Full Length Research Paper

ABSTRACT
The growth of Cyperus sesquiflorus was characterized in an experiment with 4 replications and 15 treatments (0, 6, 17, 22, 28, 34, 40, 45, 49, 55, 61, 69, 76, 83 and 90 days after transplanting - DAT). Greater leaf length, diameter of the culm, dry matter, and biomass partitioning measurements were made. Alive leaves, inflorescences, alive and dead tillers were counted. The Tukey test at 5% was used for analysis. Greater leaf length was obtained 28 DAT. The treatments 0, 6, 17, 22, 28 and 34 DAT provided the largest diameters. The number of leaves decreased over time, with 0, 6, 17, 22, 28 and 34 DAT being the best (6 to 8). The length of the root system reached 15 cm at 0 to 34 DAT, with no difference from 22 and 28 DAT. The highest number of inflorescences was obtained at 90 DAT, which also provided the highest number of tillers and total dry matter. Biomass partitioning to the underground part was greater in 6, 76 and 90 DAT and the only treatment in which biomass partitioning to the aerial part was greater was 61 DAT. Growth occurred up to 34 days, and decreased due to shading, tillering, flowering, and limitations imposed by the container, with exception of the number of inflorescences and new tillers. Thus, our study provides new information on the behavior of C. sesquiflorus that will contribute to upcoming research on the growth, control, and essential oil production of this species.
Key words: Cyperaceae, Kyllinga odorata Vahl, weeds, medicinal, clonal propagation.
INTRODUCTION
Knowledge of the bio-ecological aspect of weeds, with elucidation of growth patterns and behavior of the species, is essential for the development of control and management strategies (Soares et al., 2017). The irreversible increase in plant size, mass or volume, especially of protoplasmic material, is called growth (Reis, 1978). Plant growth is the result of complex physical-biochemical interactions and photosynthetic production and storage which, when analyzed, leads to the understanding of the physiological processes involved in the behavior of plants under different environmental or growing conditions (Benincasa, 1988).
The family Cyperaceae is composed of species that frequently form weed communities in agricultural crops (Nunes et al., 2018; Almeida et al., 2018). The growth in species of the genus Cyperus L. has been investigated by Brighenti et al. (1997) studied Cyperus rotundus L. cultivated in pipes; Rodiyati and Nakagoshi (2003) studied Cyperus brevifolius (Rottb.) Endl. ex Hassk. in its natural habitat; Bryson and Carter (2010) studied Cyperus fuscus L. in greenhouse conditions and natural habitat; and Matos et al. (2008) studied Cyperus esculentus L. cultivated in beds.
Cyperus sesquiflorus (Torr.) Mattf. & Kük. - valid botanical name for Kyllinga odorata Vahl - is a pantropical, perennial, terrestrial herbaceous plant species native to Brazil that has a cosmopolitan distribution throughout the country (Alves et al., 2015; Simpson and Inglis, 2001; GBIF, 2019; Bryson and Carter, 2008). It is considered an invasive species in crops, especially rice, easily found with reproductive structures during all months of the year (Trevisan et al., 2007; Machado Filho et al., 2015). In warm climates, C. sesquiflorus is described as having a short-lived annual or perennial habit (Carter, 2005; Bryson and Carter, 2008). It presents a caespitose growth form, rhizomes with short internodes, stramineous inflorescences, glabrous glume keels, and persistent old leaves adhered to the base of mature plants (Trevisan et al., 2007).
C. sesquiflorus produces an essential oil with medicinal properties (Gonzáles et al., 2009; Novoa et al., 2012; Peckolt and Peckolt, 2016). It is popularly named “fragrant grass” (“capim cheiroso” in Portuguese) due to its aromatic content. In Brazil, the oil from the secondary metabolism of Cyperus articulatus L. encouraged the cultivation of this species for extraction of the product as raw material for the cosmetic industry in the North region of the country (Castellani et al., 2011; Nicoli et al., 2014).
The cultivation of native medicinal plants requires the domestication of the species of interest, which implies technological mastery of all stages of their development, that is, knowledge about the propagation modes, adaptation to the cultivation environment, way of growth, senescence, optimal biomass productivity, and active principles of interest (Reis et al., 2007). Thus, aiming to expand the bio-ecological knowledge of weeds and aromatic species in Cyperaceae, the present work had the objective to characterize the growth of C. sesquiflorus.
