Full Length Research Paper

ABSTRACT
In the global agribusiness, the herbicide use is a major problem for sustainable production, in this sense, it is necessary to better understand the interaction of weed species and floristic composition such as biodiversity indicators. The objective of this study was to analyze the spatial variability of weeds in an Oxisol under no-tillage system. Samples were taken in an area of 0.5 ha, in 50 sampling points with spacing of 5 m x 10 m. Data were analyzed by means of classical statistics, geostatistics, and spatial variability of the constructed maps by the interpolation by kriging technique. All the species of weeds presented in the study area showed spatial variability with the exception of Ipomoea triloba (L.) and Heliotropium indicum (L.), which showed pure nugget effect. The range values (a) shows that the spacing between samples can be extended to all species of weeds. The study was unable to determine specifics areas of management in the local since the different species of weed infested different plots of the area.
Key words: Precision agriculture, semivariograms, site-specific management.
INTRODUCTION
MATERIALS AND METHODS



RESULTS AND DISCUSSION
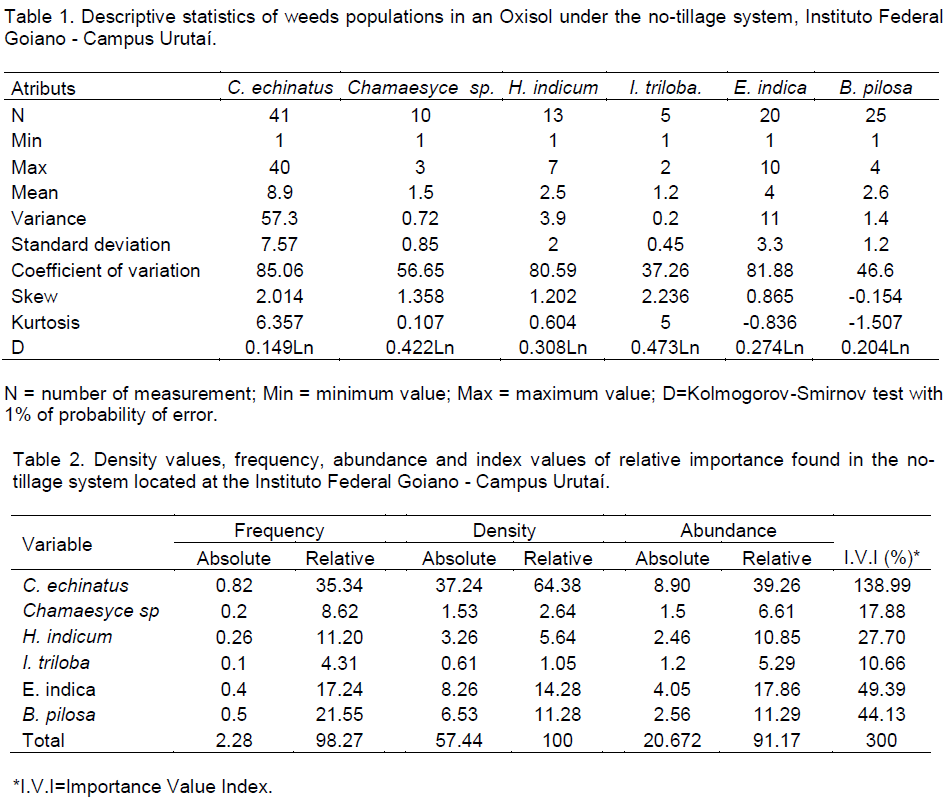
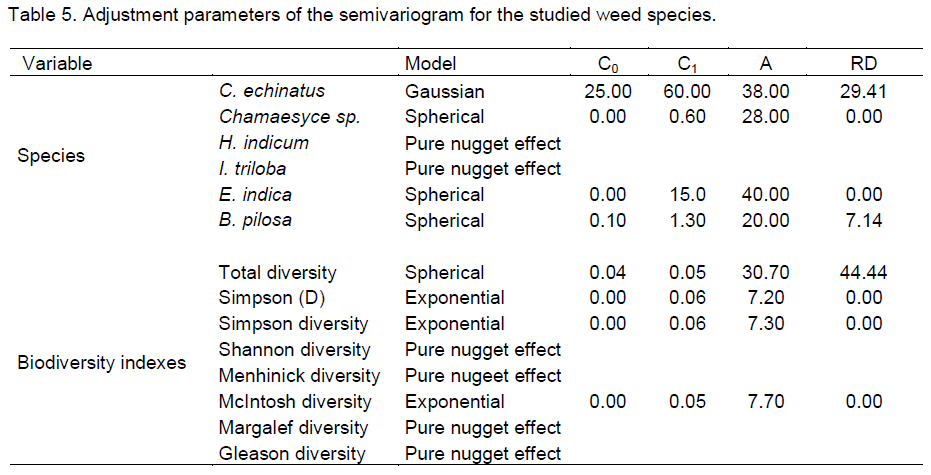
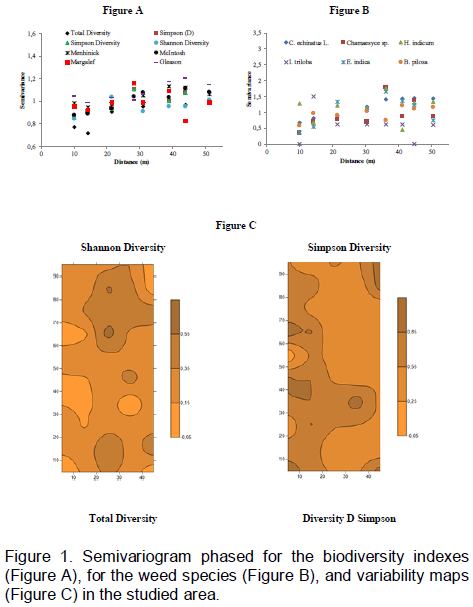
CONFLICT OF INTERESTS
ACKNOWLEDGMENTS
REFERENCES
|
Bressan GM, Koenigkan LV, Oliveira VA, Cruvinel PE, Karam D (2006). Sistema de classificação fuzzy para o risco de infestação por plantas daninhas considerando a sua variabilidade espacial. Planta Daninha 24(2):229-238. |
|
|
Calado JMG, Baschi G, Barros JFC, Carvalho M (2013). Weed management in winter wheat (Triticum aestivum L.) influenced by different soil tillage systems. 8(21):2551-2558. |
|
|
Cambardella CA, Mooman TB, Novak JM, Parkin TB, Karlem DL, Turvo RF, Konopa AE (1994). Field scale variability of soil properties in central Iowa soil. Soil Sci. Am. J. 47:1501-1511. |
|
|
Carvalho JRP, Silveira PM, Vieira SR (2002). Geoestatística na determinação da variabilidade espacial de características químicas do solo sob diferentes preparos. Pesqui. Agropecu. Bras. 37(8):1151-1159. |
|
|
