Full Length Research Paper

ABSTRACT
This work aimed at determining the optimum harvest time of African eggplant (Solanum aethiopicum L.) seeds, cv. Oforiwa and cv. Kpando for maximum physiological seed quality under tropical (TCC) and temperate oceanic climatic (TOC) conditions. Fruit morphological metrics such as fruit weight and size were also related to seed physiological quality. Seeds were harvested at different maturation stages from 20 to 82 days after anthesis (DAA). Seed quality was assessed according to moisture content, dry seed weight, seed length, thousand seed weight, percentage germination and emergence. The results indicated that seeds harvested precociously (20 and 34 days after anthesis), did not germinate or recorded very low percentage germination (0-20%). Considering all the fruit morphological traits and seed quality variables measured, the results indicate that maximum seed quality (that is, germination, emergence, seed weight and mean germination time) are obtained from fruits harvested at 76 DAA. The physiological maturity (PM) (maximum seed dry weight) for cv. Oforiwa was attained at 62 DAA while cv. Kpando took 14 days more to reach physiological maturity but coincided with maximum germination at 76 DAA. The results further showed that fruit characteristics such as weight and size are associated with seed physiological quality in African eggplant. It is thus recommended that fruits of African eggplant cv. Oforiwa and cv. Kpando should be harvested at 70 to 76 DAA for maximum seed germination and emergence.
Key words: African eggplant, Solanum aethiopicum, physiological maturity, germination, seed maturation.
INTRODUCTION
The Gilo group of the African eggplant (Solanum aethiopicum L.) is one of the most important vegetable crops cultivated by many resource constrained smallholder farmers in West and Central Africa including Ghana (Weinberger and Msuya, 2004; Osei et al., 2010). The absence of a reliable seed system for this crop has resulted in reliance on informal seed exchange among farmers and fresh fruit traders. Seeds obtained from these informal, farmer-saved or ‘trader-saved’ sources have low germination and field emergence. This phenomenon could be attributed to physiological seed dormancy as observed in Solanum melongena (Yogeesha et al., 2006) and Solanum tovum (Cutti and Kulckzynski, 2016), which are close relatives of S. aethiopicum. It could also be due to harvesting and extracting seeds that are not matured. Although seed physiological quality is genetically determined (Linkies and Leubner-Metzger, 2012; Yan et al., 2014), it is often influenced by factors such as fruit maturity stage at harvest and environment (Vidigal et al., 2011; Bortey and Dzomeku, 2016; Singkaew et al., 2017).
Studies regarding seed maturation contribute significantly in determining the ideal fruit development stage for harvesting to obtain high quality seeds. Particularly, for crops that experience continuous flowering and fruiting such as the African eggplant due to their indeterminate growth habit, fruits of different physiological maturity can be found on the same plant, making it difficult to determine the most ideal fruit harvesting time.
Seed maturation is one of the important components of seed quality and a prerequisite for successful germination and emergence. However, the stage of seed development and maturation at which seeds attain maximum quality and its association with fruit characteristics is subject of controversy. Several studies have demonstrated that there are variations among plant species and growing conditions in occurrence of maximum seed quality during development (Takac et al., 2015; Bortey and Dzomeku, 2016; Ribalta et al., 2017; Tetteh et al., 2021). Additionally, the quality of seed and its association with fruit maturation characteristics have been reported with varied conclusions (Demir and Ellis, 1992; Kortse et al., 2017; Tetteh et al., 2018).
During seed development, the maximum seed germination may coincide with the maximum dry matter accumulation that characterizes physiological maturity or mass maturity (TeKrony and Egli, 1997) as observed for okra (Demir and Ermis, 2005; Bortey and Dzomeku, 2016), selected Solanum aethiopicum species (Tetteh et al., 2021) and Allophylus edulis fruits (Kaiser et al., 2016). Other studies have also reported that maximum seed germination may not coincide with maximum dry matter accumulation as reported for tomato (Borges et al., 2019), pepper (Ruiz and Parera, 2017), safflower (Ramos et al., 2021) and eggplant (Demir et al., 2002). In the latter reports, seed germination decreases after attaining maximum germination (Ellis, 2019).
There is little known regarding the African eggplant (S. aethiopicum L.) in relation to the seed and fruit quality changes that occur during development and maturation and how this influences seed physiological quality. This study was conducted to determine the ideal harvest time for maximum seed germinability and field emergence in two cultivars produced under tropical and temperate oceanic climatic conditions. The association of fruit morphological characteristics, such as fruit weight, diameter and length (size) with physiological quality of African eggplant seeds were also investigated.
MATERIALS AND METHODS
Study location characteristics
The experiments were conducted in two locations: University of Eldoret Agriculture Research Field, Chepkoliel in Uasin Gishu County (N00° 34.468´ E 035° 18.044´) and Mabanga Agriculture Training Centre (ATC) in Bungoma County (Kenya) (N00° 36.222´E034° 37.392´). Chepkoliel (Eldoret) falls under a temperate oceanic climate (TOC), (Cfb) while Mabanga in Bungoma County falls under a tropical climate (TCC) (Am) according to Koppen climate classification (Köppen et al., 2011). Chepkoliel (Eldoret) is a highland plateau with altitude ranging between 1500 and 2100 m above sea level. Annual rainfall 1103 mm and has Oxissols soils. The maximum and minimum temperatures were 24.5 and 11.6°C, respectively during the field experiments. The tropical study area (Bungoma) has abundant and well distributed rainfall with an average annual rainfall of 1600 mm. The soils are classified as ferrasols (Jaetzold et al., 2006). The maximum and minimum temperatures recorded during the field experiments were 36.9°C and minimum of 15.3°C, respectively.
Plant
Seeds of two cultivars of African eggplant (S. aethiopicum L.) cv. Oforiwa (Round-shaped fruit) and cv. Kpando (Elongated to Blocky shaped fruit) were obtained from CSIR-Crops Research Institute, Kumasi, Ghana for this study.
