Full Length Research Paper

ABSTRACT
Piliostigma reticulatum shrub is a native legume found in fallow areas in dry and semi-dry savanna soil and is used in intercropping systems. The aim was to understand the functioning of the rhizosphere, particularly the involvement of symbiotic and free living-N fixing bacteria. Soil extracts collected from P. reticulatum roots were sampled in two contrasting areas and endophytic bacterial communities were isolated using three trap host species (F. albida, A. bivenosa and V. seyal). Potential endophytic bacteria (PEB) were characterized by RFLP, nifH PCR and by 16S rRNA gene sequencing. The subsequent behavior of P. reticulatum was monitored in vitro by measuring leaf weight, biomass and chlorophyll content, after inoculation with PEB. This approach enabled isolation of 59 bacteria belonging to different genotypes. The most abundant genera were Cohnella (27.65%) among which 11 isolates clustered together and could represent a new species closely related to C. plantaginis. The other dominant genera were Paenibacillus (21.27%), Bradyrhizobium (14.89%) and Ensifer (8.5%). The nitrogen fixing gene (nifH) was detected in 21 strains and in particular, detected in a single isolate (PZS_S04) close to Cohnella xylanilytica. The strains PZS_S05 (Ensifer) and PZG_A18 (Cohnella) significantly increased certain parameters including shoot dry weight, shrub height at 90 days and photosynthetic activity (SPAD), compared to non-inoculated controls.The result obtained showed that soil under the influence of P. reticulatum roots harbored a specific diversity of endophytic bacteria including two free living-N fixing bacteria with the potential to improve the growth of P. reticulatum in natural conditions.
Key words: Piliostigma reticulatum, microbiology, phylogeny, Potential endophytic bacteria (PEB), nitrogen-fixing bacteria.
INTRODUCTION
The rhizosphere of legumes is a fertile zone due to the accumulation of different organic compounds released from their roots (Bowen and Rovira, 1999; Barea et al., 2005). For this reason, the soil under the influence of plant roots is very favorable for the growth and activity of microbial communities, which plays a significant role in carbon and nitrogen biogeochemical cycles (Vitousek and Howarth, 1991; Toal et al., 2000). Plant-associated bacteria can also have a beneficial impact on legume nutrition as nitrogen is biologically fixed by nitrogen-fixing symbiotic bacteria or by free living-N fixing bacteria in the rhizosphere. Nitrogen-fixing symbiotic bacteria (or rhizobia) can establish symbiotic relationships and can reduce the atmospheric nitrogen (N2) into ammonium (NH4+), which can be directly assimilated by plants. In addition, it is known that rhizobia can behave like non-symbiotic endophytes of legumes or non-legume plants such as maize, rice, cotton and wheat (Ueda et al., 1995; Engelhard et al., 2000) and can establish non-specific relationships with plants. However, the effects of the micro-environment formed by the plant rhizosphere on nitrogen-fixing symbiotic bacteria diversity remain unclear, in particular, on the subfamilies of Leguminosae such as Cercidoideae (LPWG, 2017). This new subfamily proposed by the Legume Phylogeny Working Group (LPWG) currently contains 12 genera (including Piliostigma genus) which mainly grow in tropical regions and have no root nodules (LPWG, 2017).
P. reticulatum (DC.) Hochst is a pioneer species, able to grow on degraded land, and to increase plant succession by other species. The shrub induces spatial heterogeneity of soil chemical properties in arid and semi-arid environments (Diedhiou et al., 2009). Twigs and wood fragments of P. reticulatum can also be used as dead ground cover (Diedhiou et al., 2009; Yélémou et al., 2013), to restore degraded land and improve crop yields (Bright et al., 2017). The use of P. reticulatum residues may improve the ability of soils to respond to saline conditions (Sall et al., 2015). Soil under the influence of the shrub is described in terms of islands of fertility (Wezel et al., 2000; Housman et al., 2007) due to a shift in soil microbial community diversity and enzymatic functions beneath the rhizosphere of the shrub compared to bulk soil (Diedhiou et al., 2009; Diedhiou-Sall et al., 2013). However, how microbial communities including symbiotic and non-symbiotic bacteria present in the soil under the influence of P. reticulatum contribute to the growth of this shrub remains unclear. Some non- symbiotic endophytic bacteria have several beneficial effects on host plants. During colonization of the roots or soil rhizosphere, they stimulate and promote plant growth (Bai et al., 2003; Aserse et al., 2013). Microorganisms isolated from the rhizosphere of various crops have been shown to produce phytohormones such as indole acetic acid as secondary metabolites. The different biosynthetic pathways of this hormone are already well described for some bacterial genera such as Azospirillum, Azotobacter and Paenibacillus (Shokri and Emtiazi 2010). Endophytes play an important role in the degradation of plant litter and organic pollutants, which in turn, actively increases soil fertility (Wang and Dai, 2011). Some works (James et al., 2000; Turner et al., 2013) suggest that endophytic bacteria may increase nitrogen fixation.
The aims of the study were to understand the functioning soil under the influence of P. reticulatum to: 1) evaluate the diversity of bacteria (symbiotic and non-symbiotic nitrogen fixing); and 2) study their impact on the growth of P. reticulatum plants.
MATERIALS AND METHODS
Sites and sampling of shrub
This study was performed at two climatically and environmentally contrasted sites. The first site Zone Sudano-Sahelian (ZSS) is located near Ndiassane (14°55'N-16°49' W), Senegal. The area is characterized by a Sudano-Sahelian climate with mean annual rainfall ranging from 400 to 600 mm. The second site Zone Sudano-Guinean (ZSG) is located in the village of Sare Yorobana (12°50'N-14°50'W), Senegal. This southern area is characterized by a Sudano-Guinean climate with more rain, and the mean annual rainfall reaches between 800 and 1,200 mm. At each site, three P. reticulatum shrubs ~ 1.5 m high were chosen and the soil under the shrub was sampled to a depth of 25 cm. The soil samples were composed of rhizosphere soil and extended to soil located around the roots, as soil adhering to the roots and around the roots are both under the influence of the shrub.
