Review

ABSTRACT
Weeds cause great problems to humankind by interfering in food production, health, economic stability, and welfare. The overuse of synthetic herbicides for weed control eventually leads to the evolution of herbicide resistant weeds, which also resulted in growing public concern over their impacts upon human health. The intensification of arable farming has also had a simultaneous impact on the environment. This intensification of the landscape may disrupt natural processes, such as resistance to plant invasion. Therefore, the aim of this review is to address these production constraints by suggesting applications of allelopathy via smother cropping, because this may ease the incidence of herbicide resistance and in the process promote cultivated plant diversity and thereby maintaining healthy agroecosystems. Weed resistance to herbicides presents one of the greatest current economic challenges to agriculture. System-oriented approaches to weed management that make better use of alternative weed management tactics need to be developed. Plant roots exude a wide variety of metabolites, some of which may act as allelochemicals and mediate interactions between plants and other organisms. These metabolites are in essence chemicals from nature which may be exploited for weed management as an alternative weed control option. Smother crops, as well as its mulches, have been shown to release allelochemicals, which were inhibitory to weeds. The principal goal of smother crops is to control weeds by replacing an unmanageable weed population with a manageable smother crop. More data from the grey area where agriculture and ecology overlap will enable the greater use of ecosystem services for crop protection in agricultural production and consequently reduce the incidence of resistance to agricultural chemicals.
Key words: Agroecosystems, allelopathy, cultivated ecosystems, herbicide resistance, smother cropping, mulching.
INTRODUCTION
ALLELOPATHY THEORY
HERBICIDE RESISTANCE
AGROECOSYSTEMS
PRACTICAL APPLICATIONS
DISCUSSION
FUTURE RESEARCH NEEDS
CONFLICT OF INTERESTS
REFERENCES
|
Alsaadawi IS, Zwain KHY, Shahata HA (1998). Allelopathic inhibition of growth of rice by wheat residues. Allelopathy J. 5:163-169. |
|
|
An M, Pratley JJE, Haig TT (2001). Phytotoxicity of Vulpia residues: IV. Dynamics of allelochemicals during decomposition of Vulpia residues and their corresponding phytotoxicity. J. Chem. Ecol. 27:395-409. |
|
|
Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM (2003). Allelopathy and exotic plant invasion: from molecules and genes to species interactions. Science 301:1377-1380. |
|
|
Barberi P (2002). Weed management in organic agriculture: are we addressing the right issues? Weed Res. 42:177-193. |
|
|
Batish DR, Singh HP, Kohli RK, Kaur S (2001). Crop allelopathy and its role in ecological agriculture. J. Crop. Prod. 4:121-161. |
|
|
Batish DR, Tung P, Singh HP, Kohli RK (2002). Phytotoxicity of sunflower residues against some summer season crops. J. Agron. Crop. Sci. 188:19-24. |
|
|
Baucom RS (2009). A herbicide defense trait that is distinct from resistance: The evolutionary ecology and genomics of herbicide tolerance. IN: Stewart CN Jr. (ed.), Weedy and Invasive Plant Genomics. Wiley-Blackwell, Iowa, USA. |
|
|
Belz RG (2004). Evaluation of allelopathic traits in Triticum L. spp. and Secale cereale L. PhD Thesis, University of Hohenheim, Stuttgart, Germany. |