Nursery establishment and transplanting
Seeds were sown in a seed germination tray (54 cm × 28 cm × 4.3 cm) containing a mixture of top soil mixed with cocopeat at a ratio of 1:1 at the nursery. One seed was sown per cell at a depth of approximately 3 mm. Seedlings were transplanted after 6 weeks old (with a minimum of three true leaves) to an open field at a spacing of 0.75 m intra-rows and 0.6 m inter-rows rows. The experiment was laid in a randomized complete block design in a factorial arrangement with two varieties of African eggplant and six harvesting stages in four replications. Treatment or plot size was 6.5 m × 4 m. There were 8 plants per row with 6 rows per plot constituting a total of 48 plants per plot.
Flower tagging and fruit harvesting:
Plants started flowering from 65 to 70 days depending on the cultivar. Flowers were tagged at anthesis and fruits were harvested at two weeks intervals except the last harvest. In all six harvesting, maturity stages were studied: 20, 34, 48, 62, 76 and 82 days after anthesis (DAA). At each harvest stage, 20 fruits were randomly selected from the various maturity stages to determine the fruit morphometric measurements.
Fruit morphological and seed physical data collected
Fruit weight (g)
Fruit weights of 20 fruits were measured using a weighing balance (0.01 g) and the mean weight recorded and expressed in grams (g).
Fruit length (mm)
The fruit length was measured from the shoulder of the fruit to the blossom end, excluding the peduncle using a digital vernier calliper. The mean fruit length was recorded and expressed in millimetres (mm).
Fruit width (mm)
The fruit width was measured at the widest middle part of the fruit using a digital vernier calliper and the mean expressed in millimetres (mm).
Seed number per fruit
Seeds were extracted not later than 24 h after harvest by cutting fruits opened and removing seeds under water. Seeds were thoroughly washed in running tap water, followed by rinsing in deionised water. The number of seeds per fruit was determined by counting seeds extracted from 5 randomly selected fruits at each harvest stage in five replicates and the mean recorded.
Fresh seed weight (g)
To determine the fresh seed weight (g), 100 seeds were drawn from the seed bulk extracted fresh seeds and weight recorded.
Dry seed weight (g) and seed moisture content (%)
The dry seed weight (g) and seed moisture content were determined by drying seeds at a low constant temperature of 105 ± 3°C for 24 h (Brasil, 2009). 100 seeds of four replicates were used to determine the dry seed weight. For seed moisture content, 2 g of two replicates of fresh seeds were dried at the low constant temperature described earlier and calculated (fresh weight basis).
The 1000 seed weight (g) and seed length (mm)
1000 seed weight was determined by counting and weighting eight replicates of 100 seeds after seeds were air-dried for 3 days. The mean weight was expressed in grams. Due to the size of the seed, the seed length (mm) was determined by placing 10 seeds along a measuring rule and the readings were recorded. The seed length per each seed was obtained by dividing the recorded reading by 10.
Seed germination percentage determination
Four replicates of 25 seeds were placed in Petri dishes on two sheets of filter paper (PRAT DUMAS) moistened with 7 ml of distilled water and incubated (Biobase: BJPX-B400II) at 20/30°C with 16/8 h of light and dark period for 14 days (Botey et al., 2021a). Blotter paper was regularly moistened to ensure seeds have adequate moisture to germinate. The seed tests were uniform for all seed lots. Seed germination was observed and counted daily for 14 days. Seeds were considered germinated when there was a minimum of 2 mm protrusion of the radicle.
Seedling emergence test
The emergence test was conducted for 21 days from sowing to final recordings. Four replicates of 30 seeds at each harvesting maturity stage were sown in top soil mixed with cocopeat at a ratio of 1:1 as used during the nursery stage in plastic trays (54 cm × 28 cm × 4.3 cm) under a screen house condition. The maximum and minimum temperatures during the experiment were 34.4 and 12.8°C, respectively. Emerged seedlings were counted daily for 21 days and final percentage emergence was calculated at the end of the test using the formula as follows: Percentage Emergence (%) = total seedlings emerged/total number of seeds sown × 100. A seed was considered emerged when the first two true leaves protruded out at about 5 to 7 mm above the soil.
Determination of seed quantitative traits and correlation dynamics of fruit traits and seed quality
Seed quantitative traits such as mean germination time (MGT), time to 50% germination (T50), germination index (GI) and mean daily germination percent (MGD) were calculated using the Advanced Germination Measurement Tool (Khalid, 2018). The correlation dynamics between fruit morphological traits and these seed quantitative traits were determined using Microsoft Excel.
Statistical design and analysis
The data collected were subjected to analysis of variance at 5% significant level (p ≤ 0.05) using GenStat 14th Edition. In each environment the effect of African eggplant variety/cultivar and harvesting times were tested on the parameter measured in a two-way ANOVA. African eggplant variety/cultivar and harvesting time were fixed factors while block and replicate were random effects. Values expressed in percentages were arcsine transformed. Data were checked for normality of residual distribution and variance homogeneity (Shapiro-Wilk test). Treatment means were compared by Tukey test at p ≤ 0.05.
RESULTS
Seed physical and physiological changes during development of the African eggplant (S. aethiopicum L.)
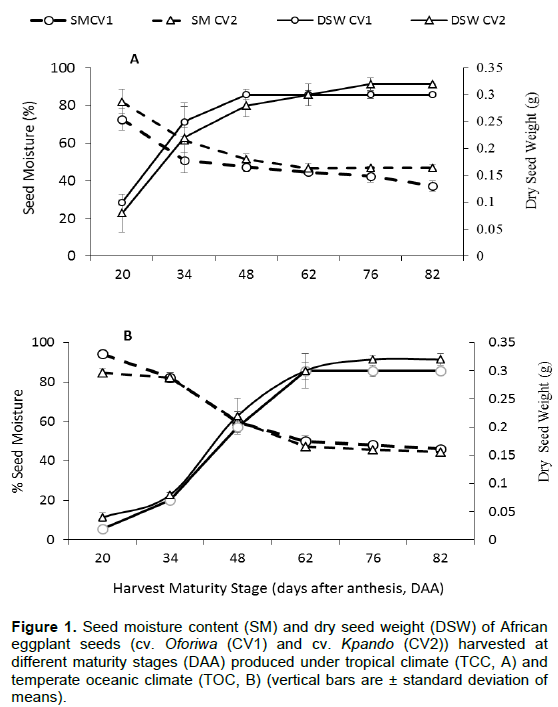
The results showed significant difference for seed moisture content and dry seed weight as a function of maturity (p < 0.001) under both environments (Figure 1). Plants began flowering 65 to 70 days after seedling emergence. Seed moisture content at first harvest (20 days after anthesis) was approximately 73 and 82% for cv. Oforiwa and cv. Kpando, respectively and declined rapidly and linearly during fruit development and maturation. Maximum seed dry weight was observed at 48 DAA in cv. Oforiwa while it took approximately a month later for cv. Kpando (76 DAA) to attain maximum seed dry weight, when seeds were produced under a tropical climate (TCC, A) (Figure 1A). At this point of physiological maturity, the seed moisture contents for cv. Oforiwa and cv. Kpando, were 59 and 46%, respectively. Similarly, seeds produced under temperate oceanic climate (TOC, B) conditions recorded higher seed moisture at early harvest and declined almost linearly as harvest delayed. It was however, observed that physiological maturity (maximum dry seed weight) for cv. Oforiwa was attained at a much earlier stage (62 DAA) while cv. Kpando was at the same maturity stage at 76 DAA (Figure 1B).