Trapping potential endophytic bacteria (PEB) associated with P. reticulatum.
The PEB present in the soil under the influence of P. reticulatum roots were studied using three trap host species (Faidherbia (syn Acacia) albida, A. bivenosa and Vachellia (syn Acacia) seyal) belonging to the mimosoid clade and the subfamily of Caesalpinioideae (LPWG, 2017). Firstly, the seeds of these species were scarified and the surface sterilized by soaking them in 95% (v/v) sulfuric acid for 30 min. The seeds of F. albida, A. bivenosa and V. seyal were thoroughly rinsed and soaked in sterile distilled water for 5 h, 7 h and 2 h for F. albida, A. bivenosa and V. seyal, respectively. The seeds were left to sprout in Petri dishes containing 2% agar and then incubated at 28°C for 48 h. The sprouting seeds were transferred under sterile conditions to Gibson tubes (Gibson 1963) containing a sterile Jensen nitrogen-free nutrient medium adjusted to pH 7 (Vincent 1970). The seedlings in the tubes were maintained in a growth chamber in the following conditions (at 28 ± 1°C with a 16 h day/8 h night photoperiod, relative humidity of 45 ± 5% and a 74 μmol.m−2. s−1 light intensity). After one week, the young plants were inoculated with 5 ml soil suspensions (10 g of soil in 90 ml of physiological water) influenced by P. reticulatum. Four repetitions were performed for each trap host species and four non-inoculated plants were used as controls.
Isolation of endophytic bacteria
After 45 days of growth, the nodules growing on the roots of the trap host species (F. albida, A. bivenosa, V. seyal) were harvested and enumerated. The nodules were then sterilized using a solution of HgCl2 (2% w/v) for 3 min, followed by several soakings in sterile water. The endophytic bacteria were isolated by streaking a loop-full of the crushed nodule on yeast mannitol agar (YMA) medium (Vincent, 1970) and incubated at 28°C for 24 h to 72 h. Similar morphological colonies to rhizobia were pricked out on YMA. Pure colonies were obtained after successive transplanting of single colonies on new plates.
After the strains were identified by sequencing, sub-groups of strains showing strong similarity with known bacterial species belonging to the family of Rhizobiaceae and Bradyrhizobiaceae were retested for their ability to nodulate their respective hosts, using Gibson tubes (Gibson, 1963) as described above.
DNA extraction and PCR of 16S rRNA and nifH genes
The genomic DNA of isolated bacteria was extracted using the NucleoSpin Tissue 96 kit (Macherey Nagel) according to the manufacturer’s instructions. All PCR were performed using the kit illustra Hot Start Mix RTG (GE Health care, Bucking Hampshire, UK). The 16S rRNA gene was amplified using pA (AGAGTTTGATCCTGGCTGAG) and pH (AAGGAGGTGATCCAGCCGCA) primers (Edwards et al., 1989). The reaction was carried out with 35 cycles as follows: denaturation for 15 s at 94 °C, primer annealing for 30 s at 55°C, and polymerization for 90 s at 72°C, plus a preheating phase of 5 min at 94°C and a terminal extension for 3 min at 72 °C. The nifH gene was amplified using the primers PolR (ATSGCCATCATYTCRCCGGA) and PolF (TGCGAYCCSAARGCBGACTC) (Poly et al., 2001) under the following conditions: 30 cycles each, denaturation for 60 s at 94 °C, annealing for 60 s at 60 °C, and elongation for 60 s at 72 °C. The cycles began with 95 °C denaturation for 15 min and ended with an extension phase for 10 min at 72 °C. PCR products were submitted to electrophoresis on 1% agarose gel for 30 min at 100 V and stained with ethidium bromide.
RFLP of 16S rRNA gene
The PCR products of the 16S rRNA gene were digested with two restriction endonucleases (Fast Digest enzyme) HaeIII and MspI (Thermo Fisher Scientific). For each PCR product, digestion was carried out in a volume of 20 μl containing 0.5 μl of enzyme, 2 μl of buffer, 5 μl of PCR products and 12.5 μl of water, and the mixtures were incubated at 37 °C for 30 min. Electrophoresis was performed on 2% agarose gel for 60 min at 100 V and stained with ethidium bromide. The results were analyzed by comparing bands, which enabled the clustering of profiles according to the position and size of bands.
Taxonomic identification of endophytic bacteria
The PCR products of 16S rRNA gene of isolates were sequenced (GATC Biotech, Constance, Germany). The nucleotide sequences were verified and corrected using SeqMan Pro (DNASTAR - Software for Molecular Biology - Sequence Analysis) and aligned with Clustal W Multiple alignment (Thompson et al., 1994). The resulting sequences were blasted against reference sequences in the NCBI database (http://www.ncbi.nlm.nih.gov/Blast.cgi). The evolutionary history was inferred using the neighbor-joining method (Saitou and Nei 1987). Evolutionary analyses were conducted in MEGA6 (Tamura et al., 2013). The percentage of replicate trees in which the associated taxa were clustered together in the bootstrap test (1000 replicates) was shown next to the branches. All sequences were submitted to NCBI with accession numbers ranging from KY992880 to KY992923.
Growth of P. reticulatum in in vitro conditions
The seeds of P. reticulatum were scarified in sulfuric acid for 1 h, then rinsed several times and soaked for 24 h in sterile water. The seeds were grown on 2% agar at 28°C for 48 h and then transferred into Gibson tubes. The seedlings in tubes were maintained in a growth chamber under the conditions described above. After 5 days, the seedlings were inoculated with 1 ml of bacterial culture (approximately 109 cfu ml−1) of each of the seven PEB. Five replicates were made for each strain and five non-inoculated plants were used as controls. The level of free-nitrogen nutritional plant was readjusted regularly in the tubes.
After three months of growth, the height of the plants was measured and chlorophyll content of the leaves was measured with a SPAD-502 chlorophyll meter (SPAD-502, Minolta Corp.; Ramsey, NJ, USA). Shoots and roots were harvested separately and dried at 70 °C to constant dry weight, after which dry weight was measured.