|
|
Belz RG, Duke SO, Hurle K (2005). Dose-response – a challenge for allelopathy. Nonlinearity Biol. Toxicol. Med. 3:173-211. |
|
|
Belz RG, Reinhardt CF, Foxcroft LC, Hurle K (2007). Residue allelopathy in Parthenium hysterophorus L. does parthenin play a leading role? Crop Prot. 26:237-245. |
|
|
Bertin C, Yang X, Weston LA (2003). The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256:67-83. |
|
|
Blum U (1996). Allelopathic interactions involving phenolic acids J. Nematol. 28:259-267. |
|
|
Brecke BJ, Shilling DG (1996). Effect of crop species, tillage, and rye (Secale cereale) mulch on sicklepod (Senna obtusifolia). Weed Sci. 44:133-136. |
|
|
Bullock JM, Pywell RF, Walker KJ (2007). Long term enhancement of agricultural production by restoration of biodiversity. J. Appl. Ecol. 44:6-12. |
|
|
Chalker-Scott L (2007). Impact of mulches on landscape plants and the environment-a review. J. Environ. Hortic. 25:239-249. |
|
|
Chao WS, Horvath DP, Anderson JV, Foley M (2005). Potential model weeds to study genomics, ecology, and physiology in the 21st century. Weed Sci. 53:927-937. |
|
|
Chou CH (1999). Roles of allelopathy in plant biodiversity and sustainable agriculture. Crit. Rev. Plant Sci. 18:609-636. |
|
|
Clergue B, Amiaud B, Pervanchon F, Lasserre-Joulin F, Plantureux S (2005). Biodiversity: function and assessment in agricultural areas. A review. Agron. Sust. Dev. 25:1-15. |
|
|
Daldon BR, Blum U, Weed SB (1983). Allelopathic substances in ecosystems: Effectiveness of sterile soil components in altering recovery of ferulic acid. J. Chem. Ecol. 9:1185-1201. |
|
|
Dorn B, Jossi W, van der Hejiden MGA (2015). Weed suppression by cover crops: comparative on-farm experiments under integrated and organic conservation tillage. Weed Res. 55(6):586-597. |
|
|
Duke SO, Cedergreen N, Velini ED, Belz RG (2006). Hormesis: is it an important factor in herbicide use and allelopathy? Outl. Pest. Manage. 17:29-33. |
|
|
Duke SO, Dayan FE, Romagni JG, Rimando AM (2000). Natural products as sources of herbicides: current status and future trends. Weed Res. 40:99-111. |
|
|
Duke SO, Rimando AM, Bearson SR, Scheffler BE, Ota E, Belz RG (2002). Strategies for the use of natural products for weed management. J. Pest. Sci. 27:298-306. |
|
|
Dyck E, Liebman M (1994). Soil fertility management as a factor in weed control: the effect of crimson clover residue, synthetic nitrogen fertilizer, and their interaction on emergence and early growth of lambsquarters and sweet corn. Plant Soil 167:227-237. |
|
|
Efthimiadou AP, Karkanis AC, Bilalis DJ, Efthimiadis P (2009). Review: the phenomenon of crop-weed competition; a problem or a key for sustainable weed management? J. Food Agric. Environ. 7:861-868. |
|
|
Einhellig FA (1996). Interactions involving allelopathy in cropping systems. Agron. J. 88:886-893. |
|
|
FAO (Food and Agriculture Organization of the United Nations) (2008). Investing in sustainable agricultural intensification: the role of conservation agriculture. Rome: FAO. |
|
|
Fay PK, Duke WB (1977). An assessment of allelopathic potential in Avena germplasm. Weed Sci. 25:224-228. |
|
|
Ferreira MI, Reinhardt CF (2010). Field assessment of crop residues for allelopathic effects on both crops and weeds. Agron. J. 102:1593-1600. |
|
|
Gaba S, Bretagnolle F, Rigaud T, Philippot L (2014). Managing biotic interactions for ecological intensification of agroecosystems. Front. Ecol. Evol. 2:1-9. |