Relationship of seed moisture content on percentage emergence as influenced by harvest maturity
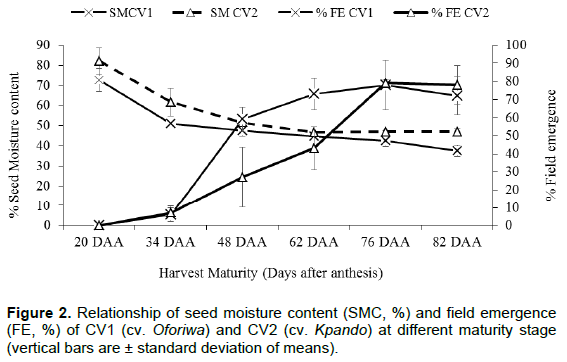
The effect of seed moisture content on seedling emergence during fruit development is as shown in Figure 2. At second harvests (34 DAA), although seeds had acquired ability to germinate (<10%), emergence was very low. As harvest was delayed, emergence percentage increased (Figure 2). Emergence percentage was the highest at 76 DAA for both cv. Oforiwa and Kpando and at 42.5 and 46% seed moisture content and then decreased steadily. It was however, observed that the increase in emergence percentage was significantly (p < 0.01) higher in cv. Oforiwa than in cv. Kpando between the development phase of 34 and 76 DAA.
The effect of harvest maturity on seed germination and field emergence under both tropical and temperate climates
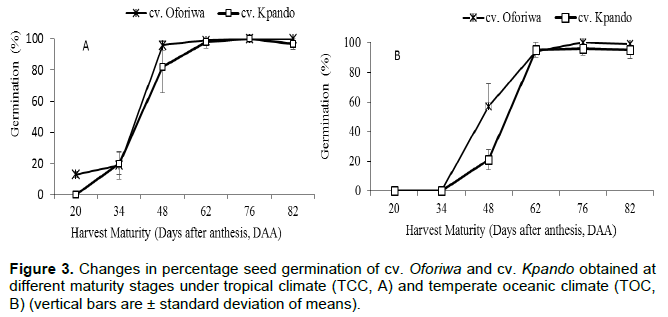
Percentage seed germination and field emergence at different harvest maturity stages is presented for seeds produced under both tropical and temperate oceanic climates (Figure 3A and B), respectively. The effects of time of harvest on increasing germination percentage and seedling emergence were significant (p < 0.01) for both seed production environments. The germination behaviour also differed among the cultivars. In the early stages of development, no germination occurred for seeds produced under a tropical climate (A) for cv. Kpando while <10% germination was recorded for cv. Oforiwa. Germination then increased sharply in fruits harvested from 48 DAA and was maximum (100%) for both cultivars at 76 DAA and maintained or marginally declined but not sharply thereafter (Figure 3A). Fruits harvested at early stages (20 and 34 DAA) from the temperate oceanic climate (B), however failed to germinate for both cultivars until the third stage of harvest (48 DAA). Later harvests recorded a steady increase in percentage germination until it reached a peak of 98% at 76 DAA and slightly declines afterwards (Figure 3B). The maximum percentage germination coincided with the physiological maturity (PM) (maximum dry seed weight) for cv. Kpando at 76 DAA but differed for cv. Oforiwa.
Effect of harvest maturity on percentage field emergence under tropical and temperate climates
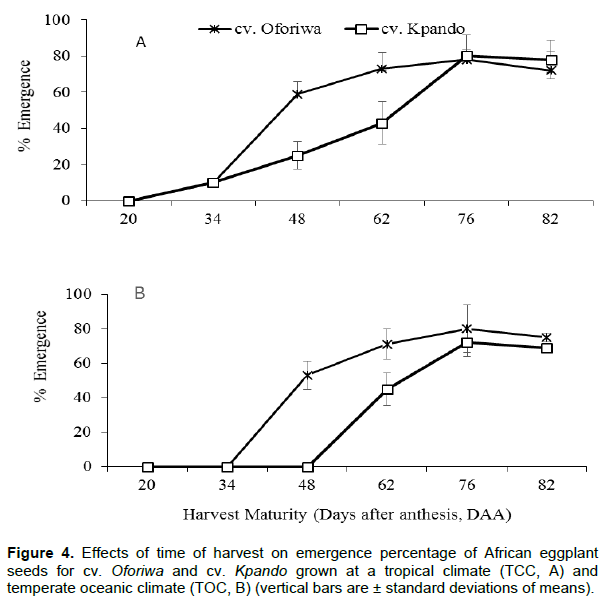
Seedling emergence tests were performed on the two cultivars produced under the two varied climatic conditions and showed significant difference between cultivars and environments (p<0.001) (Figure 4). From the tests, it was revealed that at early harvests, although seeds had acquired the ability to germinate (Figure 3), emergence was very low regardless of the moisture content. In particular, seeds of cv. Kpando obtained from the temperate oceanic climatic (TOC) conditions (Figure 4B), did not emerge until third harvest (48 DAA) and increased gradually thereafter until it reached maximum emergence of 72% at 76 DAA and marginally declined after. The pattern of seed emergence was however significantly higher in cv. Oforiwa than cv. Kpando between 48 and 76 days after anthesis at both environments. Seeds produced under the tropical conditions (TCC) (Figure 4A), however emerged after 34 DAA and increased up to 76 DAA and declined slightly. At both production environments and among the cultivars, emergence percentage was the highest when fruits were harvested at 76 days after anthesis.