Statistical analyses
Statistical analyses were performed on the different growth parameters to investigate the effect of each treatment. ANOVA was followed by comparison with controls using Dunnett’s test. The multcomp package (Hothorn et al., 2008) was used with one-sided. Dunnett tests (strains tested had positive effects on these growth parameters).
RESULTS
Capture of PEB from soil under the influence of P. reticulatum
Soil under the influence of P. reticulatum sampled from the two contrasting sites contained different numbers of nodules depending on the three trap plants (F. albida, A. bivenosa and V. seyal). A total of 140 nodules were recovered at Sudano-Sahelian site (ZSS), of which 83 nodules came from the roots of F. albida, 43 nodules from the roots of V. seyal and only 14 from the roots of A. bivenosa. At the Sudano-Guinean site, a greater number of nodules (412) formed on the roots of the three trap plants. A total of 337 nodules formed on the roots of F. albida, 74 nodules on the roots of V. seyal and only one nodule formed on roots of A. bivenosa. Next, 10 nodules were sub-sampled on each trap plant to isolate the PEB, except for A. bivenova from the Sudano-Guinean site. Morphological screening under a magnifying glass enabled the selection of 59 PEB including 26 from the site. Sudano-Sahelian site and 33 from the Sudano-Guinean. The 16S rRNA gene PCR was performed on the 59 PEB. RFLP profiles revealed 14 different clusters (I to XIV) based on the number and size of the bands (Table 1). Cluster I was the most abundant (34%) and was found with three plant hosts. Cluster II was the second biggest group with 18% of profiles most isolated from nodules of F. albida. The profile of cluster IV was characteristic of the Sudano-Guinean site and isolated from nodules of V. seyal. The other groups were poorly represented, regardless of the host plant or the origin of the soil.
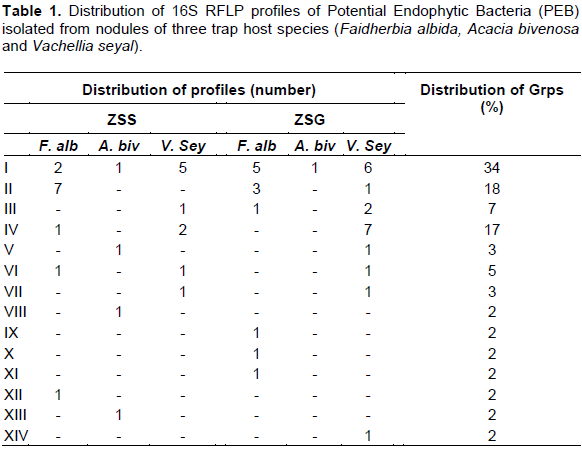
The PCR of nifH gene was also performed on 59 PEB, of which 35.6% (21 isolates) showed a positive amplification signal (Table 2), with nine positive signals of PEB at the Sudano-Sahelian site and 12 at the Sudano-Guinean site. According to the three trap host species used, the 21 nifH genes were distributed as follows: 15 positive signals (71.4%) were obtained with V. seyal; four positive signals (19%) with F. albida and only two positive signals (9.5%) with A. bivenosa.
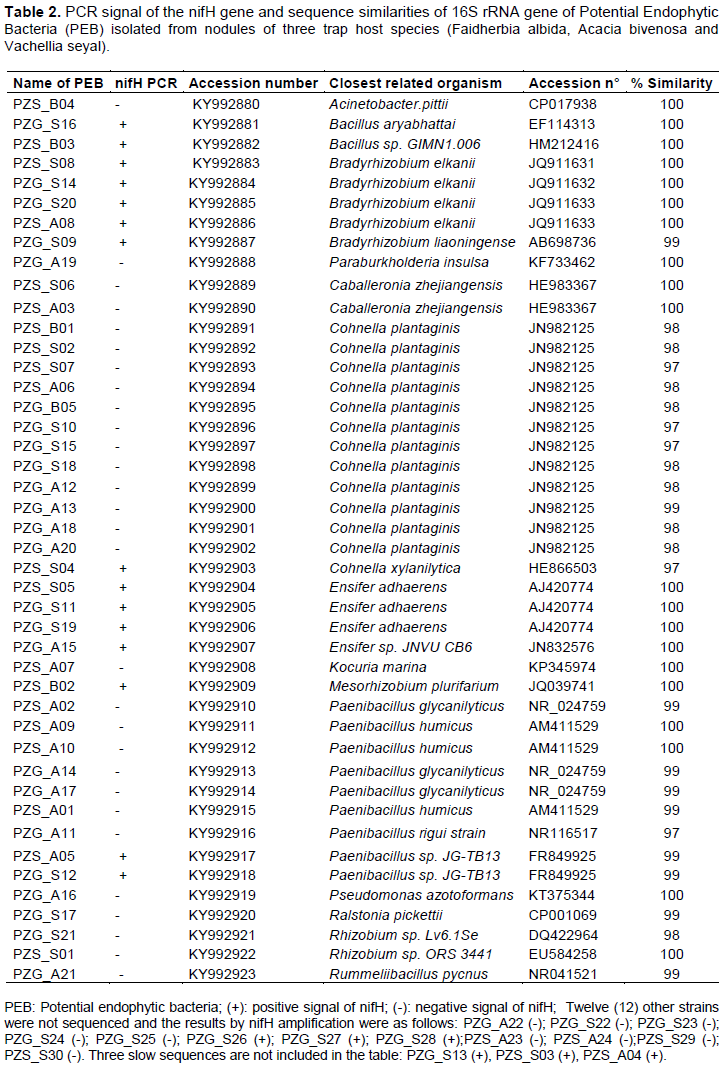
Taxonomic diversity of potential endophytic bacteria (PEB)
The results (Table 2) revealed 25 different species of PEB distributed in four clusters (Figure 1). These PEB belong to nine (9) families: Paenibacillaceae, Bacillaceae, Rhizobiaceae, Phyllobacteriaceae, Bradyrhizobiaceae, Moraxellaceae, Pseudomonadaceae, Burkholderiaceae and Micrococcaceae.
