Full Length Research Paper

ABSTRACT
The study was conducted on dry maize seed and seed subjected to sub optimal germination temperature. Four lines of maize seeds were used, two were classified as susceptible to low temperature (44 and 54) and two tolerant to low temperature (64 and 91°C). Water content and germination tests were performed at different temperatures (10 and 25°C) to evaluate the physical and physiological quality of maize seeds. For molecular study, activity of heat-resistant proteins, esterase, superoxide dismutase, catalase and α-amylase enzymes were assessed. Study of AOX1, ZmMPK5, SOD, APX, SAD, ENR and LEA genes with real time polymerase chain reaction (PCR) were carried out. AOX, SAD and LEA genes that are related as tolerant to abiotic stress showed higher expression in line 64, which was classified as tolerant to low germination temperature. The ZmMPK5 gene is associated with increased production of abscisic acid and expressed more in line 54, which was considered as susceptible to low germination temperature.
Key words: Zea mays, seed vigor, abiotic stress.
INTRODUCTION
Exposure of plants to low temperatures causes consequences for most biological processes (Allinne et al., 2009), being considered as one of the abiotic stresses that mostly affect growth, productivity and geographical distribution of agricultural crops (Laudencia-Chingcuanco et al., 2011).
Maize is one of the most cultivated cereals in the world, but its productivity is affected, among other factors, by low temperatures. In adequate moisture and temperature of 20 to 30°C, maize seedlings emerge within 4-5 days; however, in low temperature conditions, germination can take up to two weeks or more, compromising the plant stand.
Tolerance to low temperature is a multigenic and a quantitative feature, however, the number of loci that determine this characteristic is not known (Korn et al., 2008), becoming a difficult mechanism to be elucidated by the study of a single gene (Wang et al., 2011). Thus, a better understanding of the genetic basis related to cold tolerance in different maize germplasm can provide suitable information for maize breeding by marker-assisted selection (Fracheboud et al., 2004).
Regarding the seed vigor, studies with molecular techniques have been conducted to elucidate the mechanisms involved in the germination process in abiotic stress conditions such as drought stress (Zhang et al., 2014); saline stress (Wang et al., 2014; Mardani et al., 2014.); high temperature and humidity (Han et al., 2014).
Identification of genes related to characteristics of tolerance to low temperature during germination process may contribute to the development of selection techniques that can reduce the time and effort to assess genetic sources for tolerance to such adverse temperature condition.
Given the above, this work aimed to study the enzyme activities and expression of genes that are known for their importance in plant tolerance to low temperatures, in dry maize seeds and subjected to sub optimal germination temperature.
MATERIALS AND METHODS
The survey was conducted in an experimental area and in the Central Laboratory of Seeds in the Department of Agriculture of the Federal University of Lavras (UFLA), Lavras, MG, at 910 m altitude, 21°14`S latitude and 45°00W longitude. Four lines of maize seeds were used, two being classified as susceptible to low temperature (44 and 54°C) and two tolerant to low temperature (64 and 91°C). In order to evaluate the physical and physiological quality of maize seeds, water content and germination tests were conducted at different temperatures (10 and 25°C). The water content was determined by the oven-dry method at 105°C for 24 h, using two replicates of each line, according to the Rules for Testing Seeds- RAS (Brasil, 2009). Results were expressed as mean percentage (wet basis).
Temperatures of 10 and 25°C in the germination test were used (the germination test at 25°C was carried out only to identify the seed profile). For each temperature, the test was conducted with four replications of 50 seeds, where seeds were germinated in a "Germitest" paper moistened with distilled water in the ratio of 2.5 times the weight of the dry paper. The rolls were placed in plastic bags and kept in a BOD type chamber regulated at temperatures of 10 and 25°C (±3°C). Assessments of normal seedlings were performed at 4, 7, 14 and 21 days after beginning the tests. The results were expressed as mean percentage of normal seedlings from the four replications. It was considered as 'normal seedlings’, those who had at least 1 cm of taproot, with two adventitious roots of at least 1 and 1 cm shoot. Percentage of protrusion was also assessed, adopting as standard, the seedlings with at least 0.5 cm of root protrusion. The tests were performed in a completely randomized design and mean comparison test among treatments was taken by Scott-Knott test, at 5% probability, to analyze the results. Analyses were performed in the SISVAR statistical program. For proteomic (except for alpha amylase) and transcripts analyses, dry and soaked seeds at 10°C for 14 days were used.
