science and Technology 6(2):58-67. | ||||
|
Bisimwa G, Owino VO, Bahwere P, Dramaix M, Donnen P, Dibari F, Collins S (2012). Randomized controlled trial of the effectiveness of a soybean-maize-sorghum-based ready-to-use complementary food paste on infant growth in South Kivu, Democratic Republic of Congo. American Journal of Clinical Nutrition 95:1157-1164. Crossref |
||||
| Carter MR, Gregorich EG (Eds) (2008). Soil sampling and methods of analysis, 2nd edition Canadian Society of Soil Science ; CRC Press, [Pinawa, Manitoba] : Boca Raton, FL. | ||||
|
Boucho AC, Carranca C, Redondo R, Calouro F, Madeira M (2019). Biomass, nodulation and N 2 fixing response by subclover and pink serradela to phosphorus fertilization. Archives of Agronomy and Soil Science 65(10):1431-1445. Crossref |
||||
|
Chianu Jonas N, Nkonya EM, Mairura FS, Chianu Justina N, Akinnifesi FK (2011). Biological nitrogen fixation and socioeconomic factors for legume production in sub-Saharan Africa: a review. Agronomy for Sustainable Development 31(1):139-154. Crossref |
||||
|
Chibeba AM, Kyei-Boahen S, Guimarães MF, Nogueira MA, Hungria M (2018). Feasibility of transference of inoculation-related technologies: A case study of evaluation of soybean rhizobial strains under the agro-climatic conditions of Brazil and Mozambique. Agriculture, Ecosystems and Environment 261:230-240. Crossref |
||||
|
Chibeba AM, Kyei-Boahen S, Guimarães M, Nogueira MA, Hungria M (2017). Isolation, characterization and selection of indigenous Bradyrhizobium strains with outstanding symbiotic performance to increase soybean yields in Mozambique. Agriculture, ecosystems and environment 246:291-305. Crossref |
||||
| CIALCA (2010). Banana Marketing in Rwanda, Burundi and South Kivu CIALCA Project Survey Report 40. | ||||
|
Collino DJ, Salvagiotti F, Perticari A, Piccinetti C, Ovando G, Urquiaga S, Racca RW (2015). Biological nitrogen fixation in soybean in Argentina: relationships with crop, soil, and meteorological factors. Plant Soil 392:239-252. Crossref |
||||
|
de Almeida Ribeiro PR, dos Santos JV, Martins da Costa E, Lebbe L, Silva Assis E, Oliveira Louzada M, Azarias Guimarães A, Willems A, de Souza Moreira FM (2015). Symbiotic efficiency and genetic diversity of soybean bradyrhizobia in Brazilian soils. Agriculture Ecosystems and Environment 212:85-93. Crossref |
||||
|
Dey AK, Sharma M, Meshram MR (2016). An Analysis of Leaf Chlorophyll Measurement Method Using Chlorophyll Meter and Image Processing Technique. Procedia Computer Science 85:286-292. Crossref |
||||
| FAO, FAOSTAT (2018). Rome. | ||||
|
Fening JO, Danso SKA (2002). Variation in symbiotic effectiveness of cowpea bradyrhizobia indigenous to Ghanaian soils. Applied Soil Ecology 21:23-29. Crossref |
||||
|
Giller KE, Murwira MS, Dhliwayo DKC, Mafongoya PL, Mpepereki S (2011). Soyabeans and sustainable agriculture in southern International Journal of Agricultural Sustainability 9(1):50-58. Crossref |
||||
|
Gyogluu C, Boahen SK, Dakora FD (2016). Response of promiscuous-nodulating soybean (Glycine max L. Merr.) genotypes to Bradyrhizobium inoculation at three field sites in Mozambique. Symbiosis 69:81-88. Crossref |
||||
|
Hakeem KR, Chandna R, Ahmad A, Iqbal M (2012). Reactive Nitrogen Inflows and Nitrogen Use Efficiency in Agriculture: An Environment Perspective, in: Ahmad P, Prasad MNV (Eds.), Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change. Springer New York, New York pp. 217-232. Crossref |
||||
|