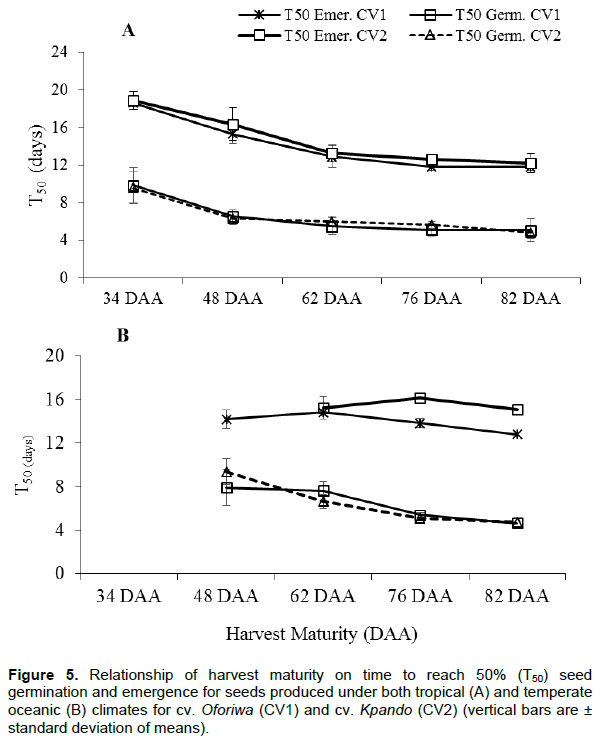
Relationship of harvest maturity on time to reach 50% germination and emergence
The time to reach 50% seed germination (T50 Germ.) under controlled conditions significantly differed (p < 0.01) from those under field conditions and during seed development. The trend was similar under both tropical and temperate oceanic conditions (B). Under standard germination test conditions, seeds harvested for cv. Oforiwa (CV1) at 34 DAA took 9.9 days to attain 50% seed germination (T50 Germ) and declined linearly to 5 days at 76 DAA (Figure 5A), where percentage seed germination and emergence were maximum (Figures 3A, B and 4A, B), respectively. It however, took 9 more days (18.6 days) for the seeds of the same maturity stage (34 DAA) to attain 50% emergence (T50 Emer.) (Figure 5A and B). Similar observations were made for cv. Kpando (CV2) under both climatic conditions (Figure 5A and B). Under both climatic conditions, cv. Kpando (CV2) however, took between 1 and 3 days more to reach 50% emergence compared to cv. Oforiwa (CV1) (Figure 5A and B).
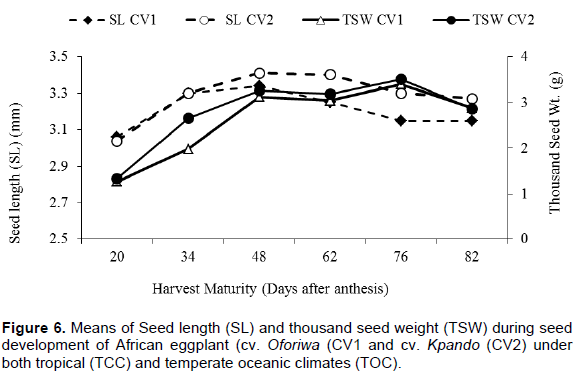
Influence of harvest maturity on seed physical attributes (seed length and weight)
Seed length (mm) and thousand seed weight (TSW) of African eggplant followed similar trend during development for both cultivars (CV1 and CV2) under both growing environments. The means of both seed length and thousand seed weight and each harvest maturity stage for both seed production environments is as shown in Figure 6. Figure 6 illustrates the changes in seed length and thousand seed weight of developing seeds of both cultivars. Seed length and thousand seed weight increased from the 20th day after anthesis, reaching a maximum at 48 and 76 DAA, respectively (Figure 6). Subsequently, both seed length and thousand seed weight declined marginally, reflecting the rapid loss of moisture that occurs after physiological maturity is reached.
Changes in fruit morphological characters (diameter, weight and seeds per fruit) during development and maturation
During fruit development and maturation, fruit width/diameter and fruit weight increased until 48 and 76 DAA, respectively and thereafter declined steadily for both cultivars and under both climates (Figure 7). For cv. Oforiwa, fruit diameter was maximum after day 48 when seeds were physiological matured and maintain same up to 76 DAA before declining. Fruit weight, on the other hand reached maximum at 76 DAA and declined slightly thereafter at both climates A and B. It was however, observed that both fruit width and weight for cv. Oforiwa produced under the tropical climate (A) significantly (p < 0.01) differed from those produced under the temperate oceanic conditions (B).
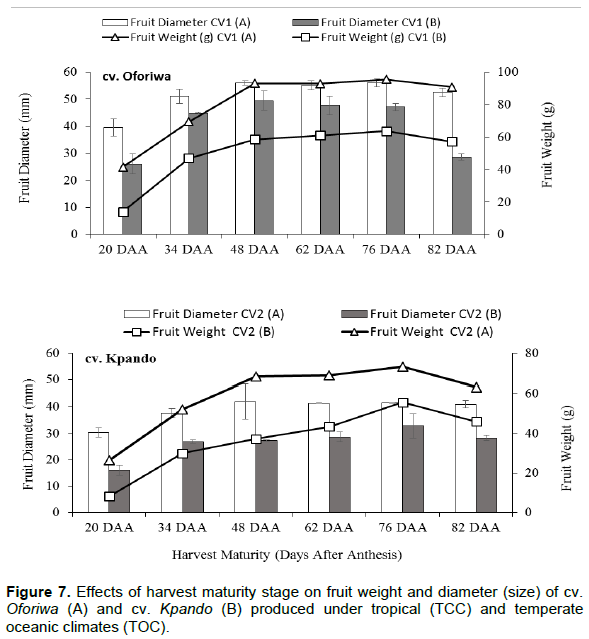
The increase in both fruit weight and width significantly differed (p < 0.01) for both cultivars at different development stages and seed production environments (Figure 7). Fruits produced under the tropical climates (A) had heavier fruit weights and larger width compared to those produced under the temperate climates (B) for cv. Oforiwa and cv. Kpando (Figure 7). Closely coinciding with the maximum fruit weight and width was the number of seeds per fruit (Figure 8A and B). Larger and heavier fruits had significantly more seeds per fruit (cv. Oforiwa) compared to fruits of cv. Kpando (Figure 8A and B) under both production environments. It was observed that significantly (p <0.01) more seeds per fruits was recorded under tropical climates (Figure 8A) than under temperate climates (Figure 8B).