MATERIALS AND METHODS
Clumps of C. sesquiflorus were collected from a population in the lawn of the Engineering and Agricultural Sciences Campus of the Federal University of Alagoas, Rio Largo, AL, Brazil (09°28’ S, 35°49’ W and 127 m of altitude). The exsiccates of the botanical material analyzed were identified by a botanist, expert in the family Cyperaceae, and deposited in the Herbarium of the Federal University of Sergipe (ASE), with voucher number ASE 42,592.
After collection, the clumps were washed and divided for propagation into formation of groups with 10 tillers. In each group, 3 tillers were randomly marked with ribbons to evaluate variables along the experiment, especially the tillering behavior. Subsequently, these groups were transplanted into polyethylene bags with a capacity of 0.5 L, filled with sieved soil from areas of natural occurrence of the species. The chemical analysis of the soil was carried out at the Soil Fertility and Plant Nutrition Laboratory (LFSNP/CECA/UFAL). The chemical characteristics of the soil were pH = 5.9; P = 55 mg.dm-3; K = 93 mg.dm-3; Ca+Mg = 6.44 cmolc.dm-3 and V% = 68. The bags were identified with numbered tags and placed in plastic trays for capillary irrigation with water replenishment every three days. The irrigation methodology was adapted from Bryson and Carter (2010).
The experiment was carried out on a bench in a controlled environment greenhouse in a completely randomized design with 4 replications (bags with 10 tillers) and 15 treatments (0, 6, 17, 22, 28, 34, 40, 45, 49, 55, 61, 69, 76, 83, and 90 Days After Transplantation - DAT). Although using time patterns as a treatment or time intervals as a response variable is not common in studies characterizing growth, we adopted the methodology of Carvalho et al. (2005) because in this work the authors analyzed Rottboelia exaltata L.f., a species of the family Poaceae, which is morphologically and phylogenetically close to Cyperaceae. Sixty bags with 10 tillers were used throughout the experiment; the initial day (0) corresponded to December 13, 2019, and the final day (90) corresponded to March 12, 2020. Therefore, the experiment was conducted during three months of the dry period.
The length (cm) of the longest leaf and of the root system was measured with a ruler, while the diameter (mm) of the culm was measured with a digital caliper 0 -150 mm Starrett tools, EC799A-6/150 Ip54, at 0.5 cm above the roots. The number of alive leaves, inflorescences, alive tillers (> 1 cm in length), and dead tillers was counted. All variables were obtained by monitoring the tillers identified in each bag. In these marked tillers (alive or dead), tillering of new individuals > 1 cm was measured.
At the end of the evaluations, the total fresh matter of the aerial and underground parts was obtained in an analytical balance Marte AUW220D, with the exception of day 0. The sampled material was then submitted to drying in a forced air circulation oven until reaching constant weight. After dehydration, the other variables were measured: total, aerial, and underground dry matter (g). For this process, in each treatment (for example 6 DAT), all tillers in the plot were used, that is, the 3 tillers previously marked on day 0 (which had the variables previously measured), the seven other tillers that were also transplanted on day 0, and the new tillers that grew on the clumps during the experiment. This procedure was repeated in the other treatments: 17, 22, 28, 34, 40, 45, 49, 55, 61, 69, 76, 83, and 90 DAT. We highlight that there was no evaluation of regrowth, since the clumps were separated into shoot and root system and the statistical analysis was performed with the mean values obtained from the tillers of each clump. Data were subjected to analysis of variance and means were compared by the Tukey test at 5% probability using the Sisvar software, version 5.6, build 86 – DES/UFLA. Data of the discrete variables were square root (x + 0.5) transformed for homogeneity of variance (ZAR, 1999).
RESULTS AND DISCUSSION
The results on length of the longest leaf, culm diameter, number of leaves, length of the root system, number of inflorescences, and number of tillers of C. sesquiflorus growing in a greenhouse on different DAT are presented in Figure 1a to f. The treatment that resulted in the greatest height (30 cm) of the aerial part (Figure 1a) was 28 DAT, without significant difference from 17, 22, 40 and 61 DAT. Treatments 49, 55, 69 and 90 DAT did not differ statistically from each other and resulted in plants of shorter height (15 cm). From 0 to 28 days of cultivation, the height doubled, going from 15 cm to 30 cm. After an interval of 21 days, recorded plant length values were 15 cm again (49 DAT), starting a new leaf growth cycle. However, in this second cycle, the only treatment in which the height did not differ from 30 cm until the end of the experiment was 61 DAT. In the study by Bryson and Carter (2010), the species C. fuscus sown in a greenhouse reached 15 cm in 6 weeks (42 days) after seedling emergence and doubled in size, reaching a maximum growth of 30 cm, in 9 weeks, that is, 21 days later.
