High salt concentrations in soil reduce the growth and productivity of crops. Salinity effects are most pronounced in arid and semiarid regions due to droughts and high evaporation during the year (Ayers and Westcot, 1999; Tester and Davenport, 2003).
However, some crops produce economically viable yields even at high levels of soil salinity, due to the higher capacity of osmotic adaptation of these species, which allows the absorption of sufficient water even under saline conditions (Ayers and Westcot, 1999). Such adaptability is very useful and allows the selection of tolerant genotypes when it is not possible to keep soil salinity at low levels (Tester and Davenport, 2003).
Salinity tolerance varies among species and within species; its effects vary between stages of development. In general, the tolerance has been identified as specific for some development stages, that is, a genotype may be more tolerant during one stage and more sensitive during other stage (Foolad et al., 1998; Silva et al., 2014). Irrigated fruit production is considered salt sensitive and requires attention to prevent salinization, particularly when the water source has high salt concentrations.
Brazil is the third largest fruit producer in the world (FAO, 2013), therefore, it is essential to generate knowledge that enables the use of saline waters and ensures sustainability of production. Among fruit crops, citrus stand out as having the highest national importance and the sweet orange (Citrus sinensis (L) Osbeck) as having the highest production and exportation (FAO, 2013). Citrus fruits also stand out in northeastern Brazil for undoubted social and economic importance. A practice that may enable the use of low quality water and salt affected soils is the use of genotypes with high tolerance to this abiotic factor.
However, the development of cultivars tolerant to salinity by breeding programs requires the study of the effects of salinity on plant physiology to identify processes responsible for any tolerance mechanisms (Silva et al., 2014). In fruit crops propagated by grafting, such as citrus, salt tolerance should be assessed considering the scion-rootstock plant. Careful selection of both scion and rootstock is critical to the success of citrus crop, given that the main effect of salinity on plants are the suspension of growth, leaf injury symptoms and yield reduction (Syvertsen and García-Sanchez, 2014).
According to Wei et al. (2013), the investigation of possible salt tolerance mechanisms in various cultivars may obtain more insight into the role of scion in citrus to tolerate salinity. Brito (2007), while evaluating 18 citrus genotypes, highlighted some varieties as being tolerant and moderately tolerant to salinity during the period of rootstock formation and indicated individuals with potential for production of citrus seedlings under salt stress. These genotypes, however, were not studied after grafting. Therefore, in this study, the growth of different combinations of citrus scion-rootstock under saline water application by irrigation during the formation of seedlings (rootstocks ungrafted and grafted) was assessed.
Study site
Experiments were conducted under greenhouse conditions at the Center of Technology and Natural Resources - CTRN of Federal University of Campina Grande - UFCG, located in Campina Grande, state of Paraiba, Brazil, in the geographic coordinates 7°15'18"S and 35°52’28" W with an altitude of 550 m.
Treatments and experimental design
Five levels of salinity of irrigation water were applied: S1 - water with electrical conductivity (ECw) of 0.8 dS m-1; S2 - ECw of 1.6 dS m-1; S3 - ECw of 2.4 dS m-1; S4 - ECw of 3.2 dS m-1; and S5 - ECw of 4.0 dS m-1. The threshold salinity of 1.7 dS m-1 described in Ayers and Westcot (1999) for 'Pera' sweet orange was used to define these levels, having two levels below and three above this limit. The levels of electrical conductivity of water (ECw) were applied in five genotypes tolerant to salinity and six moderately tolerant genotypes that were selected by Brito (2007) in an experiment that comprised only the seedling stage (probably plants of nucelar origin), rootstock production, and used ‘Rangpur’ lime (moderately sensitive) as control (Table 1). Seeds of genotypes were provided by the Citrus Breeding Program of Embrapa Cassava and Fruits - Embrapa CBP. After the formation of rootstocks, genotypes were grafted with two scion varieties: Tahiti lime [C. latifolia (Yu. Tanaka) Tanaka], and Star Ruby grapefruit (C. paradisi Macfad.), whose buds were also provided by Embrapa CBP. Combining all factors, we had 12 rootstocks x 5 salinity levels x 2 scions, totaling 120 treatments. The experimental design was a randomized block with three replications, and each plot consisted of four plants.
Sowing and management practices
The seeds were properly selected and treated with fungicide (4 g of Thiran per kg of seeds) and sown at the rate of three per tube with a capacity of 288 ml, in commercial substrate containing a combination of vermiculite, pine bark and humus, in a proportion of 1:1:1, keeping only one seedling in each container (Schäfer et al., 2005).