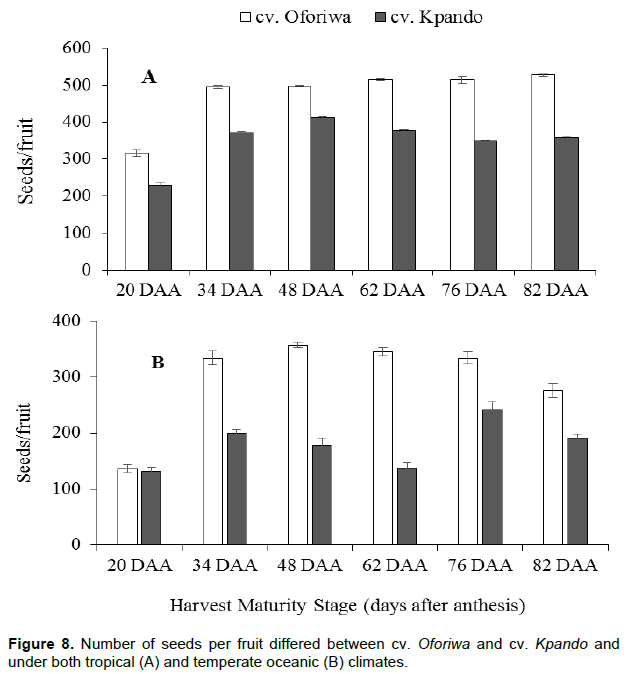
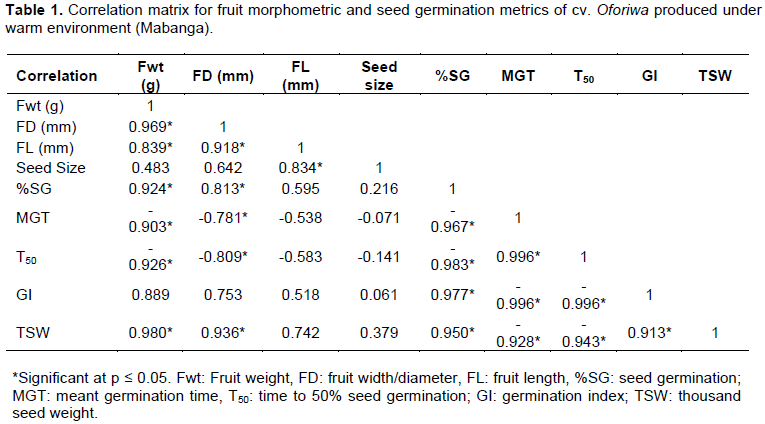
Table 1 illustrates a multiple correlation to evaluate the degree of association among the fruit dimensions and seed germination parameters of African eggplant at different maturity stages produced under both tropical and temperate climates. Strong positive correlations were found between fruit weight and percentage seed germination (r = 0.92), between fruit diameter (size) and seed germination (r = 0.81) and between seed weight (TSW) and seed germination (r = 0.95). There was however, a moderate positive correlation (r = 0.59) between fruit length and seed germination. Mean germination time (MGT) and time to 50% (T50) germination were negative and significantly related to seed germination (r = -0.96) and (r = -0.98), respectively seed germination, suggesting that seed takes less time to germinate as it seed matures (Table 1).
DISCUSSION
One important feature of seed development in fleshy fruited vegetables is that seed moisture content in these crops remains at values of about 26 to 45% (Borges et al., 2019; Ramos et al., 2021) and as high as 56% even at end of maturity (Marcos-Filho et al., 2018). In this study, seed moisture contents were 57 and 42.5% at physiological maturity of cv. Oforiwa and cv. Kpando, respectively and declined linearly, following the pattern of typical orthodox seeds (Bewley et al., 2013). The moisture content of seeds that develop in fleshy fruits generally fluctuates and remains high during the entire maturation period and even after accumulation of maximum dry matter (Vidigal et al., 2009). High seed moisture content is expected shortly after fertilization and declines steadily during development until seed reaches physiological maturation and at a moisture level suitable for harvest probably indicating that physiological maturity of seeds has been attained as observed in the present study. This observations is consistent with results for similar fleshy fruited vegetables such as tomato (Borges et al., 2019), okra (Marcos-Filho et al., 2018), pepper (Ruiz and Parera, 2017), and Capsicum baccatuum L. (Silva et al., 2015).
In fleshy fruited vegetables, seed moisture contents decline steadily while dry matter accumulates (seed dry weight increase) until it reaches maximum dry weight (physiological maturity or mass maturity) and remains at high values thereafter. The present study showed similar trend (Figure 1A and B). Maximum seed dry weight was attained at 62 DAA for cv. Oforiwa while cv. Kpando maximum seed dry weight was at 76 DAA, which is 14 days more and remained unchanged until the last harvest. Consequently, the seed moisture content progressively decreased but remained high at last harvest (82 DAA). Series of transformations occur during fruit development and maturation, including fruit tissue degradation, accumulation of sugars and organic acids (Carrari et al., 2006), leading to reduced water potential (Schumann et al., 2014). Towards the end of the maturation process however, seeds inside the fruit lose water slowly until the osmotic equilibrium is reached, which explains the lower moisture content of seeds extracted fruits harvested between 62 and 82 DAA. The gradual dehydration observed in seed moisture as maturation advanced is attributed to a decrease in the metabolism of the seed from a limit state of moisture in its tissues, an event reported as part of a natural mechanism in seeds of orthodox species (Leprince and Buitinik, 2010). Similarly, seed moisture content of eggplant decreased steadily from 53% when dry matter was maximum to about 45% at 50 DAA and then remained unchanged (Demir et al., 2002). In pepper, Ruiz and Parera (2017) observed a decline of seed moisture to 41% at physiological maturity, PM.