|
|
Gaba S, Fried G, Kazakou E, Chauvel B, Navas ML (2014). Agroecological weed control using a functional appraoch: a review of cropping system diversity. Agron. Sustain. Dev. 34:103-119. |
|
|
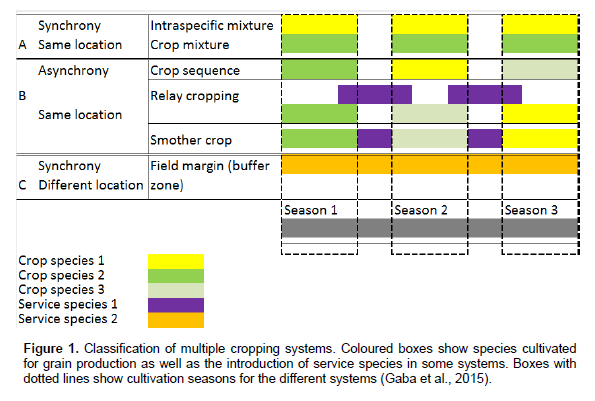
Gaba S, Lescourret F, Boudsocq S, Enjalbert J, Hinsinger P, Journet EP, Navas ML, Wery J, Louarn G, Malézieux E, Pelzer E, Prudent M, Ozier-Lafontaine H (2015). Multiple cropping system as drivers for providing multiple ecosystem services: from concepts to design. Agron. Sustain. Dev. 35:607-623. |
|
|
Gallandt ER, Molloy T, Lynch RP, Drummond FA (2005). Effect of cover-cropping systems on invertebrate seed predation. Weed Sci. 53:69-76. |
|
|
Gealy DR, Wailes EJ, Esterninos LE, Chaves RSC (2003). Rice cultivar differences in suppression of barnyardgrass (Echinoloa crus-galli) and economics of reduced propanil rates. Weed Sci. 51:601-609. |
|
|
Gressel J (1991). Why get resistance if it can be prevented or delayed. In: Caselay JC, Cussans GW, Atkin RK (eds.). Herbicide resistance |
|
|
in weeds and crops. Oxford: Butterworth-Heinemann. pp. 1-26. |
|
|
Harker KN (2013). Slowing weed evolution with integrated weed management. Can. J. Plant Sci. 93:759-764. |
|
|
Hartwig NL, Ammon HU (2002). 50th Anniversary-Invited article: Cover crops and living mulches. Weed Sci. 50:688-699. |
|
|
Heap I (2015). International survey of herbicide resistant weeds. |
|
|
Hoffman ML, Weston LA, Snyder JC, Regnier EE (1996). Allelopathic influence of germinating seeds and seedlings of cover crops on weed species. Weed Sci. 44: 579-584. |
|
|
Hoult AHC, Lovett JV (1993). Biologically active secondary metabolites of barley. III. A method for identification and quantification of hordenine and gramine in barley by high-performance liquid chromatography. J. Chem. Ecol. 19:2245-2254. |
|
|
Jones E, Jessop RS, Sindel, BM, Hoult A (1999). Utilising crop residues to control weeds. p. 373-376. IN: Bishop, A. Boersma, M. and Barnes, C.D. (ed.), Proceedings of the 12th Australian Weeds Conference. Tasmanian Weeds Society, Devonport, Australia. |
|
|
Kazinczi G, Horváth J, Takács AP (2005). Plant-plant and plant-virus interactions. Lectures and papers presented at the 7th Slovenian Conference on Plant Protection, ZreÄe, Slovenia, 8-10 March. |
|
|
Khanh TD, Chung MI, Xuan TD, Tawata S (2005). The exploitation of crop allelopathy in sustainable agricultural production. J. Agron. Crop Sci. 191:172-184. |
|
|
Kohli RK, Batish DR, Singh HP (2006). Allelopathic interactions in agroecosystems. IN: Reigosa MJ, Pedrol N, González L (Eds.). Allelopathy: a physiological process with ecological implications. Dordrecht: Springer. pp. 465-493. |
|
|
Kong CH, Wang P, Zhao H, Xu XH, Zhu YD (2008). Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol. Biochem. 40:1862-1869. |
|
|
Korres NE, Norsworthy JK (2015). Influence of a rye cover crop on the critical period for weed control in cotton. Weed Sci. 63:346-352. |
|
|
Kruidhof HM (2008). Cover crop-based ecological weed management: exploration and optimization. PhD Thesis, Wageningen University, Wageningen, The Netherlands. 156p. |
|
|
Kumar V, Brainard DC, Bellinder RR (2008). Suppression of Powell amaranth (Amaranthus powellii), shepherd's-purse (Capsella bursa-pastoris), and corn chamomile (Anthemis arvensis) by buckwheat residues: role of nitrogen and fungal pathogens. Weed Sci. 56:271-280. |