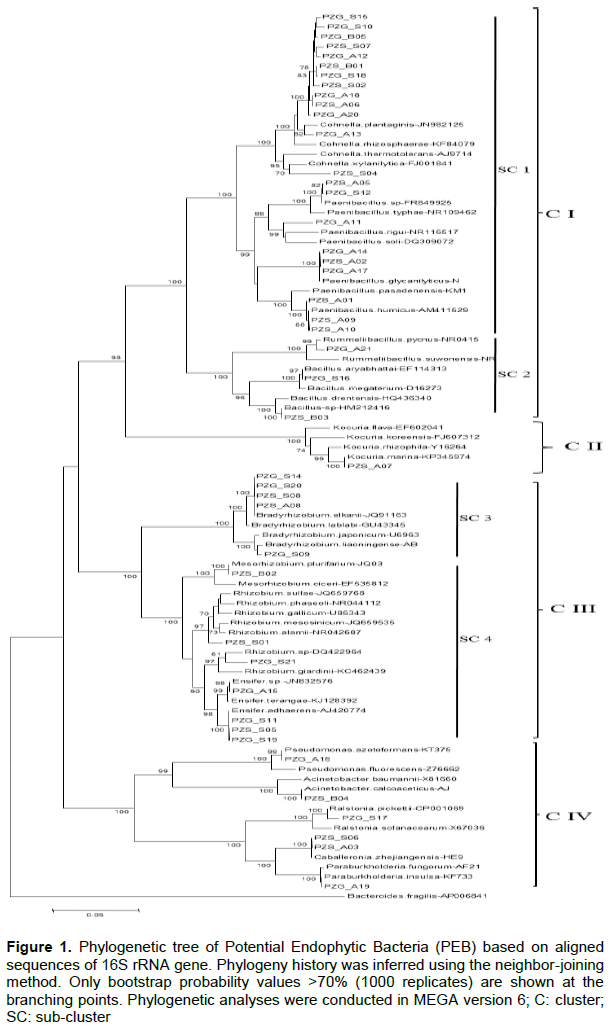
Cluster I included 25 strains and was separated in two sub-clusters (SC1 and SC2). The SC1 group contained 22 strains with strong similarity to the Paenibacillaceae family. Thirteen (13) strains were closely related to the Cohnella genus, most of which (11) were grouped in a subgroup closely related to Cohnella plantaginis isolated from plantain in China (Wang and Dai 2011). One strain (PZG_A13) was close to C. rhizosphaerae isolated from the rhizosphere environment of Zea mays (Kämpfer et al., 2015) and one (PZS_S04) formed a set with C. xylanilytica (Khianngam et al., 2010). Nine strains were closely related to Paenibacillus glycanilyticus (Kajiyama et al., 2002), to P. humicus (Vaz-Moreira et al., 2007), and to P. rigui, isolated from a freshwater wetland (Baik et al., 2011). The SC2 grouped three strains, including two strains (Bacillus aryabhattai and Bacillus sp strains (HM212416.1)), closely related to the Bacillus genus and one strain close to Rummeliibacillus suwonensis. These strains were isolated from different environments; Bacillus aryabhattai was isolated from the rhizosphere region of Lemna sp of East Kolkata wetlands of the Indian sub-continent (Ray et al., 2012), Bacillus sp strains (HM212416.1) and Rummeliibacillus suwonensis isolated from soil collected in a mountain area of South Korea (Her and Kim, 2013).
Cluster II contained a single isolated strain (PZS_A07), which was closely related to Kocuria marina sp, which is a novel Actinobacterium isolated from marine sediment (Kim et al., 2004).
Cluster III contained 12 strains separated in two sub-clusters (SC3 and SC4). Five strains were grouped in the SC3 cluster with almost 100% similarity with Bradyrhizobium elkanii (Kuykendall et al., 1992) and Bradyrhizobium liaoningense isolated from the root nodules of soybean (AB698736). The SC4 grouped seven fast growing strains, which were close to Mesorhizobium plurifarium (de Lajudie et al., 1998), to Rhizobium giardinii sp isolated from Phaseolus vulgaris nodules (Amarger et al., 1997) and to Ensifer adhaerens (Casida Jr, 1982).
Cluster IV contained six strains, which were close to different bacterial species: Pseudomonas azotoformans (Iizuka and Komagata 1963), Acinetobacter calcoaceticus (AJ888983), Ralstiona pickettii (Ralston et al., 1973), Caballeronia zhejiangensis (HE983367), and Paraburkholderia insulsa (KF733462).
Impact of PEB on the growth of P. reticulatum
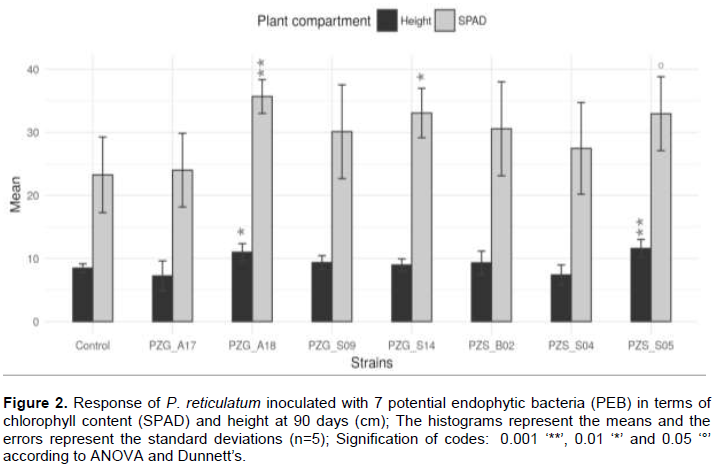
In order to evaluate the impact of PEB isolated from soil under the influence of P. reticulatum, seedlings of the shrub were inoculated with seven strains (PZG_A17, PZG_A18, PZG_S09, PZG_S14, PZS_B02, PZG_S04 and PZS_S05), mainly chosen on the basis of the presence of the nifH gene (Table 2). After three months, P. reticulatum shrubs inoculated with the three strains significantly increased the chlorophyll contents of leaves measured by SPAD (Figure 2), compared to the inoculated shrub. These were strains PZG_A18 close to Cohnella, PZG_S14 close to Bradyrhizobium elkanii and to a lesser extent, PZS_S05 close to Ensifer (p < 0.001, p < 0.01 and p < 0.05 respectively). Analyses of plant height showed that (Figure 2) the endophytic bacteria PZS_S05 and PZG_A18 significantly improved (p < 0.001 and p < 0.01, respectively) the growth of the shrub, compared to controls.