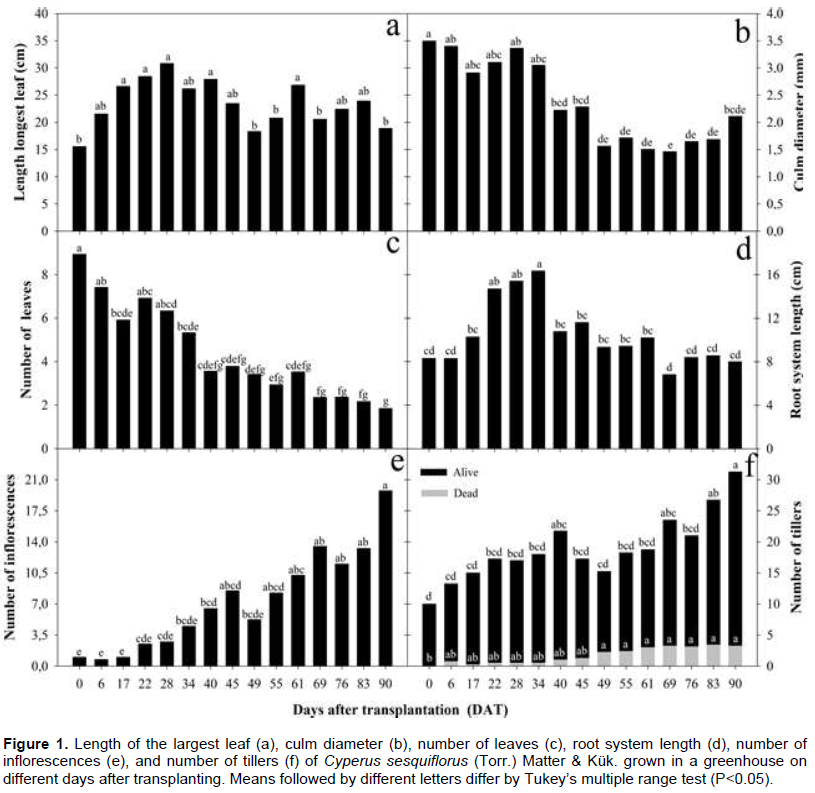
Regarding the diameter of the culm (Figure 1b), it was observed that treatments ranging from 0 to 34 DAT provided maximum diameter values. After this period, the values decreased. As for the number of alive leaves (Figure 1c), there was a marked decrease over the treatments. The best treatment was day 0 (plants with up to 9 leaves) and plants with more than 5 leaves (between 6 and 7.5) were observed until 34 DAT. Leaf senescence may occur as a function of the arrangement of the leaves, since the older ones, located lower in the plant, are shaded by the younger ones, which become a source of photosynthetic assimilates (Moraes and Palhano, 2002). Senescence of leaves in C. rotundus was studied by Brighenti et al. (1997), who also observed that the species had spurts of growth interspersed with rest periods.
Regarding the root system (Figure 1d), the best treatment was 34 DAT, with almost double the value on day 0, followed by days 28 and 22, with a mean of around 15 cm of root. There was a large drop in the growth rate after the peak reached at 34 days, returning close to the length verified on day 0, around 10 cm. In this case, the shortening of the root system may be due to the cultivation container, which, despite facilitating the taking of measures, as mentioned by Benincasa (1988), limits the expansion of the roots as they fold and, consequently, break in the moment of separation for the analysis of this variable. Previous studies indicate the presence of essential oil in the leaves, shoots and underground part of C. sesquiflorus, where there are secretory cells and abundant oil droplets in the parenchyma of the rhizomes (Gonzáles et al., 2009; Novoa et al., 2012; Peckolt and Peckolt, 2016). As whole plants are used to obtain the essential oil from C. sesquiflorus, root length data suggest that cultivation for up to 34 days is the most efficient, as greater root production occurs within this time. Regarding the number of inflorescences and tillers, there was an increase in the production of these organs throughout the cycle (Figure 1e and f), contrary to the other variables that decreased over the days. The treatment with the highest flowering was 90 DAT and the treatments with the lower numbers were 0, 6 and 17 DAT. The peak of flower production suggests that in cultivation to extract essential oil, keeping C. sesquiflorus up to 17 DAT is ideal, since Peckolt and Peckolt (2016) recommend the extraction of oil from this species before flowering.