Plants grown in tubes were transplanted after germination and early growth (five months), to plastic bags 35 cm high, 22 cm upper diameter and 20 cm bottom diameter. Grafting was performed after transplantation and establishment of rootstocks in the bags. Irrigation was applied in same volume, depending on the evapotranspiration in the control treatment, which was obtained by weighing bags of different blocks, adding a leaching fraction equivalent to 20% (LF = 0.2). The control of weeds and prevention of insects, as well as fertilization, were carried out as usually recommended for production of citrus (Rozane et al., 2007).
Growth measurements
Every 30 days, from the beginning of treatment until grafting [240 days after sowing (DAS)], the number of leaves (NL), plant height (PH, cm) and stem diameter (SD, mm) from rootstocks were recorded. After grafting until 330 DAS, the number of leaves in the scion (NLScion), the length of the scion’s stem (cm) measured from grafting point to stem's apex of scion, the diameter of rootstock’s stem (mm), measured near the substrate by using a digital caliper rule, and the scion’s stem diameter (mm) measured at 2 cm above the grafting point were obtained.
Statistical analysis
The factor “irrigation water salinity” was evaluated by analysis of variance using 'F' test, and polynomial regression (linear and quadratic). The Scott-Knott test was used to evaluate the factor “rootstock”, and the Tukey test was used to verify differences between scions. All tests were performed at 95% of probability (Ferreira, 2011).
Seedlings growth
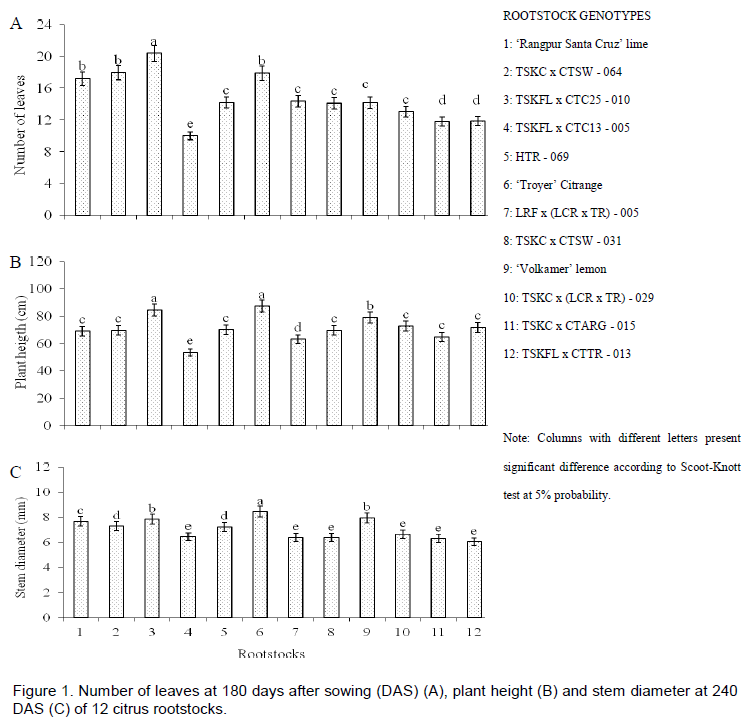
According to the Scott-Knott test, seedlings with the largest number of leaves were relatives to the hybrids TSKFL x CTC25 - 010, and had about 21 leaves at 180 DAS (Figure 1A). More than four groups of genotypes were obtained, with emphasis to TSKFL x CTC13 - 005, which appears in the group with lower mean (about 10 leaves). However, it should be noted that greater or lesser number of leaves may be related to genetic material and not just due to the potential effects of salinity. This can be verified when comparing different genotypes for this characteristic, taking as base the average salinity and the lack of significant effect on interaction, which indicates that behavior was similar among salinities.
Fochesato et al. (2007), when studying growth of citrus rootstocks on different substrates, found a higher number of leaves on ‘Rangpur’ lime and a lower number in citrange 'C13'. This last genotype is composed of the hybrid TSKFL x CTC13 - 005, which confirms the results shown in this study.