Closely coinciding with maximum seed dry weight was thousand seed weight (Figure 6). It has been proposed that seeds reach physiological maturity when dry seed weight is maximal (TeKrony and Egli, 1997) and water content is considerably low (Bewley et al., 2013). The loss in water content is due to the accumulation of sugars and organic acids, and plant/fruit-to-seed nutrient transfer which tend to increase the seed dry weight and thousand seed weight maturation phase (Santos et al., 2020). Similar observations have been made for tomato (Borges et al., 2019), okra (Santos et al., 2020), aubergine (Vidigal et al., 2009), and safflower (Ramos et al., 2021).
Thus, taking into account these three factors, seed moisture, dry seed weight and thousand seed weight, it can be reported that African eggplant seeds attain physiologically maturity at 62 DAA for cv. Oforiwa and 76 DAA for cv. Kpando. Vidigal et al. (2011) also reported that mass maturity or physiological maturity of sweet pepper (Capsicuum annuum) seeds occurred at 75 DAA. The differences observed for the two cultivars regarding the maturity stage to physiological maturity could be attributed to their genetic differences. Days to first flowering and fruit set in cv. Kpando takes 10-12 and 5-7 more days, respectively compared to cv. Oforiwa.
The physiological quality in terms of percentage seed germination and emergence significantly improved with maturation (Figures 3 and 4). Seeds harvested early (20 and 34 DAA) depending on the production environment had already presented germination capacity, although in low percentages. Percent germination however increased from 48 DAA, consistent with results by Martins et al. (2012) for eggplant and C. baccatuum (Silva et al., 2015), although percentage emergence was low in the present results. However, in okra, seed germination commenced as early as 30 DAA recording 44% (Santos et al., 2020). Harrington (1972) had hypothesized and later supported by Ellis (2019) that maximum seed quality was attained at physiological maturity (PM), after which deterioration started and seed germination and vigour declines. This view has been observed for okra (Demir and Ermis, 2005; Bortey and Dzomeku, 2016), selected S. aethiopicum spp. (Tetteh et al., 2021) and A. edulis fruits (Kaiser et al., 2016). In okra (Demir and Ermis, 2005; Bortey and Dzomeku, 2016), maximum germination and percentage emergence occurred at 30 DAA and coincided with maximum seed dry weight. In the case of Solanum accessions, Tetteh et al. (2021) observed maximum seed germination (86 - 94%) when 100 seed weights were maximum (0.284 - 0.62 g). Thus, the present results for cv. Kpando was consistent with the hypothesis that “maximum seed quality is reached at physiological maturity (maximum seed dry matter)” which in this case occurred at 76 DAA.
In contrast, cv. Oforiwa did not attain maximum germinability until after 14 days under tropical climates (Figure 3A) and 28 days under temperate climate (Figure 3B) after physiological maturity. This suggests that maximum germination for cv. Oforiwa occurred days after physiological maturity (PM). This phenomenon of maximum seed germination and PM not coinciding is also consistent with the views by Martins et al. (2012) and Santos et al. (2020), particularly for fleshy fruited species such as tomato, pepper, okra and eggplant. This could be attributed to genetic difference in respect of their growth habits and seed characteristics. Seed of cv. Kpando at maturity were heavier and bigger compared to cv. Oforiwa. This suggests that cv. Kpando had both a greater rate and greater duration of seed filling than cv. Oforiwa. Consequently, there is likely to be more food reserves accumulation in cv. Kpando and more time to attain physiological maturity which synchronizes with maximum germination and emergence compared to cv. Oforiwa. The explanation concurs with Zanakis et al. (1994), who observed similar cultivar difference in attainment of physiological majority among three soybean. It can however be reported that, regardless of the seed production environment and cultivar, the African eggplant seeds attain maximum germinability (that is, percent germination and emergence) when seeds were harvested 76 days after anthesis. Silva et al. (2015) and Santos et al. (2020) obtained similar results, in a study with seeds of C. baccatum L. and okra, respectively, that best seed quality was found for fruits harvested between 60 and 70 DAA. In tomato, Borges et al. (2019) concluded harvesting at 70 DAA gave highest germination quality after attaining PM at 60 DAA. Ruiz and Parera (2017) reported maximum germination for pepper at 9 weeks after anthesis (WAA) when PM had already occurred at 8 WAA. In the case of Safflower, Ramos et al. (2021) observed just 3 day interval between (36 - 39 days after flowering) between maximum seed germination and occurrence of PM. Recently, Santos et al. (2020) reported that okra attains PM at 30 DAA but maximum germination occurs 20 days after (50 DAA).
All fruit attributes (that is, diameter, weight and length) recorded significant increases (p < 0.05) between the first harvest stage (20 DAA) and the third harvest stage 48 DAA, when fruit diameter was maximum for both cultivars. While fruit diameter maintained up to 76 DAA and thereafter declined, fruit weight continued to increase marginally and peaked at 76 DAA under both conditions. The increase in values recorded for fruit diameter and weight as maturity progresses could be an indication of increase in the accumulation of assimilates during fruit maturation. This is in agreement with reports by Raz et al. (2001) and Bentsink and Koornneef (2008) that as an embryo undergoes maturation, there is food reserve accumulation resulting in fruit size and weight increase. While it can be said that the maximum fruit size for both cultivars was attained at 48 DAA as no further increase was observed in diameter, fruit weight however increased. This could indicate dry matter accumulation which resulted only in weight increase from this stage. This further point to the fact that there is usually an increase in sink strength of the fruit at this stage of development resulting in an enhanced transport of nutrients towards the fruit until maturation (Marcelis, 1996; Tanksley, 2004). Recent studies by Meng et al. (2020), has reported a linkage of increase cell numbers and endogenous IAA content to fruit size of cucumber during development. However, same cannot be justified in the present study.