In order to extract heat-resistance proteins, seeds were ground in a crucible with ice and liquid nitrogen, then a buffer solution was added (50 mM tris-HCL-7.5; 500 mM NaCL; 5 mM MgCl2; 1 mM PMSF) 1:10 (material weight: volume of extraction buffer) and transferred to 1500 μL capacity microtubes. The homogenates were centrifuged at 14,000 rpm for 30 min at 4°C and the supernatant was incubated in a water bath at 85°C for 15 min and then centrifuged again. The supernatant was poured into microtubes, and the pellet discarded. Before application in the gel, the sample tube containing 70 μL extract + 40 μL sample buffer (2.5 ml glycerol, 0.46 g SDS, 20 mg Bromophenol blue and the volume completed to 20 ml of Tris pH 7.5 extraction buffer) were placed in a water bath with boiling water for 5 min (BLACKMAN et al., 1991). 50 mL of protein extract + sample buffer were applied per well in the polyacrylamide gel electrophoresis SDS-PAGE (12.5% separating gel) and 6% (concentrating gel). Electrophoresis was performed at 120 V and gels were stained with 0.05% Coomassie Blue for 12 h and distained in 10% acetic acid solution (Alfenas, 2006).
For extraction of α-amylase enzyme, seeds were germinated in paper roll for 14 days, at a temperature of 10°C. After this period, seeds were cold ground in crucibles with liquid nitrogen. For extraction, 200 mg ground seed powder was suspended in 600 µL extraction buffer (0.2M Tris-HCl, pH 8.0 + 0.4% PVP). Then, there was application of 40 µL protein extract in each well in polyacrylamide gels at 7.5% (separating gel + 1% soluble starch) and 4.5% (concentrating gel). Electrophoresis was performed at 120 V for 6 h.
For catalase, esterase, peroxidase and superoxide dismutase extraction, the following were used: Tris HCL buffer 0.2 M pH 8.0 + (0.1% mercaptoethanol), 250 μL per 100 mg of seeds. The material was homogenized by vortex and kept for 12 h in a refrigerator, followed by centrifugation at 14,000 rpm for 30 min at 4°C. Electrophoresis was performed in polyacrylamide gel 7.5 (separating gel) and 4.5% (concentrating gel), 60 µL sample's supernatant was applied into the gel and electrophoresis was performed at 120 V for 5 h. Gel revelation was according to Alfenas (2006). For extraction of RNA, it was used four seed lines (44, 54, 64 and 91), dry and soaked for 14 days at a temperature of 10°C.
Maize seeds were ground with liquid nitrogen with Pure Link RNA Plant® (Invitrogen) reagent, according to manual instruction. RNA integrity was checked with 1% agarosis gel. RNA quantification was measured in a spectophotometer at 260 and 280 nm wave-length. After nucleic acid extraction, samples were treated with DNA Free DNase (Ambion) to avoid DNA contamination. Protocol was performed according to manufacturer's recommendations. After RNA extraction and purification, cDNA synthesis was performed using High Capacity cDNA Reverse Transcription cDNA® kit (Applied Biosystems), following the manufacturer's recommended protocol.
By gene expression analysis by qRT-PCR, genes were selected based on literature review due to the importance of studies on plant tolerance to low temperatures. Primers were designed using Primer Express 3.0 software (Applied Biosystems) following the search for gene sequences in http://www.ncbi.nlm.nih.gov site (Table 1). Ubiquitin gene was used as endogenous control.