The best treatment to stimulate vegetative reproduction was also 90 DAT, with the mean density exceeding 30 tillers per clump, that is, it took three months for the number of tillers to triple. Six DAT, the number of tillers increased to five. On the other hand, doubling in tillering occurred initially at 22 DAT. In this study, mortality remained stable from 45 DAT onwards, with about five dead tillers by clump.
In a study on the increase in tillers of C. brevifolius in a temperate region, Rodiyati and Nakagoshi (2005) found a mean number of 9.22 tillers within six weeks, influenced by heat. Nicoli et al. (2014) reported a mean production of 35 rhizomes per plant in C. articulatus growing in a fertilized garden for nine months produced, and a maximum of 62 rhizomes when planted at larger spacing (0.5 m x 0.5 m) and harvested over 12 months later, with surface cleaning and fertilization after 6 months.
According to Hartmann et al. (2011), rhizomes grow by elongation of the growth points produced at the end of terminal and lateral branches, and their length also increases as a function of the growth of the intercalary meristems in the lower part of the internodes. According to Brighenti et al. (1997), the appearance of underground structures competes for the assimilates produced with the vegetative structures of the aerial part, also inducing leaf senescence together with shading, as mentioned by Moraes and Palhano (2002).
The greater tiller emission after three months of cultivation suggests that this is the most viable time for dividing clumps, if the goal is to propagate the species. It was noted that sexual and asexual reproduction happened simultaneously, but further studies are needed to analyze the efficiency of sexual reproduction in terms of seed production and germination. Similar results were found by Carvalho et al. (2005) with the weed R. exaltata (Poaceae). Floral racemes and vegetative branches were produced simultaneously. The decrease in the variables height of the aerial part, number of leaves, diameter and length of roots at 90 days can be explained by the greater emission of tillers, since the tillering phase can result in limited conditions for the growth of other organs. The species C. sesquiflorus exerts great effort in vegetative reproduction, which is typical of perennial plants such as C. brevifolius (Rodiyati and Nakagoshi, 2003).
The data found are in line with the description of Trevisan et al. (2007) about the caespitose growth and the presence of persistent old leaves adhered to the base of mature C. sesquiflorus plants. It was also observed that the clumps presented a perennial habit, and individual tillers have an annual habit, contrasting with the description of annual or perennial habit of short duration in hot climates by Carter (2005) and Bryson and Carter (2008).
The aerial, underground and total dry matter of C. sesquiflorus is shown in Figure 2a to d. A gradual increase in aerial part dry matter was observed and the treatment 90 DAT resulted in a greater weight of approximately 5.0 g (Figure 2a). The dry matter of the underground part was higher in the treatments 76, 83 and 90 DAT, reaching 7.0 g. This is possibly due to the density of new tillers in the clumps (Figure 2b). As for the total dry matter, the treatment with the highest accumulation was 90 DAT, with a value around 10 g (Figure 2c). It was observed that the treatments 6, 76, and 90 DAT resulted in greater biomass in the underground than the aerial parts (Figure 2d). Treatments 17, 22, 28, 34, 40, 45, 49, 55, 69, and 83 DAT caused the partitioning of biomass to be equally distributed between the two parts. The only treatment in which there was greater partitioning of biomass to the aerial part was 61 DAT. According to Peixoto et al. (2020), when plants reach their definitive size, they enter the senescence phase, the leaf area index decreases and less luminous energy is intercepted, resulting in a decrease in dry matter accumulation along with translocation of this matter to reserve organs, and consequently degeneration of the photosynthetic system.