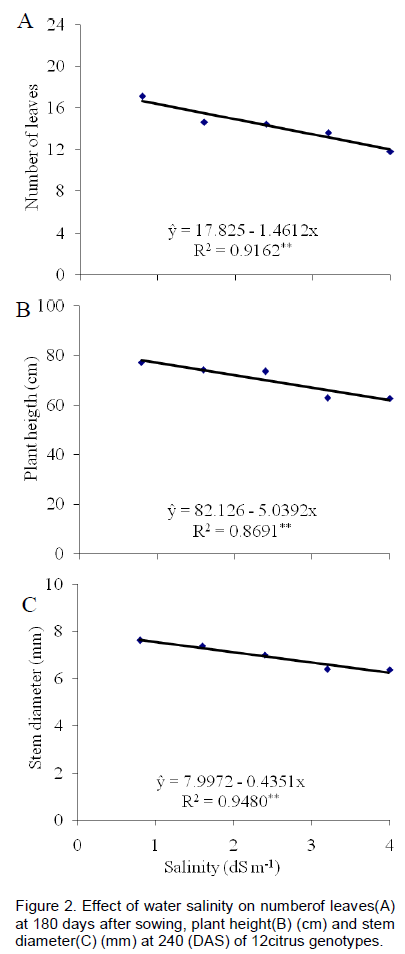
Salinity reduced the number of leaves in 8.7% at 180 DAS (Figure 2A) with unit increase in salinity of water (dS m-1), based on the lowest salinity level.
Levy and Syvertsen (2004) reported in their reviews that the absorbed chloride accumulates in leaves of citrus causing abscission and/or reducing growth. This effect may have happened in our research, which led to a reduction in the formation of leaves with increased salinity, because we did not consider the fall of leaves.
With regard to plant height, the TSKFL x CTC25 - 010 and 'Troyer' citrange stood out in the group of genotypes with the highest averages (Scott-Knott test p < 0.05; Figure 1B). The lowest average was observed in the group of the hybrid TSKFL x CTC13 - 005. Simpson et al. (2014), studying the growth response of grafted and ungrafted citrus plants under saline irrigation, did show a difference among three genotypes evaluated by them, with the best result for C146 and C22, and smaller in the sour orange (C. aurantium L.) from dates of relative growth rate in ungrafted plants, such as found in the data of this study. This can mean that the effect of salinity is variable among genotypes and these results reinforce the theory about the need to choose the best genotype for use under saline water conditions.
The height of rootstocks decreased linearly by increasing water salinity (dS m-1) (Figure 2A) to 6.4% at 240 DAS. Presence of genotypes with reduced growth under salinity conditions is indicative of materials potentials, as identified by Fernandes et al. (2011) and Silva et al. (2014). However, these low values during citrus plants in the ungrafted stage can be relative to phase, such as observed by Simpson et al. (2014); these authors did show that plants during the ungrafted stage are more tolerant than in the grafted stage. But keeping in view that stress was applied in the substrate without salinity, in other words, the increase in substrate salinity was gradual, the plants may have hardened and became resistant to saline stress, as cited by Syvertsen and García-Sanchez (2014) in their reviews about multiple stress on citrus under salinity.
The values of stem diameters at 240 DAS ranged from 6.0 to 8.5 mm. 'Troyer' citrange stood out as having the highest average values (Figure 1C). Stem diameter is one of the most important variables in study of citrus rootstocks because it indicates the plant’s potential for grafting. Genotypes with larger diameter have more success when grafted with buds. Based on the lowest level of salinity, the increase of one unit in the salinity of water also reduced the diameter by 5.6% at 240 DAS (Figure 2C). These results were similar to those obtained by Soares et al. (2006), who studied three citrus rootstocks in saline waters, in a nursery, in São Paulo, Brazil. According to Syvertsen and García-Sanchez (2014) salinity may have caused nutritional disturbances, reduced accumulation of biomass and plant growth. Moreover, salinity may cause toxic disturbances by the action of specific ions or drought stress by reducing the water potential in soil.
Plant growth after grafting
The increase of rootstock diameter after grafting may be an evidence of adaptation of scion to rootstock; in this relationship, the rootstock provides support and input of water and nutrients absorbed from soil, while the scion produces organic compounds that are transported to all organs, including roots. The best adaptation was found in ‘Rangpur’ lime under high salt concentration. Average values of ‘Rangpur’ lime ranged from 9.6 mm at the lowest level of salinity to 8.5 mm at the highest level, while in the last assessment before grafting 7.8 mm in diameter was obtained (Figure 1C). The ‘Rangpur Santa Cruz’ lime, TSKFL x CTC25 - 010, HTR - 069, 'Troyer' citrange and 'Volkamer' lemon, which had higher estimated averages, showed linear decreases in stem diameter of 3.3, 3.4, 5.7, 4.4 and 6.0%, respectively, with 1 dS m-1 increase in EC of water with respect to the lowest level of salinity (Figure 3).