For qRT-PCR, quantitative analysis of gene expression, ABI PRISM 7500 Real-Time PCR (Applied Biosystems) equipment, SYBR Green detection system and cDNA obtained from RNA extracted from the seeds were used. The thermal reaction conditions were 2 min at 50°C, 10 min at 95°C, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C, ending with 15 s at 95°C. Data were collected and stored in the 7500 software program (version 2.0.1). For each reaction, 1.0 µl cDNA diluted 1: 5, 0.2 µL of each primer and 5.0 µl SYBR green Master Mix UDG with ROX (Invitrogen) were used to obtain a final volume of 10.0 µL/sample. Negative controls and melting curves were included in all the analyses. Three biological repetitions were used in three replicates techniques for each studied gene and the results were normalized using the threshold cycle (CT) obtained by Ubiquitin reference gene expression.
The CT was determined by the number of cycles in which the fluorescence generated within a reaction crossed the baseline (threshold). The comparative CT method was used. To do so, first, a validation experiment was performed to prove that the efficiency of the target gene amplification and references were similar and very close to 100% (TYAGI; BRATU; KRAMER, 1998). Standard curves for the studied genes were generated from the following dilutions: 1: 5, 1:25, 1: 125, 1: 625 and 1: 3125. This procedure also defined the best cDNA dilutions in each reaction, which was 1: 5.
For each gene, samples of low expression were used as calibrator samples and the method used to measure the relative expression was Relative Standard Curve Method described in the instrument’s procedure handbook (Applied Biosystems). For the quantification of gene expression by real time PCR, values obtained corresponding to the samples' mRNA levels were compared to the values of the control's mRNA levels. After obtaining the raw data, they were analyzed using the 7500 SDS Software (Version 2.0.1). To calculate the expression level of the interest genes, the following was considered: Ct (PCR product exponential growth) of the target gene and the endogenous control, ΔCt = Ct (sample) – Ct (endogenous control) and the ΔΔCt = ΔCt (sample) - ΔCt (calibrator). Then the level of expression was calculated with the formula: RQ = 2-ΔΔCt. For graph plot, the SigmaPlot program was used.
RESULTS AND DISCUSSION
The average water content of seeds at the time of testwas 12.6%, with a maximum variation of 1%. Regarding the physiological quality of maize lines seeds used in this study, it could be observed that when the germination test was carried out at 25°C (which is a temperature suitable for maize germination), there was no statistical difference in germination among materials (Table 2). When seeds were germinated at 10°C temperature, stratification of lots in different vigor levels was observed (Table 2). Regarding the germination percentage, lines 44 and 54 showed no seedlings have reached the minimum standard adopted to consider them as normal. With regards to the percentage of protrusion, line 54 was lower than the other lines. This result reflects the effect of genotype on seed tolerance to low germination temperature.
Li et al. (2013), working with induction of cold tolerance in wheat seeds during germination found that when seeds were germinated at 12°C, the germination rate was well below the germination rate of the same materials at 22°C. It is expected that materials more tolerant to low-temperature germination conditions present a well-protected membrane system and an efficient antioxidant defense systems, that is, with increased expression of scavenging enzymes.