Hartman GL, West ED, Herman TK (2011). Crops that feed the World 2. Soybean-worldwide production, use, and constraints caused by pathogens and pests. Food Security 3:5-17. Crossref |
||||
|
Hungria M, Franchini JC, Campo RJ, Crispino CC, Moraes JZ, Sibaldelli RNR, Mendes IC, Arihara J (2006). Nitrogen nutrition of soybean in Brazil: Contributions of biological N 2 fixation and N fertilizer to grain yield. Canadian Journal of Plant Science 86(4):927-939. Crossref |
||||
|
Hungria M, Franchini JC, Campo RJ, Graham PH (2005). The Importance of Nitrogen Fixation to Soybean Cropping in South America, in: Werner D, Newton WE (Eds.), Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment. Springer-Verlag, Berlin/Heidelberg pp. 25-42. Crossref |
||||
|
Irisarri P, Cardozo G, Tartaglia C, Reyno R, Gutiérrez P, Lattanzi FA, Rebuffo M, Monza J (2019). Selection of Competitive and Efficient Rhizobia Strains for White Clover. Frontiers in microbiology 10:768. Crossref |
||||
|
Jaiswal SK, Dakora FD (2019). Widespread Distribution of Highly Adapted Bradyrhizobium Species Nodulating Diverse Legumes in Africa. Frontiers in Microbiology P 10. Crossref |
||||
|
JeníÄek V, Grófová Š (2016). The least developed countries & ndash; the case of the Congo D.R. Agricultural Economics-Zemedelska Ekonomika 61:135-148. Crossref |
||||
|
Khojely DM, Ibrahim SE, Sapey E, Han T (2018). History, current status, and prospects of soybean production and research in sub-Saharan Africa. Crop Journal 6:226-235. Crossref |
||||
|
Kinugasa T, Sato T, Oikawa S, Hirose T (2012). Demand and supply of N in seed production of soybean (Glycine max) at different N fertilization levels after flowering. Journal of Plant Research 125:275-281. Crossref |
||||
|
Kismul H, Hatløy A, Andersen P, Mapatano M, Van den Broeck J, Moland KM (2015). The social context of severe child malnutrition: a qualitative household case study from a rural area of the Democratic Republic of Congo. International Journal of Equity Health 14:47. Crossref |
||||
|
Koskey G, Mburu SW, Njeru EM, Kimiti JM, Ombori O, Maingi JM (2017). Potential of Native Rhizobia in Enhancing Nitrogen Fixation and Yields of Climbing Beans (Phaseolus vulgaris L.) in Contrasting Environments of Eastern Kenya. Frontier Plant Science P 8. Crossref |
||||
|
Lambrecht I, Vanlauwe B, Maertens M (2016). Integrated soil fertility management: from concept to practice in Eastern DR Congo. International Journal of Agricultural Sustainance 14:100-118. Crossref |
||||
|
Lapinskas EB (2007). The effect of acidity on the distribution and symbiotic efficiency of rhizobia in Lithuanian soils. Eurasian Soil Science 40:419-425. Crossref |
||||
|
Maass BL, Katunga Musale D, Chiuri WL, Gassner A, Peters M (2012). Challenges and opportunities for smallholder livestock production in post-conflict South Kivu, eastern DR Congo. Tropical Animal Health Production 44:1221-1232. Crossref |
||||
|
Miles AA, Misra SS,Irwin JO (1938). The estimation of the bactericidal power of the blood. Journal of Hygiene 38:732-749. Crossref |
||||
|
Musiyiwa K, Mpepereki S, Giller KE (2005). Symbiotic effectiveness and host ranges of indigenous rhizobia nodulating promiscuous soyabean varieties in Zimbabwean soils. Soil Biology and Biochemistry 37(6):1169-1176. Crossref |
||||
|
Nash DJ, Endfield GH (2002). A 19th century climate chronology for the Kalahari region of central southern Africa derived from missionary correspondence. International Journal of Climatology 22:821-841. Crossref |