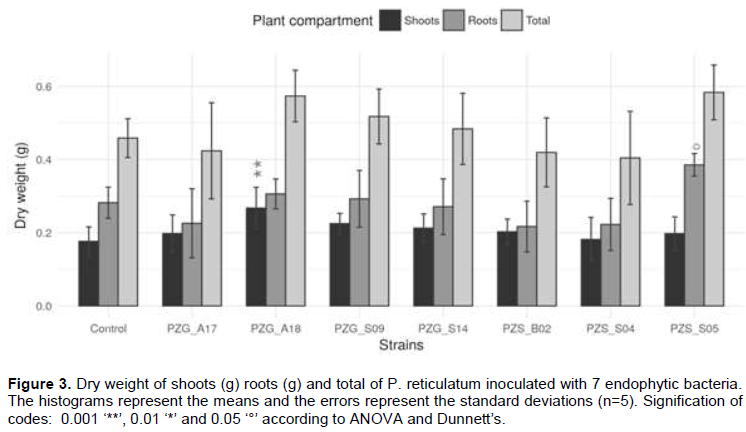
In terms of shoot dry weight (Figure 3), only the endophytic bacteria PZG_A18 differed significantly (p < 0.001) from inoculated plants. The dry weight of roots (Figure 3) revealed no significant difference between the inoculated plants and the controls, except for PZS_S05, which increased root weight (p < 0.05). In terms of total dry weight, no significant difference was observed between inoculated plants and controls (Figure 3). In total, the strains PZS_S05 close to Ensifer and PZG_A18 close to Cohnella induced a significant increase in certain parameters including dry weight of roots, growth at 90 days and photosynthetic activity, compared to the inoculated shrub.
DISCUSSION
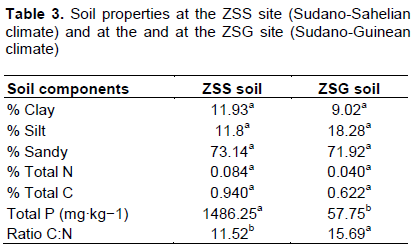
The use of three host trap species (F. albida, V. seyal and A. bivenosa) provided access to the diversity of potential endophytic bacteria (PEB) present in the soil under the influence of P. reticulatum roots in two contrasting areas (one Sudano-Sahelian and one Sudano-Guinean). Indeed, the number of nodules formed on the roots of host plants showed that the soil from Sudano-Guinean site induced more nodules than the soil from Sudano-Sahelian site. These differences seem to be due mainly to the contrasting soil properties (Table 3) at the two sites. In fact, soils from the two regions of Senegal used in this study are both sandy, but the soil from the Sudano-Guinean site had a higher percentage of silt and clay than the soil from the Sudano-Sahelian site. Similar results were found using Senegalia senegal inoculated with soils from arid and semi-arid regions of Senegal (Bakhoum et al., 2014) and A. saligna inoculated with soils from northern and southern areas of Algeria (Amrani et al., 2010). As expected, the diversity of RFLP profiles was also higher in the soil from Sudano-Guinean site than the soil from Sudano-Sahelian site.
On the other hand, the results showed that the number of nodules formed with F. albida was higher than the number obtained with A. bivenosa and V. seyal regardless of the origin of the soil. The largest number of taxonomic groups was obtained on V. seyal host despite the fact that the same number of nodules was sub-sampled. This is in agreement with the fact that V. seyal belongs to the group of plant species that can be nodulated by the Ensifer genus (Sinorhizobium) (Odee et al., 2002; Romdhane et al., 2005; Cordero et al., 2016; Sankhla et al., 2017) and Mesorhizobium (Diouf et al., 2007). In contract, A. bivenosa and F. albida are frequently nodulated by the Bradyrhizobium genus (Odee et al., 2002; Perrineau et al., 2012; Sprent et al., 2017). Even though the trap plants are selective with respect to soil bacteria, they have the advantage of selecting endophytic bacteria, including Rhizobia, more easily.
The results showed that the isolated bacterial belonged to nine families: Paenibacillaceae, Bacillaceae, Rhizobiaceae, Phyllobacteriaceae, Bradyrhizobiaceae, Moraxellaceae, Pseudomonadaceae, Burkholderiaceae and Micrococcaceae. More than 60% of these bacterial species were denoted non-symbiotic bacteria. A similar study carried out by Burbano et al. (2015) found fewer families of bacteria associated with Colophospermum mopane. As P. recticulatum, Colophospermum mopane also belongs to the family of non-nodulate plants (LPWG, 2017). Thus, the trap species used in this study clearly revealed the presence of symbiotic and nitrogen-fixing bacteria (Mesorhizobium, Bradyrhizobium and Rhizobium) in soil under influence of P. reticulatum. In addition, all the strains identified as belonging to the large group of rhizobia were able to form nodules on roots of their respective host plants (data not shown). This shows that P. reticulatum is unable to form visible nodules, but could influence the bacterial diversity of soil. This is in agreement the work of Diedhiou et al. (2013) who used phospholipid fatty acid analysis (PLFA) of microbial communities and also reported that Gram-positive bacteria (non-symbiotic) were in the majority in the soil under the influence of P. reticulatum.