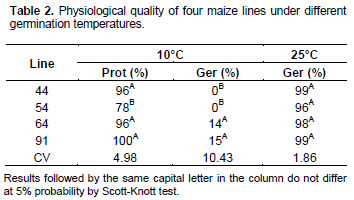
Figure 1 shows esterase enzyme activity. It can be seen in this figure that when the seeds were soaked for 14 days at 10°C, seeds from line 54 were the ones that had higher activity of esterase enzyme. The increased activity of this enzyme in these seeds may have triggered a lipid peroxidation, thus contributing to a lower tolerance of this line to cold and thus resulting in low percentages of protrusion and non-occurrence of germination (Table 2). Regarding the superoxide dismutase enzyme (Figure 1), in line 64, there was an expression of an isoform that is not expressed in other materials when seeds were submitted to immersion in stress conditions. Under stress conditions, at low temperatures, reactive oxygen species (ROS) are produced at high levels and may result in damage to DNA, proteins and lipids (Miller et al., 2008). So, the more efficient the antioxidant system, the smaller the damage caused by ROS. The greatest expression of SOD in line 64 may have contributed to this material protrude and originate normal seedlings, even in the temperature of 10°C. Low temperatures affect the germination of hybrid maize seeds because they increase the time for seeds to germinate, reduce the germination percentage and contribute to enhanced superoxide dismutase expression (Farooq et al., 2008).
Regarding the expression of catalase enzyme in the maize lines studied, it appears that for both dry and soaked seeds at 10°C, line 54 showed the lowest expression of this enzyme (Figure 1). Line 54 had a lower percentage of protrusion at 10°C, and the inefficiency of the antioxidant system may have been one of the reasons for this result. Catalase enzyme reduce expression in line 54, even in dry seeds, is an important feature, because it allows the selection of a material during the breeding programs without necessarily making the seeds to germinate.
The increased tolerance to cold may be accompanied by increased expression of specific genes encoding antioxidant enzymes (Baek and Skinner, 2003), plants with increased expression of SOD and other scavenging enzymes designed to increase stress tolerance (Xi et al., 2010). In seeds of line 44, lower expression of heat-resistant proteins (Figure 2) and high protrusion percentage was observed, however after 21 days of soaking, normal seedlings were not observed. Low expression of heat-tolerant proteins may have influenced this result, due to a decreased stability of the membrane system.
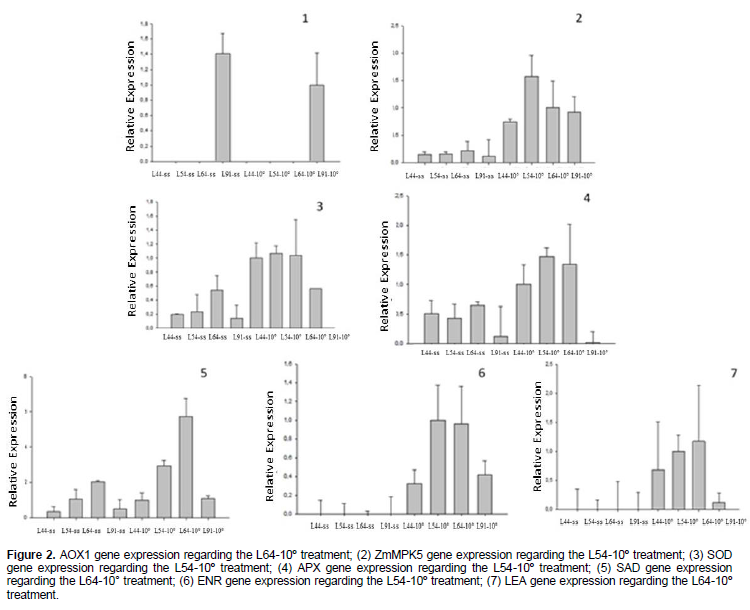
In Figure 2, it can be seen that line 64 had the highest expression of α-amylase, so this material was rated as the one with greater tolerance to cold during the germination process, however, line 91, which was also considered to be more tolerant, did not show high activity of this enzyme. These data reflect the complexity of the germination process. In seeds of two recombinant lines which were damaged in controlled conditions, it was possible to identify 65 QTLs for germination energy characteristics, germination percentage, dry root weight and dry shoot weight. However, only five of these QTLs were responsible for representing more than 10% of the phenotypic variation, with highest ratio of 11.92% (Han et al., 2014), the results of this study reinforce the complexity of the germination process. The mechanisms involved in cold tolerance have were also studied through expression of transcripts in a range of species. Figure 1 represents the alternative oxidase gene expression (AOX). It can be seen that only line 64 showed expression of this gene both for dry seed and for soaked seeds at 10°C for 14 days. The expression on dry seeds makes this gene a potential marker for tolerance to low temperature in maize seeds, because it speeds up the selection process. Surge et al. (2006), studying the Arabidopsis response to stress caused by low temperature, found that the AOX activity resulted in attenuation of ROS production. According to Karpova et al. (2002), expression of high levels of AOX in maize occurs due to respiratory failure or membrane depolarization which is responses to cold stress.