The observed decline in fruit weight and size after 76 DAA reflects a possible rapid loss of moisture that occurs after physiological maturity. These figures show that both fruit weight and diameter were reduced with the maturation process as a function of water loss resulting from shrivelling of fruits during the latter stage of maturity when the rains had ceased. Similar results were reported by Silva (2002) with the fruit size of Cnidosculus phyllacantus reaching maximum values 53 days after flowering (DAF) while Physalis angulata L. from the Solanaceae family also reached maximum fruit weight and diameter at 35 DAA (Santiago et al., 2019). In S. melongena L., Kortse et al. (2017)recorded maximum fruit diameter and weight at 35 days after anthesis.
The present study further established a strong positive correlation between the fruit morphological metrics and seed physiological traits (Table 1). A strong correlation between fruit weight, diameter (size) and percentage seed germination and thousand seed weight was found to be statistically significant (p < 0.05). This observation suggests that the size and weights of fruit can be related to the seed weight (mass) resulting from the accumulation of food reserves as there is enhanced transport of nutrients towards the fruit and subsequent transfer to the seed during maturation (Tanksley, 2004). Consequently, seeds with heavier weights (mass) are likely to possess enhanced seed vigour and germinability. It has been reported that distinct seed sizes (mass) have different levels of starch and other energy reserves which influence the expression of germination and initial growth of seedlings (Shahi et al., 2015). Thus, germination depends on the ability of the seed to use reserves more efficiently (Bewley et al., 2013) by mobilization of seed reserves for the germination traits.
CONCLUSION
Considering all the fruit morphological traits and seed quality variables, the results of this study of the African eggplant (S. aethiopicum L.) under both tropical and temperate oceanic climates have shown that maximum seed quality (that is germination, emergence, thousand seed weight and mean germination time) occurs in fruits harvested at 76 days after anthesis for both cultivars. This stage coincides with physiological maturity for cv. Kpando. Cultivar Oforiwa, however attains physiological maturity at 62 DAA before achieving maximum germination 14 days later. It is recommended that for the purpose of seed production, fruits should be harvested 76 DAA for maximum seed quality.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Bentsink L, Koornneef M (2008). Seed Dormancy and Germination. The Arabidopsis Book. American Society of Plant Biologists, pp. 18-18. |
|
|
Bewley JD, Bradford KJ, Hilhorst, HWM, Nonogaki H (2013). Seeds: physiology of development, germination and dormancy. 3 Ed. New York: Springer. pp. 85-131. . |
|
|
Borges SRS, Silva PP, Araújo FS, Souza FFJ, Nascimento WM (2019). Tomato seed image analysis during the maturation. Journal of Seed Science 41(1):022-031. |
|
|
Bortey HM, Dzomeku BM (2016). Fruit and seed quality of okra [Abelmoschus esculentus (L.) Moench] as influenced by harvesting stage and drying method. Indian Journal of Agricultural Research 50(4):330-334. |
|
|
Brasil (2009). Ministério da Agricultura, Pecuária e Abastecimento - Secretaria de Defesa Agropecuária. Regras para análise de sementes. [Rules for seed testing, eggplant]. Brasília: Mapa/ACS, p. 82. |
|
|
Carrari F, Baxter C, Usadel B, Urbanczyk-Wochniak E, Zanor MI, Nunes-Nesi A, Nikiforova V, Centero D, Ratzka A, Pauly M, Sweetlove LJ, Fernie AR (2006). Integrated analysis of metabolite and transcript levels reveals the metabolic shifts that underline Tomato fruit development and highlight regulatory aspects of metabolic network behavior. Plant Physiology 142(4):1380. |
|
|
Cutti L, Kulckzynski SM (2016). Treatment of Solanum torvum seeds improves germination in a batch-dependent manner. Pesquisa Agropecuária Tropical 46(4):464-469. |
|
|
Demir I, Ellis RH (1992). Changes in seed quality during seed development and maturation in tomato. Seed Science Research 2(2):81-87. |
|
|
Demir I, Ermis S (2005). Effect of harvest maturity and drying method on okra seed quality. Seed Technology 27:81-88. |
|
|
Demir I, Mavi K, Sermenli T, Ozcoban M (2002). Seed Development and Maturation in Aubergine (Solanum melongena L.). Gartenbauwissenschaft 67(4):148-154. |
|
|
Ellis RH (2019). Temporal patterns of seed quality development, decline, and timing of maximum quality during seed development and maturation. Seed Science Research 29(2):135-142. |
|
|
Harrington JF (1972). Seed storage longevity. In: Kozlowski TT, editor. Seed Biology. Academic Press, New York 3:145-245. |
|
|
Jaetzold R, Schmidt H, Hornetz B, Shisanya C (2006). Ministry of Agriculture Farm Management Handbook of Kenya VOL. II-Part C Subpart C1. Nairobi, Kenya: Ministry of Agriculture. |
|
|
Kaiser DK, Malavasi MM, Malavasi UC, João Alexandre Lopes Dranski JA, Freitas LCN, Kosmann CR, Andrioli KK (2016). Physiological maturity of seeds and colorimetry of the fruits of Allophylus edulis [(A. St.-Hil., A. Juss. & Cambess.) Hieron. ex Niederl.]. Journal of Seed Science 38(2):092-100. |
|
|
Khalid F (2018). Advanced Germination Measurement Tool, AGRON Info-Tech. Available at: https://agronexcel.blogspot.com/2018/06/this-tutorial-is-about-advanced-seed.html |
|
|
Köppen W, Volken E, Brönnimann S (2011). The thermal zones of the earth according to the duration of hot, moderate and cold periods and to the impact of heat on the organic world (Translated from: Die Wärmezonen der Erde, nach der Dauer der heissen, gemässigten und kalten Zeit und nach der Wirkung der Wärme auf die organische Welt betrachtet, Meteorol Z 1884, 1, 215-226). Meteorologische Zeitschrift 20(3):351-360. |