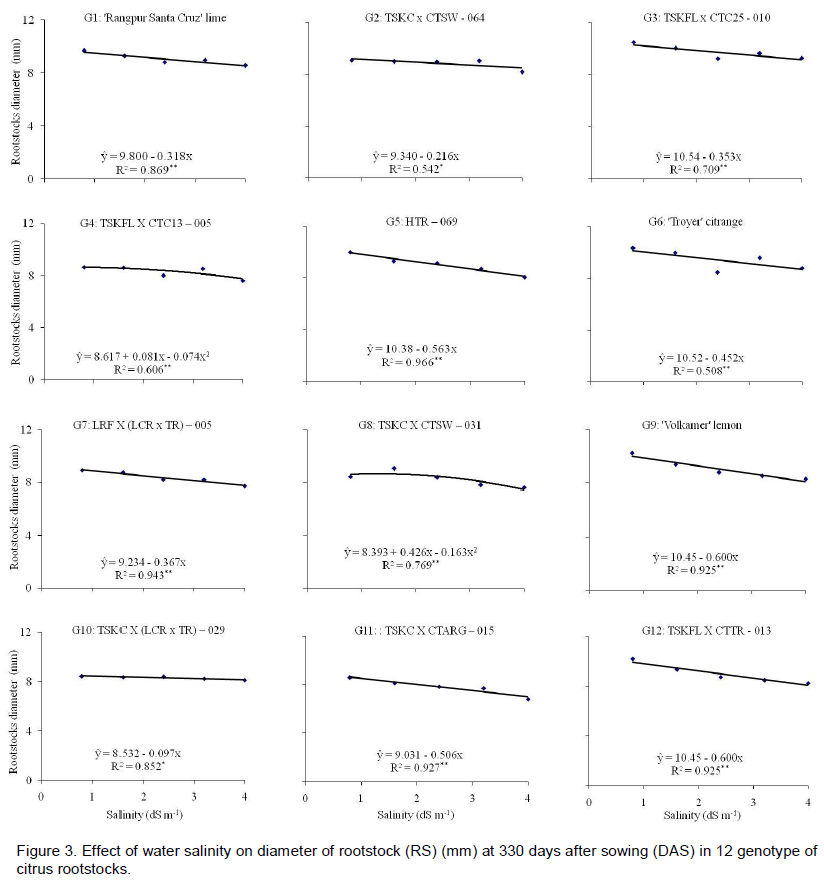
According to Fageria and Gheyi (1997), when comparing yields of material (genotype) under salt stress to the same material without stress, and the difference is less than 20%, this material is considered tolerant. Therefore, taking the rootstock’s stem diameter as a criterion for selection of genotypes, with respect of tolerance to salinity, it can be concluded that all are tolerant to salt stress. However, it is not appropriate to take only the stem diameter of rootstock as criterion.
The behavior of rootstocks on each scion genotype relative to the number of leaves is observed in Table 2 with linear and quadratic equations. 'Rangpur Santa Cruz' lime grafted under 'Tahiti' lime stood out with highest means under the lowest level of salinity (0.8 dS m-1), showing a gradual reduction and quadratic trend, with the increase in electrical conductivity of irrigation water. On the other hand, when ‘Rangpur’ is grafted with 'Star Ruby' grapefruit, a linear decrease with increase in water salinity can be noted, with reduction of about 24% between 0.8 and 4.0 dS m-1 ECw. Also, in this scion (‘Star Ruby’), other genotypes with potential can be noted, highlighting the HTR - 069 and ‘Troyer’ citrange with a reduction of 39 and 35% in the number of leaves when salinity was increased from 0.8 to 4.0 dS m-1. Thus, this may mean that it is better to choose the combination scion/rootstock to be successful in a citrus crop under salinity conditions.

Quadratic behavior was also observed in the TSKFL x CTC25 - 010, TSKFL x CTC13 - 005, LRF x (LCR x TR) - 005 and TSKC x CTSW - 031 grafted with 'Tahiti' lime and 'Star Ruby' grapefruit, and in TSKC x (LCR x TR) - 029 grafted with 'Star Ruby' (Table 2). Remaining combinations decreased the number of leaves with increasing salinity. Even in the quadratic model showing the best adjustment, salinity decreased the number of leaves. This may be related to toxic effects of ions in the water, which compete in nutrition and osmotic adjustment. Thus, the plants use more energy to maintain their photosynthetic apparatus (Taiz and Zeiger, 2009).
