Full Length Research Paper

ABSTRACT
Water stress is the most important problem in plant growth and development. Greenhouse trial is carried out on 2 durum wheat (Triticum durum Desf) genotypes (MBB and WAHA), to evaluate the effect of water stress on the biochemical content of leaves, and the possibility of reducing this effect by applying a growth regulator Indole-3-acetic acid (IAA) by seeds soaking and foliar spraying. Water stress causes a large accumulation of proline and soluble sugars and a decrease in the chlorophyll content of the leaves. The results obtained show that the effect of the hormone on the above-mentioned parameters is different according to the genotype and the mode of treatment, as well as the growth phase of the plant, without being able to promote application by soaking or spraying. The application of IAA has relatively reduced the effect of water stress by promoting the synthesis of proline and soluble sugars as osmotic regulators, and by increasing the chlorophyll content of the leaves. proline and soluble sugars concentrations showed negative and significant correlations with those of total chlorophyll. Our present study highlights some biochemical responses of plants to tolerate a water deficit and the possible involvement of exogenus application of IAA, as a phytohormone, in these regulatory mechanisms.
Key words: Triticum durum Desf, water stress, Indole-3-acetic acid (IAA), seed soaking, foliar spraying.
INTRODUCTION
Water is a vital element for the growth of plants. It is the most important factor that determines the growth and development of organisms. Plants are exposed to a variety of abiotic stresses in nature and exhibit unique and complex responses to these stresses depending on their degree of plasticity. When plants grow in a water-limited environment, they undergo morpho-physiological, phenological and biochemical modifications, to maintain constant cellular water potential, considered as water deficiency adaptation mechanisms (Manavalan et al., 2009; Basu et al., 2016). Plants accumulate osmolytes or compatible solutes to protect the cellular machinery from various environmental stresses (Giri, 2011). The most well-known osmolytes are glycine betaine (GB), sugars (mannitol, sorbitol, and trehalose), polyamines, and proline. These osmolytes get accumulated under various abiotic stresses and confer tolerance to cell without interfering with the cellular machinery of the plant (Chen, 2002; Anjum et al., 2017).The accumulation of proline and sugars is a clear marker for environmental stress, particularly in plants under drought stress (Watanabe et al., 2000). Proline and sugars accumulation may also be part of the stress signal influencing adaptive responses in drought stress conditions by helping to maintain membrane stability, preventing and protecting membrane fusion and; keeping protein so as to remain functional (Maggio et al., 2002; Xonostle-Cazares et al., 2010; Arabzadeh, 2012). Drought stress caused a large decline in chlorophyll a, chlorophyll b content, and the total chlorophyll content in all sunflower varieties investigated Ommen et al. (1999) and Manivannan et al. (2007) reported that leaf chlorophyll content decreases as a result of drought stress. Severe drought stress also inhibits the photosynthesis of plants by causing changes in chlorophyll content, affecting cholorophyll components and damaging the photosynthetic apparatus (IturbeOrmaetxe et al., 1998).
Phytohormones are the key regulators of plant growth and developmental processes and also crucial for biotic and abiotic stress response throughout their life cycle (Sah et al., 2016; Ullah et al., 2017). Phytohormones including ABA, jasmonic acid (JA), ethylene (ET), and salicylic acid (SA) are involved in osmotic adjustment and other drought-related processes (Khan et al., 2015; Vishwakarma et al., 2017). Auxins are the group of phytohormones which play a significant role in plant growth, development and response to various stresses (Singh et al., 2017). IAA is an auxin that participates in many plant processes including oxidative stress defense. It has become more evident that adaptation to drought is accompanied by an increase in the IAA levels (Zholkevich and Pustovoitova, 1993). Indole acetic acid (IAA) plays a vital role in maintaining plant growth under stress conditions (Gulnaz et al., 1999; Iqbal and Ashraf, 2007).
In the present study, we investigated the effects of exogenous indole − 3-acetic acid (IAA) on biochemical changes of durum wheat under drought stress conditions. Two genotypes were selected for this study: WAHA, which is considered a relatively drought-resistant genotype, and MBB, which is on the other hand sensitive to water deficit Subjected to different levels of water stress while counteracting this constraint by soaking the seeds in the IAA solution before transplanting, and by spraying the plant leaves with the IAA solution at three growth stages. To study the plant responses, some biochemical parameters (the ratio of proline, dissolved sugars, chlorophyll) are evaluated, which are supposed to contribute to plant adaptation under abiotic stress conditions and that can be adopted in the selection of plant varieties programs for this purpose.
MATERIALS AND METHODS
Trials management
The study was conducted in a plastic house located in the compound of the lead section - University of Skikda, Algeria; its estimated temperature was between 9°C and 15°C at night and between 24°C and 42°C during the day; its humidity ranged between 75 and 100%. The plastic house opens daily during hot days. 3 levels of watering were used:
S0 level = Field capacity watering.
S1 level = 50% field capacity Watering.
S2 level = 25% field capacity Watering.
The hormone is applied by seed soaking (7 ppm IAA solution) and leaf spraying (0.5ppm IAA solution). The experiment design was 2 factorials (3 water levels and 2 durum wheat genotypes (WAHA and MBB) and two types of IAA application laid in completely randomized design (CRD) with three replications. A total of 36 pots (28 cm deep with 25 cm diameter at top) were prepared with each 6 kg of Homogeneous agricultural soil which is relatively rich in organic matter.
Field capacity (100%) of the soils was determined through gravimetric methods before use.
Seeds of each cultivar were presoaked in hormonal solution (7ppm) in the dark for 48 h.