|
|
Lichtfouse E, Navarrete M, Debaeke P, Souchère V, Alberola C, Ménassieu J (2009). Agronomy for sustainable agriculture: A review. Agron. Sustain. Dev. 29:1-6. |
|
|
Liebman M, Davis A (2000). Integration of soil, crop and weed management in low-external-input farming systems. Weed Res. 40:27-48. |
|
|
Liebman M, Mohler CL (2001). Weeds and the soil environment. In: Ecological Management of Agricultural Weeds, Liebman M, Mohler CL, Staver CP (Eds.), Cambridge University Press, Cambridge. pp. 210-268. |
|
|
Llewellyn RS, Lindner RK, Pannell DJ, Powles SB (2007). Herbicide resistance and the adoption of integrated weed management by Western Australian grain growers. Agric. Econ. 36:123-130. |
|
|
Mamolos AP, Kalburtji KL (2001). Significance of allelopathy in crop rotation. J. Crop Prod. 4:197-218. |
|
|
Marble SC (2015). Herbicide and mulch interactions: a review of the literature and the implications for the landscape maintenance industry. Weed Technol. 29:341-349. |
|
|
McCalla TM, Norstadt FA (1974). Toxicity problems in mulch tillage. p. 29. IN: Physiology of stressed crops. Volume III. The Stress of Allelochemicals, 2005. U.S. Gupta (ed.), University of Georgia, Science Publishers, Inc., Enfield (NH), USA. |
|
|
MédiÑne S, Valantin-Morison M, Sarthou JP, de Tourdonnet S, Gosme M, Bertrand M, Roger-Estrade J, Aubertot JN, Rusch A, Motisi N, Pelosi C, Doré T (2011). Agroecosystem management and biotic interactions: A review. Agron. Sustain. Dev. J. 31:491-514. |
|
|
Melander B, Munier-Jolain N, Charles R (2013). European perspectives on the adoption of non-chemical weed management in reduced-tillage systems for arable crops. Weed Technol. 27:231-240. |
|
|
Mohler CL, Teasdale JR (1993). Response of weed emergence to rate of Vicia villosa Roth and Secale cereale L. residue. Weed Res. 33:487-499. |
|
|
Nichols V, Verhulst N, Cox R, Govaerts B (2015). Weed dynamics and conservation agriculture principles: A Review: Field Crops Res. 183:56-68. |
|
|
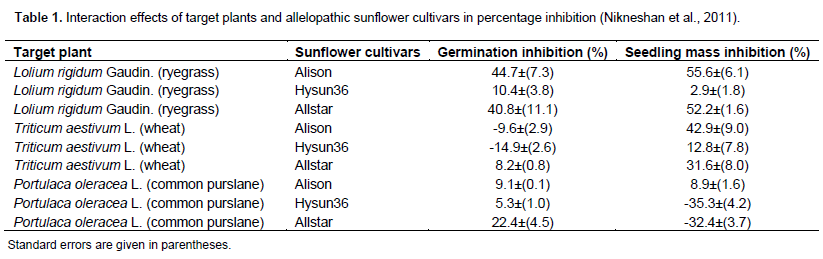
Nikneshan P, Karimmojeni H, Moghanibashi M, Hosseinei N (2011). Allelopathic potential of sunflower on weed management in safflower and wheat. Aust. J. Crop Sci. 11:1434-1440. |
|
|
Olofsdotter M (2001). Getting closer to breeding for competitive ability and the role of allelopathy – an example from rice (Oryza sativa). Weed Technol. 15:798-806. |
|
|
Olofsdotter M, Navarez D, Rebulanan M, Streibig JC (1999). Weed-suppressing rice cultivars – does allelopathy play a role? Weed Res. 39:441-454. |
|
|
Olson DM, Wäckers FL (2007). Management of field margins to maximize multiple ecological services. J. Appl. Ecol. 44:13-21. |
|
|
Paine LJ, Harrison H (1993). The historical roots of living mulch and related practices. HortTechnology 3:137-142. |
|
|
Peachey RE, Williams RD, Mallory-Smith C (2004). Effect of no-till or conventional planting and cover crops residues on weed emergence in vegetable row crop. Weed Technol. 18:1023-1030. |
|
|
Phatak SC (1992). An integrated sustainable vegetable production system. HortTechnology 27:738-741. |
|
|
Pieterse PJ (2010). Herbicide resistance in weeds – a threat to effective chemical weed control in South Africa. S. Afr. J. Plant Soil. 27:66-73. |
|
|
Prati D, Bossdorf O (2004). Allelopathic inhibition of germination by Alliaria petiolate (Brassicaceae). Am. J. Bot. 91:285-288. |
|
|