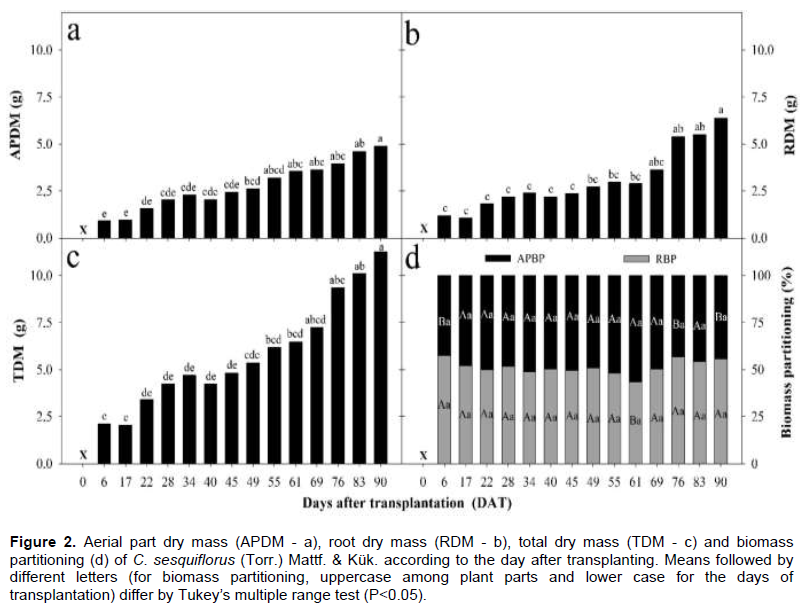
Finally, plant growth is influenced by factors such as age and environmental conditions in which plants live (Reis, 1978). The results obtained here represent the behavior of C. sesquiflorus individuals transplanted as a function of the applied methodology.
CONCLUSIONS
The maximum values of length of the longest leaf and of the root system, diameter of the culm, and number of leaves in cultivated C. sesquiflorus were found up to 34 DAT. The maintenance of the species during the three months of cultivation caused reductions in the values ??of these variables, except for the number of inflorescences and tillers, which increased throughout the experiment, demonstrating that sexual and asexual reproduction occur simultaneously in C. sesquiflorus. The senescence observed in the species occurred due to the shading resulting from the appearance of new leaves. The growth of the underground part was limited by the volume of the container. Thus, our study provides new information on the behavior of C. sesquiflorus that will contribute to upcoming research studies on the growth, control, and essential oil production of this species. It also indicates the period with the lowest number of inflorescences (17 DAT) and highest number of tillers, with confirmation of clump propagation at 90 DAT. However, further investigation under field conditions is necessary.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are thankful to the Coordination for the Improvement of Higher Education Personnel for the financial support provided to the research that originated this article.
REFERENCES
|
Almeida UO, Andrade Neto RC, Lunz AMP, Tavella, LB, Marinho JTS, Nogueira SR (2018). Ocorrência de plantas daninhas em cultivo de bananeira comprida em diferentes espaçamentos no estado do Acre. Journal os Basic Education, Technical e Technological 5(1):188-203. |
|
|
Alves M, Hefler SM,Trevisan R, Silva Filho PJS, Ribeiro ARO (2015). Cyperaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. |
|
|
Benincasa MMP (1998). Análise de crescimento de plantas (Noções básicas). Jaboticabal, FUNEP 42 p. |
|
|
Brighenti AM, Silva JF, Sediyama T, Silveira JSM, Sediyama CS (1997). Análise de crescimento da tiririca (Cyperus rotundus L.), Revista Ceres 44(251):94-110. |
|
|
Bryson CT, Carter R (2008). The Significance of Cyperaceae as Weeds. In. NACSI RFC, FORD BA (Orgs.), Sedges: uses, diversity, and systematics of the Cyperaceae. St. Louis: Missouri Botanical Garden pp. 15-100. |
|
|
Bryson CT, Carter R (2010). Spread, Growth Parameters, and Reproductive Potential for Brown Flatsedge (Cyperus fuscus). Invasive Plant Science and Management 3(3):240-245. |
|
|
Carvalho SJP, Moreira MS, Nicolai M, Vejero RFL, Christoffoleti PJ, Medeiros D (2005). Crescimento e Desenvolvimento da Planta Daninha Capim-Camalote. Bragantia 64(4):591-600. |
|
|
Carter R (2005). An introduction to the sedges of Georgia. Tipularia 20:15-44. |
|
|
Castellani DC, Domenico CI, Roncoletta LMA, Silva AC, Tozaki RM, Oliveira DH (2001). Coeficientes técnicos de produção da priprioca (Cyperus articulatus L.) em sistema orgânico, na região de Belém (PA). Revista Brasileira de Plantas Medicinais 13:606-611. |
|
|