Figure 2 shows ZmMPK5 gene expression. Accumulation of ABA and hydrogen peroxide contributes to increased ZmMPK5 expression (Lin et al., 2009). The ZmMPK5 expression was higher in soaked seeds at 10°C than in dry seeds for all the studied lines. Increased expression of this gene was found in seeds soaked for 14 days at 10°C from line 54. This line presented lowest percentage of protrusion when compared with other lines and did not provide seedlings with the minimum characteristics to be considered normal. This susceptibility to low temperature may be due to accumulation of ROS and also higher abscisic acid content, as these two factors may have contributed to greater ZmMPK5 expression. Abscisic acid (ABA) is a phytohormone known to modulate growth of plants in response to stresses (Christmann et al., 2006). ABA regulates important aspects of the plant development, including the initiation and maintenance of seed dormancy. This hormone stabilizes seed dormancy to ensure that germination takes place in a suitable environmental condition (Finch-Savage and Leubner-Metzger, 2006).
Figure 2 presents the expression of superoxide dismutase. This enzyme had higher expression in the dry seeds from line 64, regarding dry seeds of other lines. But for all lines, expression was higher in seeds submitted to cold stress. Seeds from line 91 were those who had lowest expression of SOD. Despite being one of the main antioxidant enzymes, the study of a single gene is not enough to explain the cold tolerance during germination process. Even though line 91 have low SOD expression, it was considered tolerant to low temperatures. This can be explained by the fact that there are several other antioxidant enzymes that could have prevented the production or eliminated the ROS, contributing to the good performance of line 91. In tomato acclimation to cold there is an increase in the NADPH oxidase activity, increase in hydrogen peroxide levels in the cell apoplast and increase in the activity of antioxidant enzymes in an attempt to alleviate the oxidative process and maintain the stability of the membrane (Zhou et al., 2012). Another important antioxidant enzyme is ascorbate peroxidase. The expression of this enzyme is represented in the Figure 2. The expression of this enzyme was lower in line 91, when seeds were soaked at 10°C. This enzyme neutralizes peroxide through ascorbate. For seeds of other lines soaked at 10°C, the expression in the materials was similar. Figure 2 show the expression of genes related to lipid composition of membranes. In Arabidopsis thaliana, expression of desaturases (FAD8) was strongly induced by low temperature (Gibson, 1994). Liu et al. (2006) studying the effect of temperature on tomato crops verified that the expression of desaturases (LeFAD7) was induced by stress caused by low temperature (4°C), but inhibited by high temperature (45°C) in leaves. In rice, FAD2 gene seems to be related to stress resistance in plants grown under unfavorable temperature conditions (Shi et al., 2012). Kodama et al. (1995) observed that unsaturated fatty acid is one of the factors involved in tolerance to low temperatures in young tobacco leaves. Figure 2 shows the expression of putative stearoyl-ACP desaturase gene (SAD). Desaturase converts saturated fatty acid into unsaturated. In general, the SAD expression was higher in soaked seeds than in dry seeds in the four studied lines. In dry seeds, greatest expression of this gene was verified in line 64. When seeds were soaked at 10°C, the behavior was the same, and seeds of line 64 had higher SAD expression than other lines. Among the studied materials, line 64 was considered tolerant to low germination temperature. Putative enoyl-ACP reductase (ENR) gene expression is presented on Graph 6. The expression of this gene was higher in seeds soaked at 10°C than in dry seeds and when seeds were soaked, lines 54 and 64 had higher expression of this gene. Gene expression related to LEA protein (late embryogenesis abundant) is represented in the Figure 2. The lower expression of this gene was found in line 91 when seeds were soaked at 10°C. Heat tolerance proteins have been studied as important in plant tolerance to cold stress. Line 91 had high percentage of protrusion at 10°C, however, it did not have high expression of the antioxidant genes studied in this work, nor LEA proteins and desaturase. This result indicates the complexity of cold tolerance characteristics and that studies of few genes are not enough to select materials with tolerance to low temperatures during the germination process. However, this work shows that there are differences in expression between the materials and that some genes related to tolerance to low temperature are expressed more in line 64 which has also been identified as tolerant. AOX, LEA and SAD gene can be mentioned.