To study the growth of P. reticulatum in in-vitro conditions, nitrogen fixing genes among the isolated strains were investigated. Among nitrogenase proteins, the nifH sub-unit is the most conserved. In this study, the screening of 59 PEB showed that a little less than half the isolates produced positive signals to nifH gene amplification, including almost endophytic bacteria belonging to the rhizobia group (except PZS_S01, Rhizobium sp). The absence of amplification of the latter strain may be due to a sequence divergence between the primers used and the gene present in the genome.
Strains closely related to Cohnella species were the most abundant endophytes sampled. The majority of these isolates clustered together with a high bootstrap value and was closely related to C. plantaginis. This cluster could represent a new Cohnella species (Figure 4). All the strains in this sub-cluster showed negative signals for the nifH gene. However, Wang et al. (2012) recently reported Cohnella plantaginis to be a novel nitrogen-fixing species, but by using the acetylene reduction assay. In contrast to this sub-cluster, PZS_S04, similar to Cohnella xylanitytica (97%), showed positive amplification of the nifH gene. C. xylanitytica is a xylan-degrading bacterium and was recently proposed as a new Cohnella species (Khianngam et al., 2010). It is known that many members of the Bacillus and Peanibacillus genera are diazotrophic bacteria. This is certainly the case of strains PZS_B03, PZG_S16 (belonging to the Bacillus genus), PZS_A04 and PZG_S12 (belonging to the Peanibacillus genus). In fact, the Bacillus genus also comprised a high percentage of endophytic bacteria associated with the roots of Colophospermum mopane (Burbano et al., 2015).

Comparison of the results of this study and those of Burbano et al., showed that PEB from P. reticulatum were more diversified than those associated with the roots of C. mopane and Cohnella sp. was not found associated to with roots of C. mopane. The different approaches used in the two studies (direct isolation or functional approach) could explain these differences.
In addition, the presence of nitrogen-fixing species suggests that these species could be involved in the supply of nitrogen to the plant, as well as enriching the soil with nitrogen. These groups are frequently encountered in the nodules of legumes (Zakhia et al., 2006). Their role in the nodules remains obscure, but they probably play a role in biological fixation in the nodules and/or beneficial effects via production of plant hormones that promote plant growth. In all, seven PEB were selected and inoculated on young plants of P. reticulatum. Compared to controls, PZS_S05 (Ensifer sp.) and PZG_A18 (Cohnella sp.) were able to significantly increase the height of the shrub. These two same strains also significantly increased chlorophyll contents, as is the case for PZG_S14 strain. Regarding dry weight, only strains PZG_A18 and PZS_S05 significantly increased dry shoot and root dry weight, respectively, but to a lesser extent. The mechanisms involved in these significant increases in plant growth should now be investigated, although many studies have shown that non-symbiotic endophytic bacteria can been used as inoculants to promote plant growth, nodulation, and to increase yields (Bai et al., 2003; Liu et al., 2010; Stajkovic et al., 2011). Stajković and collaborators (2011) also showed that shoot dry weight and nitrogen content in common bean plants were improved after co-inoculation of Rhizobium phaseoli and Bacillus sp. On the other hand, it appears that the increase of chlorophyll (SPAD) increased dry weight of leaves for PZG_A18, whereas in contrary the dry weight of the roots is increasend with PZS_S05. The current state of our knowledge on this shrub does not know the mechanism involved, which seems not related to bacterial strains. It will be interesting to test these strains in field conditions and to measure their impact on the growth of associated millet. As recently shown by Diakhaté and collaborators (2016) P. reticulatum appears to promote greater diversity of microorganisms in the root zone of cereal millet.
CONCLUSION
P. reticulatum is an important shrub for soil fertilization. Thus knowledge of bacterial diversity and their impacts on growth and nitrogen status of this shrub could help make better use of P. reticulatum in the arid and semi-arid zones of the Sahel. The results of this study revealed that the soil under the influence of P. reticulatum is associated with greater bacterial diversity, the extent of which varies depending on its area of origin (northern or southern Senegal).
The methods used including trapping bacteria, PCR- RFLP and 16S rRNA gene sequencing efficiently demonstrated that P. reticulatum exerts an impact on the bacterial communities of the soil. The strains PZS_S05 (Ensifer) and PZG_A18 (Cohnella) were able to improve P. reticulatum growth and increase chlorophyll content. These results pave the way for the use of the endophytic bacteria / P. reticulatum association to improve the growth of the shrub.