||||
| Ndusha BN (2014). Effectiveness of rhizobia strains isolated from South Kivu sois on nodulation and growth of soybeans. Msc Thesis University. Nairobi 111. | ||||
|
Ojo A, Dare MO, Fagbola O, Babalola O (2015). Variations in infectivity of indigenous rhizobial isolates of some soils in the rainforest zone of Nigeria. Archives of Agronomy and Soil Science 61(3):371-380. Crossref |
||||
| Okalebo JR, Gathua KW, Woomer P (2002). Laboratory Methods of Soil and Plant Analysis: A Working Manual, Second Edition. TSBF, Nairobi, Kenya. | ||||
| Okereke GU, Okey OC (2007). Survival of cowpea Bradyrhizobia in carrier material and inoculation response in soil. African Crops Science Conference Proceeding 8:1183-1186. | ||||
|
Osunde AO, Gwam S, Bala A, Sanginga N, Okogun JA (2003). Responses to rhizobial inoculation by two promiscuous soybean cultivars in soils of the Southern Guinea savanna zone of Nigeria. Biology of Fertilized Soils pp. 274-279. Crossref |
||||
|
Owino VO, Bahwere P, Bisimwa G, Mwangi CM, Collins S (2011). Breast-milk intake of 9-10-mo-old rural infants given a ready-to-use complementary food in South Kivu, Democratic Republic of Congo. American Journal of Clinical Nutrition 93:1300-1304. Crossref |
||||
|
Potapov PV, Turubanova SA, Hansen MC, Adusei B, Broich M, Altstatt A, Mane L, Justice CO (2012). Quantifying forest cover loss in Democratic Republic of the Congo, 2000-2010, with Landsat ETM+ data. Remote Sensing of Environment 122:106-116. Crossref |
||||
|
Pypers P, Sanginga JM, Kasereka B, Walangululu M, Vanlauwe B (2011). Increased productivity through integrated soil fertility management in cassava-legume intercropping systems in the highlands of Sud-Kivu, DR Congo. Field Crops Research 120:76-85. Crossref |
||||
|
Ronner E, Franke AC, Vanlauwe B, Dianda M, Edeh E, Ukem B, Bala A, van Heerwaarden J, Giller KE (2016). Understanding variability in soybean yield and response to P-fertilizer and rhizobium inoculants on farmers' fields in northern Nigeria. Field Crops Research 186:133-145. Crossref |
||||
|
Salvagiotti F, Cassman KG, Specht JE, Walters DT, Weiss A, Dobermann A (2008). Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Research 108:1-13. Crossref |
||||
|
Sanginga N, Abaidoo R, Dashiell K, Carsky RJ, Okogun A (1996). Persistence and effectiveness of rhizobia nodulating promiscuous soybeans in moist savanna zones of Nigeria. Applied Soil Ecology 3:215-224. Crossref |
||||
|
Sanginga N, Okogun JA (2003). Can introduced and indigenous rhizobial strains compete for nodule formation by promiscuous soybean in the moist savanna agroecological zone of Nigeria? Biology of Fertilized Soils 38:26-31. Crossref |
||||
|
Saturno DF, Cerezini P, Moreira da Silva P, Oliveira AB, de Oliveira MCN, de Hungria M, Nogueira MA (2017). Mineral nitrogen impairs the biological nitrogen fixation in soybean of determinate and indeterminate growth types. Journal of Plant Nutrition 40:1690-1701. Crossref |
||||
| Shurtleff W, Aoyagi A (2009). History of soybeans and soyfoods in Africa (1857-2009): extensively annotated bibliography and sourcebook. Soy info Center, Lafayette, CA. | ||||
| Sinclair TR (2004). Improved carbon and nitrogen assimilation for increased yield. In:H.R. Boerma and J.E. specht (eds), Soybeans: Improvement, Production, and Uses. American Society of Agronomy, Madison WI. pp. 537-568. | ||||