Chiba MK, Filho OG, Vieira SR (2010). Variabilidade espacial e temporal de plantas daninhas em Latossolo Vermelho argiloso sob semeadura direta. Acta Sci. Agron. 32(4):735-742. |
|
|
Domingos MKR, Laca-Buendia JP (2010). Levantamento fitossociológico das plantas daninhas na pré-colheta da cultura do sorgo granífero em Uberaba/MG. FAZU Rev. 7:68-72. |
|
|
Goel PK, Prasher SO, Landry JA, Patel RM, Bonnell RB, Viau AA, Miller JR (2003). Potential of airborne hyperspectral remote sensing to detect nitrogen deï¬ciency and weed infestation in corn. Comput. Electronics Agric. 38:99-124. |
|
|
Johnson GA, Mortensen DA, Young LJ, Martin AR (1996). Parametric sequential sampling on multistage estimation of the negative binomial parameter k. Weed Sci. 44(4):555-559. |
|
|
Journel AG, Huijbregts CJ (1978). Mining geostatistics. London, Academic Press, 600 p. |
|
|
Lacerda ALS, Victoria Filho R, Mendonça CG (2005). Levantamento do Banco de Sementes em dois Sistemas de Manejo de Solo Irrigados por Pivô Central. Planta Daninha 23(1):1-7. |
|
|
Lorenzi H (2000). Manual de identificação e controle de plantas daninhas: Plantio direto e convencional. 5.ed, Nova Odessa: Plantarum. 384 p. |
|
|
Lutman PJW, Perry NH (1999). Methods of weed patch detection in cereal crops, In: The 1999 Brighton Conference – Weeds, Brighton 1999, Proceedings, Brighton: BCPC. pp. 627-634. |
|
|
Monquero PA, Amaral LR, Binha DP, Silva PV, Silva AC, Martins FRA (2008). Mapas de infestação de plantas daninhas em diferentes sistemas de colheita da cana-de-açúcar. Planta Daninha 26(1):47-55. |
|
|
Mortensen DA, Dielemann JA, Johnson GA (1998). Weed spatial variation and weed management. In: Hatfield JL, Buhlerard DD, Stewart BA (Eds.). Integrated Weed and Soil Management. Ann Arbor Press, Chelsea, MI, pp. 293-309. |
|
|
Mueller-Dombois D, Ellenberg HA (1974). Aims and methods of vegetation ecology. New York: John Wiley. 547 p. |
|
|
Rodrigues ACP, Costa NV, Cardoso LA, Campos CF, Martins D (2010). Períodos de interferência de plantas daninhas na cultura do sorgo. Planta Daninha 28(1):23-31. |
|
|
Rodrigues WC (2015). DivEs - Diversidade de Espécies v3.0 - Guia do Usuário. Entomologistas do Brasil. 33 p. 2015. Disponível em: http://dives.ebras.bio.br |