ACKNOWLEDGEMENTS
This work was carried out with the support of the French Embassy in Senegal (SCAC), PAPES (Support Project for the Promotion of Researchers and Teachers-researchers from Senegal), WAAPP (West African Agriculture Productivity Program), USAID / ERA (United States Agency for International Development / Education Research in Agriculture). The authors appreciate the Ampere laboratory of Ecole Centrale de Lyon (France), and the joint laboratory of microbiology (LCM) of Dakar. They are also grateful to Dr. Abdoulaye Soumaré, Omar Touré, Paul Tendeng and Mathieu Ndigue Faye for their technical help.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Amarger N, Macheret V, Laguerre G (1997). Rhizobium gallicum sp. nov. and Rhizobium giardinii sp. nov., from Phaseolus vulgaris nodules. International Journal of Systematic and Evolutionary Microbiology 47:996-1006. |
|
|
Amrani S, Noureddine N-E, Bhatnagar T, Argandona M, Nieto JJ, Vargas C (2010). Phenotypic and genotypic characterization of rhizobia associated with Acacia saligna (Labill.) Wendl. in nurseries from Algeria. Systematic and Applied Microbiology 33:44-51. |
|
|
Aserse AA, Räsänen LA, Aseffa F, Hailemariam A, Lindström K (2013). Diversity of sporadic symbionts and nonsymbiotic endophytic bacteria isolated from nodules of woody, shrub, and food legumes in Ethiopia. Applied microbiology and biotechnology 97:10117-10134. |
|
|
Bai Y, Zhou X, Smith DL (2003). Enhanced soybean plant growth resulting from coinoculation of Bacillus strains with Bradyrhizobium japonicum. Crop Science 43:1774-1781. |
|
|
Baik KS, Lim CH, Choe HN, Kim EM, Seong CN (2011). Paenibacillus rigui sp. nov., isolated from a freshwater wetland. International Journal of Systematic and Evolutionary Microbiology 61:529-534. |
|
|
Bakhoum N, Le Roux C, Diouf D, Kane A, Ndoye F, Fall D, Duponnois R, Noba K, Sylla SN, Galiana A (2014). Distribution and diversity of rhizobial populations associated with Acacia senegal (L.) Willd. provenances in senegalese arid and semiarid regions. Open Journal of Forestry 4136. |
|
|
Barea J-M, Pozo MJ, Azcon R, Azcon-Aguilar C (2005). Microbial co-operation in the rhizosphere. Journal of experimental botany 56:1761-1778. |
|
|
Bowen GD, Rovira AD (1999). The rhizosphere and its management to improve plant growth. In: Advances in agronomy. Elsevier,pp. 1-102. |
|
|
Bright MBH, Diedhiou I, Bayala R, Assigbetse K, Chapuis-Lardy L, Ndour Y, Dick RP. (2017) .Long-term Piliostigma reticulatum intercropping in the Sahel: Crop productivity, carbon sequestration, nutrient cycling, and soil quality. Agriculture Ecosystems and Environment 242:9-22. |
|
|
Burbano CS, Grönemeyer JL, Hurek T, Reinhold-Hurek B (2015). Microbial community structure and functional diversity of nitrogen-fixing bacteria associated with Colophospermum mopane. FEMS Microbiology Ecology 91:fiv030. |
|
|
Casida Jr LE (1982). Ensifer adhaerens gen. nov., sp. nov.: a bacterial predator of bacteria in soil. International Journal of Systematic and Evolutionary Microbiology 32:339-345. |
|
|
Clustal W (1994) improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice Thompson, Julie D.; Higgins, Desmond G.; Gibson, Toby J. Nucleic Acids Research 22:4673-4680. |
|
|
Cordero I, Ruiz-Díez B, de la Pe-a TC, Balaguer L, Lucas MM, Rincón A, Pueyo JJ (2016). Rhizobial diversity, symbiotic effectiveness and structure of nodules of Vachellia macracantha. Soil Biology and Biochemistry 96:39-54. |
|
|
de Lajudie P, Willems A, Nick G, Moreira F, Molouba F, Hoste B, Torck U, Neyra M, Collins MD, Lindström K (1998). Characterization of tropical tree rhizobia and description of Mesorhizobium plurifarium sp. nov. International Journal of Systematic and Evolutionary Microbiology 48:369-382. |
|
|
Diakhate S, Gueye M, Chevallier T, Diallo NH, Assigbetse K, Abadie J, Diouf M, Masse D, Sembène M, Ndour YB (2016). Soil microbial functional capacity and diversity in a millet-shrub intercropping system of semi-arid Senegal. Journal of Arid Environments 129:71-79. |
|
|
Diedhiou S, Dossa EL, Badiane AN, Diedhiou I, Sene M, Dick RP (2009). Decomposition and spatial microbial heterogeneity associated with native shrubs in soils of agroecosystems in semi-arid Senegal., Pedobiologia 52:273-286. |
|
|
Diedhiou-Sall S, Dossa EL, Diedhiou I, Badiane AN, Assigbetsé KB, Samba N, Arona S, Khouma M, Sène M, Dick R (2013). Microbiology and macrofaunal activity in soil beneath shrub canopies during residue decomposition in agroecosystems of the Sahel. Soil Science Society of America Journal 77:501-511. |
|
|
Diouf D, Samba-Mbaye R, Lesueur D, Ba AT, Dreyfus B, de Lajudie P (2007). Genetic diversity of Acacia seyal Del. rhizobial populations indigenous to Senegalese soils in relation to salinity and pH of the sampling sites. Microbial ecology 54:553-566. |
|
|
Edwards U, Rogall T, Blöcker H, Emde M, Böttger EC (1989). Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Research 17:7843-7853. |
|
|
Engelhard M, Hurek T, Reinhold-Hurek B (2000). Preferential occurrence of diazotrophic endophytes, Azoarcus spp., in wild rice species and land races of Oryza sativa in comparison with modern races. Environmental Microbiology 2:131-141. |
|
|
Gibson AH (1963). Physical environment and symbiotic nitrogen fixation. Australian Journal of Biological Sciences 16:28-42 |
|
|
Her J, Kim J (2013). Rummeliibacillus suwonensis sp. nov., isolated from soil collected in a mountain area of South Korea. Journal of Microbiology 51:268-272. |
|
|
Hothorn T, Bretz F, Westfall P (2008). Simultaneous inference in general parametric models. Biometrical journal 50:346-363 |
|
|
Housman DC, Yeager CM, Darby BJ, Sanford RL, Kuske CR, Neher DA, Belnap J (2007). Heterogeneity of soil nutrients and subsurface biota in a dryland ecosystem. Soil Biology and Biochemistry 39:2138-2149. |
|
|
James EK (2000). Nitrogen fixation in endophytic and associative symbiosis. Field crops research 65:197-209. |
|
|