Another gene that also had an interesting correlation was ZmMPK5. This gene showed higher expression in line 54, which had the lowest percentage of protrusion at 10°C when compared with the other three lines. Expression of this gene is associated with ABA accumulation, and it is known that abscisic acid is a hormone that negatively affects the germination, being related to seed dormancy process.
So it is possible to suggest that research in the field of molecular analysis focused on seeds, can bring great benefits to understanding the mechanisms involved in tolerance to abiotic stresses and contribute to selection or development of tolerant materials to these stresses. However, studies must be conducted to identify a larger number of genes that can be used to select maize materials with tolerance to low germination temperature.
CONCLUSIONS
1. For some studied genes, there was correlation of increased expression with higher tolerance to low temperature. AOX, SAD and LEA genes that are related to tolerance to abiotic stress had higher expression in line 64, after materials were subjected to soaking at 10°C for 14 days.
2. ZmMPK5 gene is associated with increased production of abscisic acid and was expressed more in line 54, which was classified as susceptible to low germination temperature.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alfenas AC (2006). Eletroforese de isoenzimas e proteínas afins: fundamentos e aplicações em plantas e microorganismos. Viçosa, MG: UFV 574 p. |
|
|
Allinne C, Maury P, Sarrafi A, Grieu P (2009). Genetic control of physiological traits associated to low temperature growth in sunflower under early sowing conditions. Plant Sci. 177:349-359. |
|
|
Baek Kh, Skinner Dz (2003). Alteration of antioxidant enzyme gene expression during cold acclimation of near-isogenic wheat lines. Plant Sci. 165:1221-1227. |
|
|
Brasil (2009). Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Brasília 399 p. |
|
|
Christmann A, Moes D, Himmelbach A, Yang Y, Tang Y, Grill E (2006). Integration of abscisic acid signalling into plant responses, Plant Biol. 8:314-325. |
|
|
Farooq M, Aziz T, Cheema ZA, Hussain M, Khaliq A (2008). Activation of Antioxidant System by KCl Improves the Chilling Tolerance in Hybrid Maize. J. Agron. Crop Sci. 94:438-448. |
|
|
Finch-Savage WE, Leubner-Metzger G (2006). Seed dormancy and the control of germination. New Phytol. 171:501-523. |
|
|
Fracheboud Y, Jompuk C, Ribaut JM, Stamp P, Leipner J (2004). Genetic analysis of cold-tolerance of photosynthesis in maize. Plant Mol. Biol. 56:241-253. |
|
|
Gibson S, Arondel V, Iba K, Somerville C (1994). Cloning of a Temperature-Regulated Gene Encoding A Chloroplast Omega-3 Desaturase From Arabidopsis-Thaliana. Plant Physiol. 106(4):1615- 1621. |
|
|
Han Z, Ku L, Zhang Z, Zhang J, Guo S, Liu H, Zhao R, Ren Z, Zhang L, Su H, Dong L, Chen Y (2014). QTLs for Seed Vigor-Related Traits Identified in Maize Seeds Germinated under Artificial Aging Conditions. PLoS ONE 9(3):e92535. |
|
|