| Singh B, Kaur R, Singh K (2008). Characterization of Rhizobium strain isolated from the roots of Trigonelle foenumgraceum (fenugreek). African Journal of Biotechnology 7:3671-3676. | ||||
| Singleton PW, Tavares JW (1986). Inoculation Response of Legumes in Relation to the Number and Effectiveness of Indigenous Rhizobium Populations. Applied Environmental Microbiology 51:6. | ||||
|
Slattery J (2004). Effects of resident rhizobial communities and soil type on the effective nodulation of pulse legumes. Soil Biology and Biochemistry 36:1339-1346. Crossref |
||||
|
Somasegaran P, Hoben HJ (1994). Handbook for Rhizobia. Springer New York, New York. Crossref |
||||
| Tas EV, Leinonen P, Saano A, Piippola S, Hakola S (1996). Assessment of Competitiveness of Rhizobia Infecting Galega orientalis on the Basis of Plant Yield, Nodulation, and Strain Identiï¬cation by Antibiotic Resistance and PCR. Applied Environmental Microbiology 62:7. | ||||
|
Tefera H (2011). Breeding for Promiscuous Soybeans at IITA, in: Sudari A. (Ed.), Soybean - Molecular Aspects of Breeding. InTech. Crossref |
||||
| Thies JE, Singleton PW, Bohlool BB (1991). Influence of the Size of Indigenous Rhizobial Populations on Establishment and Symbiotic Performance of Introduced Rhizobia on Field-Grown Legumest. Applied Environmental Microbiology 57:10. | ||||
|
Ulzen J, Abaidoo RC, Mensah NE, Masso C, AbdelGadir AH (2016). Bradyrhizobium Inoculants Enhance Grain Yields of Soybean and Cowpea in Northern Ghana. Frontier of Plant Science P 7. Crossref |
||||
| van Engelen V, Verdoodt A, Dijkshoorn K, Ranst EV (2006). Soil and Terrain Database of Central Africa - DR of Congo, Burundi and Rwanda (SOTERCAF, version 1.0). ISRIC FAO, Report 2006/07, 28. | ||||
|
Van Heerwaarden J, Baijukya F, Kyei-Boahen S, Adjei-Nsiah S, Ebanyat P, Kamai N, Wolde-meskel E, Kanampiu F, Vanlauwe B, Giller K (2018). Soyabean response to rhizobium inoculation across sub-Saharan Africa: Patterns of variation and the role of promiscuity. Agriculture, Ecosystems and Environment 261:211-218. Crossref |
||||
|
Vanlauwe B, Bationo A, Chianu J, Giller KE, Merckx R, Mokwunye U, Ohiokpehai O, Pypers P, Tabo R, Shepherd KD, Smaling EMA, Woomer PL, Sanginga N (2010). Integrated soil fertility management Operational definition and consequences for implementation and dissemination. Outlook Agriculture 39:17-24. Crossref |
||||
| Walangululu MJ, Shukuru BL, Bamuleke KD, Bashagaluke BJ, Anjelani AA, Baijukya F (2014). Response of introduced soybeans varieties to inoculation with rhizobium in Sud Kivu province of Democratic Republic of Congo. 4th Ruforum Biennial Regional conference proceedings, Maputo pp. 273-279. | ||||
|
Wongphatcharachai M, Staley C, Wang P, Moncada KM, Sheaffer CC, Sadowsky MJ (2015). Predominant populations of indigenous soybean-nodulating Bradyrhizobium japonicum strains obtained from organic farming systems in Minnesota. Journal of Applied Microbiology 118:1152-1164. Crossref |
||||
| Woomer PL, Karanja N, Kisamuli SM, Murwira M, Bala A (2011). A revised manual for rhizobium methods and standard protocols available on the project website, www.N2Africa.org, 69 p. | ||||
|
Xu XP, Liu H, Tian L, Dong XB, Shen SH, Qu LQ (2015). Integrated and comparative proteomics of high-oil and high-protein soybean seeds. Food Chemistry 172:105-116. Crossref |
||||
|
Zanon AJ, Streck NA, Grassini P (2016). Climate and Management Factors Influence Soybean Yield Potential in a Subtropical Environment. Agronomy Journal 108(4):1447-1454. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