|
|
Kortse PA, Oketa A, Apaa F (2017). Effects of Stage or Fruit Harvesting and After-Ripening on the Seed Quality of Garden Egg (Solanum Melongena L.). Journal of Agriculture and Veterinary Science 10(9):10-145. |
|
|
Leprince O, Buitink J (2010). Desiccation tolerance: from genomics to the field. Plant Science 179(6):554-564. |
|
|
Linkies A, Leubner-Metzger G (2012). Beyond gibberellins and abscisic acid: how ethylene and jasmonates control seed germination. Plant Cell Reports 31:253-270. |
|
|
Marcelis LFM (1996). Sink strength as a determinant of dry matter partitioning in the whole plant. Journal of Experimental Botany 47:1281-1291. |
|
|
Martins DC, Vilela FKJR, Guimarães M, Gomes LAA, da Silva PA (2012). Physiological maturity of eggplant seeds. Revista Brasileira de Sementes 34(4):534-540. |
|
|
Meng H, Xiaoxiao L, Yupeng P, Liu C, Yuanyuan D, Wang X, Z Cheng Z (2020). Cucumber Fruit Size and Shape Variations Explored from the Aspects of Morphology, Histology, and Endogenous Hormones. Plants 772(9):1-17. |
|
|
Osei MK, Banful B, Osei CK, Oluoch MO (2010). Characterization of African Eggplant for Morphological Characteristics. Journal of Agricultural Science and Technology 4(3):33-37. |
|
|
Ramos AR, Bassegio D, Nakagawa J, Maurício Dutra Zanotto MD (2021). Harvest times and seed germination of three safflower genotypes. Ciência Rural, Santa Maria 51(5):1-7. |
|
|
Raz V, Bergervoet JH, Koornneef M (2001). Sequential steps for developmental arrest in Arabidopsis seeds. Development 128:243-252. |
|
|
Ribalta FM, Pazos-Navarro M, Nelson K, Edwards K, Ross JJ, Bennett RG, Munday C, W Erskine W, Sergio JO, Croser JS (2017). Precocious floral initiation and identification of exact timing of embryo physiological maturity facilitate germination of immature seeds to truncate the lifecycle of pea. Plant Growth Regulation, pp. 1-9. |
|
|
Ruiz MB, Parera CA (2017). Effect of harvesting time on seed quality of two bell pepper cultivars (Capsicum annuum). Revista de la Facultad de Ciencias Agrarias pp. 67-77. |
|
|
Santiago WR, Gama JSN, Torres SB, Bacchetta G (2019). Physiological maturity of Physalis angulata L. seeds. Revista Ciência Agronômica 50(3):431-438. |
|
|
Santos RF, Gomes-Junior FG, Julio Marcos-Filho J (2020). Morphological and physiological changes during maturation of okra seeds evaluated. Scientia Agricola 77(3):1-9. |
|
|
Schumann C, Schlegel HJ, Grimm E, Knoche M, Lang A (2014). Water potential and its components in developing sweet cherry. Journal of the American Society for Horticultural Science 139(4):349-355. |
|
|
Shahi CV, Vibhuti KB, Bargali SS (2015). How seed size and water stress affect the seed germination and seedling growth in wheat varieties. Current Agriculture Research Journal 3(1):60-68. |
|
|
Silva LMM (2002). Maturação fisiológica de sementes de Cnidosculus phyllacanthus Pax & K. Hoffm. 61f. Tese (Doutorado em Agronomia) - Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, p. 24. |
|
|
Silva MIL, Voigt EL, Grangeiro LC, Cunha EE (2015). Determination of harvest maturity in Capsicum baccatum L. seeds using physiological and biochemical markers. Australian Journal of Crop Science (11):1010-1015. |
|
|
Singkaew J, Miyagawa S, Wongs-Aree C, Vichitsoonthonkul T, Sokaokha S, Photchanachai S (2017). Season, Fruit Maturity and Storage affect on the physiological quality of F1 Hybrid 'VTM580' Tomato seeds and seedlings. The Horticulture Journal 86(1):121-131. |
|
|
Takac A, Popovic V, Glogovac S, Dokic V, Kovac D (2015). Effects of fruit maturity stages and seed extraction time on the seed quality of eggplant (Solanum melongena L.). Ratarstvo Povrtarstvo 52(1):7-13. |
|
|
Tanksley SD (2004). The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. The Plant Cell 16:S181-S189. |
|
|
TeKrony DM, Egli DB (1997). Accumulation of Seed Vigour During development and Maturation. In: Basic and Applied Aspects of Seed Biology (Ed: R.H. Ellis, M. Black, A.J. Murdoch and T. D. Hong), Kluwer Academic Publishers, Boston, pp. 369-384. |
|
|
Tetteh R, Aboagye LM, Boateng SK, Darko R (2021). Seed quality of six eggplant cultivars as influenced by harvesting time. Journal of Applied Horticulture 23(1):24-27. |
|
|
Tetteh R, Aboagye LM, Darko R, Osafo EA (2018). Effect of maturity stages on seed quality of two tomato accessions. African Crop Science Journal 26(2):237. |
|
|
Vidigal DS, Dias DC, Von Pinho ER, Dias LA (2009). Sweet pepper seed quality and lea-protein activity in relation to fruit maturation and post-harvest storage. Seed Science and Technology 37(1):192-201. |
|
|
Vidigal DS, Dias DCFS, Dias LAS, Finger FL (2011). Changes in seed quality during fruit maturation of sweet pepper. Scientia Agricola 68(5):535-539. |
|
|
Weinberger K, Msuya J (2004). Indigenous vegetables in Tanzania: Significance and Prospects. Shanhua, Taiwan: AVRDC -The world vegetable center, Technical Bulletin no. 31, AVRDC Publication 04-600, p. 70. |
|
|
Yan D, Duermeyer L, Leoveanu C, Nambara E (2014). The functions of the endosperm during seed germination. Plant and Cell Physiology 55:1521-1533. |
|
|
Yogeesha HS, Upreti KK. Padmini K, Bhanuprakash K, Murti GSR (2006). Mechanism of seed dormancy in eggplant (Solanum melongena L.). Seed Science and Technology 34:319-325. |
|
|
Zanakis GN, Ellis RH, Summerfield RJ (1994). Seed quality in relation to seed development and maturation in three genotypes of soyabean (Glycine max). Experimental Agriculture 30:157-170. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