Karpova OV, Kuzmin EV, Elthon TE, Newton KJ (2002). Differential expression of Alternative Oxidase Genes in maize mitochondrial mutants. The Plant Cell 14:3271-3284. |
|
|
Kodama H, Horiguchi G, Nishiuchi T, Nishimura M, Iba K (1995). Fatty-Acid Desaturation During Chilling Acclimation Is One Of The Factors Involved In Conferring Low-Temperature Tolerance To Young Tobacco-Leaves. Plant Physiol. 107(4):1177-1185. |
|
|
Korn M, Peterek S, Mock H, Heyer AG, Hincha DK (2008). Heterosis in the freezing tolerance, and sugar and flavonoid contents of crosses between Arabidopsis thaliana accessions of widely varying freezing tolerance. Plant Cell Environ. 31:813-827. |
|
|
Laudencia-Chingcuanco D, Ganeshan S, You F, Fowler B, Chibbar R, Anderson O (2011). Genome-wide gene expression analysis supports a developmental model of low temperature tolerance gene regulation in wheat (Triticum aestivum L.). BMC Genom. 12:299. |
|
|
Li X, Jiang H, Liu F, Cai J, Dai T, Cao W, Jiang D (2013). Induction of chilling tolerance in wheat during germination by pre- soaking seed with nitric oxide and gibberellin. Plant Growth Regul. 71:31-40. |
|
|
Lin F, Ding H, Wang J, Zhang H, Zhang A, Zhang Y, Tan M, Dong W, Jiang M (2009). Positive feedback regulation of maize NADPH oxidase by mitogen-activated protein kinase cascade in abscisic acid signaling. J. Exp. Bot. 60:3221-238. |
|
|
Liu XY, Yang JH, Li B, Yang XM, Meng QW (2006). Antisense-mediated depletion of tomato chloroplast omega-3 fatty acid desaturase enhances thermal tolerance. J. Integr. Plant Biol. 48(9)1096-1107. |
|
|
Mardani Z, Rabiei B, Sabouri H, Sabouri A (2014). Identification of molecular markers linked to salt-tolerant genes at germination stage of rice. Plant Breed. 133:196-202. |
|
|
Miller G, Shulaev V, Mittler R (2008). Reactive oxygen signaling and abiotic stress. Physiol. Plantarum. 133:481-489. |
|
|
Shi J, Cao Y, Fan X, Li M, Wang Y, Ming F (2012). A rice microsomal delta-12 fatty acid desaturase can enhance resistance to cold stress in yeast and Oryza sativa. Mol. Breed. 29(3):743-757. |
|
|
Wang YY, Hecker AG, Hauser BA (2014). The APX4 locus regulates seed vigor and seedling growth in Arabidopsis thaliana. Planta 239:909-919. |
|
|
Wang Z, Wang F, Zhou R, Wang J, Zhang H (2011). Identification of quantitative trait loci for cold tolerance during the germination and seedling stages in rice (Oryza sativa L.). Euphytica 181:405-413. |
|
|
Xi DM, Liu WS, Yang GD, Wu CA, Zheng CC (2010). Seed-specific overexpression of antioxidant genes in Arabidopsis enhances oxidative stress tolerance during germination and early seedling growth. Plant Biotechnol. J. 8:796-806. |
|
|
Zhang H, Cui F, Wang H (2014). Detection of quantitative trait loci (QTLs) for seedling traits and drought tolerance in wheat using three related recombinant inbred line (RIL) populations. Euphytica 196:313-330. |
|
|
Zhou J, Wang J, Shi K, Xia XJ, Zhou YH, Yu JQ (2012). Hydrogen peroxide is involved in the cold acclimation-induced chilling tolerance of tomato plants. Plant Physiol. Biochem. 60:141-e149. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