|
|
Santos CMA (2007). Estatística Descritiva – Manual de auto-aprendizagem; Edições Sílabo. |
|
|
Schaffrath VR, Tormena CA, Gonçalves ACA, Junior RSO (2007). Variabilidade espacial de plantas daninhas em dois sistemas de manejo de solo. Rev. Bras. Engenharia Agrícola Ambient. 11(1):53-60. |
|
|
Shiratsuchi LS, Molin JP, Christoffoleti PJ (2004). Mapeamento da distribuição espacial da infestação de Panicum maximum durante a colheita da cultura de milho. Planta Daninha 22(2):269-274. |
|
|
Shiratsuchi LS, Fontes JRA, Resende AV (2005). Correlação da distribuição espacial do banco de sementes de plantas daninhas com a fertilidade dos solos. Planta Daninha 23(3):429-436. |
|
|
Siqueira GM, Dafonte DJ, Valcárcel AM (2015). Correlación espacial entre malas hierbas en una pradera y su relación con la conductividad eléctrica aparente del suelo (CEA). Planta Daninha 33(4):631-641. |
|
|
Siqueira GM, Vieira SR, Ceddia MB (2008). Variabilidade espacial de atributos físicos do solo determinados por métodos diversos. Bragantia .67(1):203-211. |
|
|
USDA (1999). United States Department of Agriculture –Soil taxonomy: a basic system of soil classification for making and interpreting soil surveys. Washington. 871 p. |
|
|
Vieira SR (2000). Geoestatística em estudos de variabilidade espacial do solo. In: Novais RF, Alvarez VH, Schaefer GR (Eds.). Tópicos em Ciência do solo. Viçosa: Soc. Bras. Ciênc. Solo 1:1-54. |
|
|
Vieira SR, Tillotson PM, Biggar JW, Nielsen DR (1997). Scaling of semivariograms and the kriging estimation of field-measured properties. Rev. Bras. Ciênc. Solo .21:525-533. |
|
|
Warrick AW, Nielsen DR (1980). Spatial variability of soil physical properties in the field. In: Hillel D (Ed.). Applications of soil physics, New York: Academic Press. |
|
|
Webster R, Olivier MA (1990). Statistical methods in soil and land resource survey. Oxford, Oxford University Press. 316 p. |
|
|
Wiles LJ, Oliver GW, York AC, Gold HJ, Wilkerson GG (1992). Spatial distribution of broadleaf weeds in North Carolina soybean (Glycine max) fields. Weed Sci. 40(5):554-557. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