Kajiyama S, Kawasaki H, Yagi M, Seki T, Fukusaki E, Kobayashi A, others (2002). Paenibacillus glycanilyticus sp. nov., a novel species that degrades heteropolysaccharide produced by the cyanobacterium Nostoc commune. International Journal of Systematic and Evolutionary Microbiology 52:1669-1674. |
|
|
Kämpfer P, Martin K, McInroy JA, Glaeser SP (2015). Proposal of Novosphingobium rhizosphaerae sp. nov., isolated from the rhizosphere. International Journal of Systematic and Evolutionary Microbiology 65:195-200. |
|
|
Khianngam S, Tanasupawat S, Akaracharanya A, Kim KK, Lee KC, Lee J-S (2010). Cohnella xylanilytica sp. nov. and Cohnella terrae sp. nov., xylanolytic bacteria from soil. International Journal of Systematic and Evolutionary Microbiology 60:2913-2917. |
|
|
Kim SB, Nedashkovskaya OI, Mikhailov VV (2004). Kocuria marina sp. nov., a novel actinobacterium isolated from marine sediment. International journal of systematic and evolutionary microbiology 54:1617-1620. |
|
|
Kuykendall LD, Saxena B, Devine TE, Udell SE (1992). Genetic diversity in Bradyrhizobium japonicum Jordan 1982 and a proposal for Bradyrhizobium elkanii sp. nov. Canadian Journal of Microbiology 38:501-505. |
|
|
Iizuka H, Komagata K (1963). New species of Pseudomonas belonging to fluorescent group. (Studies on the microorganisms of cereal grains. Part V). Nippon Nogeikagaku Kaishi 37:137-141. |
|
|
LPWG (2017). A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny 66:44-77. |
|
|
Odee DW, Haukka K, McInroy SG, Sprent JI, Sutherland JM, Young JP W (2002). Genetic and symbiotic characterization of rhizobia isolated from tree and herbaceous legumes grown in soils from ecologically diverse sites in Kenya. Soil Biology and Biochemistry 34:801-811. |
|
|
Perrineau MM, Galiana A, de Faria SM, Bena G, Duponnois R (2012). Monoxenic nodulation process of Acacia mangium (Mimosoideae, Phyllodineae) by Bradyrhizobium sp. Symbiosis 56:87-95. |
|
|
Poly F, Monrozier LJ, Bally R (2001). Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Research in Microbiology 152:95-103. |
|
|
Ralston E, Palleroni NJ, Doudoroff M (1973). Pseudomonas pickettii, a new species of clinical origin related to Pseudomonas solanacearum. International Journal of Systematic and Evolutionary Microbiology 23:15-19. |
|
|
Ray S, Datta R, Bhadra P, Chaudhuri B, Mitra AK (2012) From space to Earth: Bacillus aryabhattai found in the Indian sub-continent. Biosci Discov 3:138-145. |
|
|
Romdhane SB, Nasr H, Samba-Mbaye R, Neyra M, Ghorbal M H, De Lajudie P (2005) Diversity of Acacia tortilis rhizobia revealed by PCR/RFLP on crushed root nodules in Tunisia. Annals of Microbiology 55:249. |
|
|
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular biology and evolution 4:406-425. |
|
|
Sall SN, Ndour NYB, Diédhiou-Sall S, Dick R, Chotte JL (2015) Microbial response to salinity stress in a tropical sandy soil amended with native shrub residues or inorganic fertilizer. Journal of environmental management 161:30-37. |
|
|
Sankhla IS, Tak N, Meghwal RR, Choudhary R S, Tak A, Rathi S, Sprent J, James E, Gehlot H (2017) Molecular characterization of nitrogen fixing microsymbionts from root nodules of Vachellia (Acacia) jacquemontii, a native legume from the Thar Desert of India. Plant and Soil 410:21-40. |
|
|
Shokri D, Emtiazi G (2010). Indole-3-acetic acid (IAA) production in symbiotic and non-symbiotic nitrogen-fixing bacteria and its optimization by Taguchi design. Current microbiology 61:217-225. |
|
|
Sprent JI, Ardley J, James EK (2017) Biogeography of nodulated legumes and their nitrogen-fixing symbionts. New Phytologist 215:40-56. |
|
|
Stajkovic O, Delic D, Josic D, Kuzmanović DD, Rasulić N, Knežević-VukÄević J (2011). Improvement of common bean growth by co-inoculation with Rhizobium and plant growth-promoting bacteria. Rom Biotechnol Lett 16:5919-5926. |
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution 30:2725-2729. |
|
|
Toal ME, Yeomans C, Killham K, Meharg AA (2000). A review of rhizosphere carbon flow modelling. Plant and Soil 222:263-281. |
|
|
Turner TR, Ramakrishnan K, Walshaw J (2013). Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. The ISME Journal 7:2248. |
|
|
Ueda T, Suga Y, Yahiro N, Matsuguchi T (1995). Remarkable N2-fixing bacterial diversity detected in rice roots by molecular evolutionary analysis of nifH gene sequences. Journal of Bacteriology 177:1414-1417. |
|
|
Vaz-Moreira I, Faria C, Nobre MF,Schumann P, Nunes OC, Manaia CM (2007). Paenibacillus humicus sp. nov., isolated from poultry litter compost. International Journal of Systematic and Evolutionary Microbiology 57:2267-2271. |
|
|
Vincent JM (1970). A manual for the practical study of the root-nodule bacteria. A manual for the practical study of the root-nodule bacteria Blackwell Scientific Publications, Oxford |
|
|
Vitousek PM, Howarth RW (1991). Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13:87-115. |
|
|
Wang L-Y, Chen S-F, Wang L, Zhou YG, Liu HC (2012). Cohnella plantaginis sp. nov., a novel nitrogen-fixing species isolated from plantain rhizosphere soil. Antonie van Leeuwenhoek 102:83-89. |
|
|
Wang Y, Dai C-C (2011). Endophytes: a potential resource for biosynthesis, biotransformation, and biodegradation. Annals of Microbiology 61:207-215. |
|
|
Wezel A, Rajot JL, Herbrig C (2000). Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. Journal of arid environments 44:383-398. |
|
|
Yélémou B, Yaméogo G, Zerbo L, Koala J, Ganaba S, Millogo-Rasolodimby J (2013). Adaptation of the genus piliostigma to climatic aridity in the sahel-soudanian zone: Effect of pedoclimatic factors on the root system. Asian Journal of Plant Science and Research 3:84-94. |
|
|
Zakhia F, Jeder H, Willems A, Gillis M, Dreyfus B, De Lajudie P (2006). Diverse bacteria associated with root nodules of spontaneous legumes in Tunisia and first report for nifH-like gene within the genera Microbacterium and Starkeya. Microbial Ecology 51:375-393. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