In Table 3, the behavior of scion/rootstock combinations for length of scion stems is shown, with a significant triple interaction (salinity x rootstock x scion). Linear decrease was observed when the rootstocks TSKC x CTSW - 064, TSKFL x CTC25 - 010, 'Troyer' citrange and TSKFL x CTTR - 013 were grafted with 'Tahiti' or 'Star Ruby'. A similar situation was observed when ‘Rangpur’ lime and the hybrid TSKFL x CTC13 - 005 were combined with 'Star Ruby', and when the hybrid LRF x (LCR x TR) - 005 was grafted with 'Tahiti'. There was a quadratic behavior for the remaining genotypes, except for the hybrid TSKC x (LCR x TR) - 029. The effect of salt promoted reduction both in number of leaves and plant height. Soares et al. (2006) and Fernandes et al. (2011) also identified a decrease in height of citrus under salt stress. These authors emphasized that growth occurs by cell division and expansion and this latter occurs by increase in cell turgor pressure by storage of water in vacuoles (Taiz and Zeiger, 2009).

Thus, salt stress reduces water uptake and the plant may have difficulties to carry out such processes, which results in growth limitations. Relative to variable stem diameter of scion, the behavior of scion-rootstock combinations under water salinity is shown in Table 4. The highest values for this parameter were found in combinations of 'Rangpur Santa Cruz' lime with 'Tahiti' and 'Star Ruby', however, this reduces with increase in salinity of irrigation water (dS m-1). Besides these combinations, the hybrid TSKFL x CTC25 - 010 grafted with 'Tahiti', as well as the hybrid TSKFL x CTC13 - 005 and ‘troyer’ citrange grafted with 'star ruby' had a decrease in stem diameter of the scion until the plants died with increasing salinity.

The remaining genotypes best fitted to the quadratic model, however, had a gradual decrease with increasing salinity. Behavior similar to ‘Rangpur’ lime combined with the scion varieties was observed in 'Volkamer' lemon, with greater stem diameter of scion in the scion-rootstocks combination "Star Ruby/Volkamer" in some salinity levels.
Another interesting combination between hybrid trifoliate (HTR - 069) as rootstock and scion of grapefruit was that except for the first salinity level (S1, 0.8 dS m-1), the estimated averages were not close to those of "Star Ruby-Rangpur" and "Star Ruby-Volkamer"; moreover, in this combination no significant reduction occurred when water up to ECw = 2.4 dS m-1 was used. Thus, for a wider range of materials to choose from, HTR-069 under scions of 'Star Ruby’ may be added to the list of best combinations, and it can use water with EC equal to 2.4 dS m-1. It also shows that using 'Star Ruby' grapefruit as scion can improve tolerance to salinity.
The rootstock has a critical role in the growth of the scion genotype. The appropriate combination of rootstock and scion genotypes is essential for plant development and success of crops, especially under conditions of irrigation with saline water. This fact is corroborated in studies carried out by Singh et al. (2003) and Syvertsen and Garcia-Sanchez (2014). Another observation in this study is that the salinity effect is more in ungrafted rootstocks, because the growth reduction was highest in the grafted phase, and death of plants was noted with
irrigation water of ECw = 4.0 dS m-1, as observed by Simpson et al. (2014).
In most combinations growth was reduced due to increased salinity, but not up to zero, which allows to affirm that salinity provokes different effects among genotypes and among phases of development. The combination scion/rootstock plays a critical role in this process. Several approaches of plant adaptation to salt stress are present in literature (Syvertsen and Garcia-Sanchez, 2014). It is believed that non-tolerant plants do their adjustment by partitioning salts in vacuoles of old leaves, as described by Taiz and Zeiger (2009). Thus, when the canopy was cut from rootstock, salts began to accumulate in the graft tissue, which become thin and, thus, fail to grow. This fact is associated to the combinations of TSKFL x CTC25 - 010 under 'Tahiti' and 'Star Ruby' and TSKFL x CTC13 - 005 under 'Star Ruby'.
Although some genotypes have not been prominent among the ones with the highest estimated means, TSKC x CTSW - 064, LRF x (TR x LCR) - 005 and TSKFL x CTTR - 013 have the potential to be used in genetic breeding programs in order to obtain plants with tolerance to salinity, since by analysing averages of these genotypes in the highest and the lowest levels of salinity, the difference is less than 10% (Table 4). Compatibility between individuals (scion and rootstock) should be verified during the formation of citrus plants (scion-rootstock combination), especially because these are new materials. Thereby, according to the results obtained in this study there was no "elephant foot" phenomenon. However, as the materials are young, and the demand for scions is still small, definitive conclusions in this regard must be obtained from field studies.