Price AJ, Norsworthy JK (2013). Cover crops for weed management in southern reduced tillage vegetable cropping systems. Weed Technol. 27:212-217. |
|
|
Purvis CE (1990). Differential Response of Wheat to Retained Crop Stubbles. I. Effect of Stubble Type and Degree of Decomposition. Aust. J. Agric. Res. 41:225-42. |
|
|
Putnam AR, DeFrank J (1983). Use of phyto toxic plant residues for selective weed control. Crop Prot. 2:173-181. |
|
|
Qasem JR, Foy CL (2001). Weed allelopathy, its ecological impacts and future prospect: a review. J. Crop Prod. 4:43-120. |
|
|
Qasem JR, Hill TA (1989). Possible role of allelopathy in the competition between tomato, Senecio vulgaris L. and Chenopodium album L. Weed Res. 29:349-356. |
|
|
Reinhardt CF, Meissner R, Labuschagne N (1994). Allelopathic interaction of Chenopodium album L. and certain crop species. S. Afr. J. Plant Soil 11:45-49. |
|
|
Singh HP, Batish DR, Kohli RK (2001). Allelopathy in agroecosystems: an overview. J. Crop Prod. 4:1-41. |
|
|
Singh HP, Batish DR, Kohlia RK (2003). Allelopathic interactions and allelochemicals: new possibilities for sustainable weed management. Crit. Rev. Plant Sci. 22:239-307. |
|
|
Sozeri S, Ayhan A (1998). Effect of stubble on wheat seed germination and seedling growth. p. 29. In: Physiology of stressed crops. Volume III. The Stress of Allelochemicals 2005. Gupta US (Ed.), University of Georgia, Science Publishers, Inc., Enfield (NH),USA. |
|
|
Storkey F, Lutman P (2008). How can weed management support the development of a more multifunctional agriculture? In: Proceedings of the Fifth International Weed Science Congress, 22-27 June 2008. Vancouver. Canada [abstract only]. pp. 215-216. |
|
|
Teasdale JR, Abdul-Baki AA, Bong Park Y (2008). Sweet corn production and efficiency of nitrogen use in high cover crop residue. Agron. Sustain. Dev. 28:559-565. |
|
|
Teasdale JR, Mohler CL (2000). The quantitative relationship between weed emergence and the physical properties of mulches. Weed Sci. 48:385-392. |
|
|
Teasdale JR, Pillai P (2005). Contribution of ammonium to stimulation of smooth pigweed (Amaranthus hybridus L.) germination by extracts of hairy vetch (Vicia villosa Roth) residue. Weed Biol. Manage. 5:19-25. |
|
|
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005). Landscape perspectives on agricultural intensification and biodiversity – ecosystem service management. Ecol. Lett. 8:857-874. |
|
|
Vyvyan JR (2002). Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 58:1631-1646. |
|
|
Weston LA (1996). Utilization of allelopathy for weed management in agroeco-systems. Agron. J. 88:860-866. |
|
|
Weston LA, Duke SO (2003). Weed and crop allelopathy. Crit. Rev. Plant Sci. 22:367-389. |
|
|
Wu HW, Pratley J, Haig T (2003). Phytotoxic effects of wheat extracts on a herbicide-resistant biotype of annual ryegrass (Lolium rigidum). J. Agric. Food Chem. 51:4610-4616. |
|
|
Wu HW, Pratley J, Lemerle D, Haig T (1999). Crop cultivars with allelopathic capability. Weed Res. 39:171-180. |
|
|
Xuan TD, Tawata S, Khanh TD, Chung MI (2005). Decomposition of allelopathic plants in soil. J. Agron. Crop Sci. 191:162-171. |
|
|
Young SL (2012). True integrated weed management. Weed Res. 52:107-111. |
|
|
Yuan JS, Tranel PJ, Stewart CN Jr (2007). Non-target-site herbicide resistance: A family business. Trends Plant Sci. 12:6-13. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