GBIF (2019). Cyperus sesquiflorus (Torr.) Mattf. & Kük. no Secretariado do GBIF: Taxonomia de backbone do GBIF. Conjunto de dados da lista de verificação. |
|
|
Gonzáles Y, Mercado MI, Degen R, Ponessa GI (2009). Morfoanatomía y etnobotánica de rizoma, tallo y escapo de "kapi'i kati", Kyllinga odorata (Cyperaceae) y SUS sustituyentes de Asunción del Paraguay y lrededores. Lilloa 46(1-2):58-67. |
|
|
Hartmann HT, Kester DE, Davies JRFT, Geneve RL (2011). Plant propagation: principles and practices 8th ed. New Jersey: PrenticeHall P 915. |
|
|
Machado Filho HO, Couto EA, Bezerra CP, Mello JIM (2015). Composição e similaridade da flora associada a sítios antropizados do município de João Pessoa - Paraíba. Planta Daninha 33(1):57-66. |
|
|
Matos FJA, Cavalcanti FS, Parente JP (2008). Estudo agronômico qualitativo e quantitativo de Cyperus esculentus L. (junça) - Uma fonte inexplorada de alimento energético. Revista Ciências Agronômicas 39(01):124-129. |
|
|
Moraes A, Palhano AL (2002). Fisiologia da produção de plantas forrageiras. In. Wachowicz CM, Carvalho RIN (Orgs.), Fisiologia Vegetal - Produção e Pós-Colheita. Curitiba: Champagnat pp. 249-272. |
|
|
Nicoli CML, Homma AKO, Menezes AJEA (2014). Aproveitamento da biodiversidade amazônica: o caso da priprioca. In: Homma AKO, Extrativismo vegetal na Amazônia, história ecologia, economia e domesticação. Brasília: EMBRAPA pp. 99-105. |
|
|
Novoa M, Colares M, Arambarri A (2012). Anatomy of monocotyledons: stems and rhizomes of land herbs used as medicinal in the Río de La Plata area (Argentina). Bonplandia 21(2):149-157. |
|
|
Nunes FS, Schaedler CE, Chiapinotto DM (2018). Weed Phytosociological Survey in Irrigated Rice. Planta daninha, 36. |
|
|
Peckolt T, Peckolt G (2016). História das plantas úteis e medicinais do Brasil. Fino Traço 904 p. |
|
|
Peixoto CP, Almeida AT, Santos, JMS, Poelking VGC, Oliveira ER (2020). Análise Quantitativa do Crescimento de Plantas. In. Peixoto CP, AlmeidaAT, Santos JMS, Poelking VGC, Oliveira ER. Curso de Fisiologia vegetal. Cruz das Almas, BA: UFRB pp. 153-168. |
|
|
Reis GG (1978). Análise de crescimento de plantas - mensuração do crescimento. Belém: CPATU/EMBRAPA P 23. |
|
|
Reis MS, Mariot A (2007). Diversidade e domesticação de plantas medicinais. In. Simões CMO, Schenkel EP, Gosmann G, Mello JCP, Mentz LA, Petrovick PR. Farmacognosia: da planta ao medicamento. Porto Alegre: Editora da UFRGS pp. 45-74. |
|
|
Rodiyati A, Nakagoshi N (2003). Annual Growth and Phenology of Kyllinga brevifolia (Rottb.) Hassk. In Temperate and Tropical Regions. Journal of International Development and Cooperation 10(1):1-10. |
|
|
Simpson DA, Inglis CA (2001). Cyperaceae of economic, ethnobotanical, and horticultural importance: A checklist. Kew Bulletin 56:257-360. |
|
|
Soares DJ, Oliveira WS, Uzuele EL, Carvalho SJP, Ovejero RFL, Christoffoleti PJ (2017). Growth and development of Conyza bonariensis based on days or thermal units. Pesquisa agropecuária brasileira 52(1). |
|
|
Trevisan R, Lüdtke R, Boldrini II (2007). O gênero Kyllinga Rottb. (Cyperaceae) no Rio Grande do Sul, Brasil. Revista Brasileira de Biociências pp. 27-36. |
|
|
Zar JH (1999). Biostatiscal Analyses. New Jersey: Prentice Hall P 663. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