Proline accumulation in the fresh leaves was determined according to the method of Bates et al. (1973). Free proline was extracted using aqueous sulfosalicylic acid. The filtrate (1 ml) was mixed with equal volumes of glacial acetic acid and ninhydrin reagent (1.25 g ninhydrin, 30 ml of glacial acetic acid, 20 ml of H3PO4) and incubated for 1 h at 100°C. The reaction was stopped by placing the test tubes in cold water. The reaction mixtures were rigorously mixed with 3 ml toluene. The absorbance of toluene phase was estimated at 520 nm using a spectrophotometer. The proline concentration was determined using a standard curve.
Soluble sugars were determined based on the phenol-sulphuric acid method (Dubois et al., 1956). 0.1 g of dry leaves was homogenized with deionized water, filtered and the extract was treated with 1 ml of phenol (5%) and 5 ml of sulphuric acid (96%). The mixture was incubated at room temperature for 1h and then absorbance at 490 nm was read on a spectrophotometer. Contents of soluble sugar were determined by using glucose as a standard and expressed as µg/mg.
Chlorophyll pigments was extracted from 100 mg crushed fresh leaves in a sufficient volume of 80% acetone; chlorophylls a, b and total chlorophyll were estimated by following the method of Inskeep and Bloom (1985).
Statistical analysis
The data are presented as the mean values ± SE. Every treatment was replicated three times. Statistical analysis was performed using two-way ANOVA and differences between the mean values were compared using the LSD test at P ≤ 0.05.
RESULTS
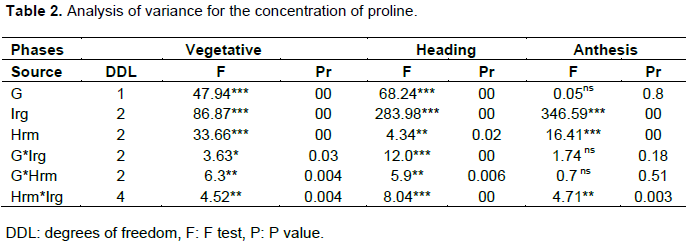
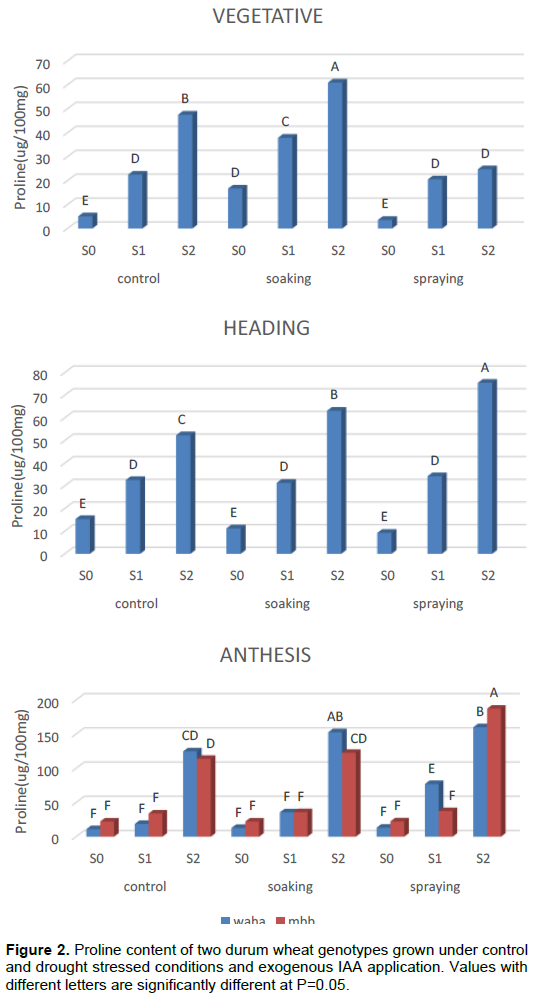
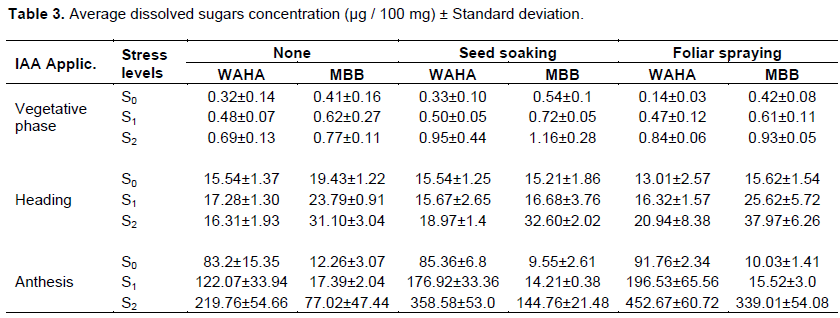
Table 1 and Figure 1 show the effect of the genotype, hormone treatment and the irrigation levels on the average leaf content of proline during plant development. Results showed significant variation in free proline content within a variety as well as among varieties under different treatments. Both varieties have the ability to accumulate proline under osmotic stress to different degrees. The amount of free proline was high under osmotic stress compared to unstressed conditions. Comparing the results of the three phases, the of proline increased by increasing the duration of water stress, Plants in the S2 irrigation level recorded the highest proline content at each stage while those in the S0 treatment had the lowest. Proline content of the two wheat genotypes increased significantly under drought treatment in all plant development stages. WAHA accumulates more proline than MBB during the vegetative phase. The effect of hormone treatment varied from one phase to another, where the seed soaking has a positive effect during the vegetative phase for WAHA. Spraying has negative effect on the two genotypes; whereas during the other two phases it is the spraying treatment that has a positive effect on the accumulation of proline in both genotypes.
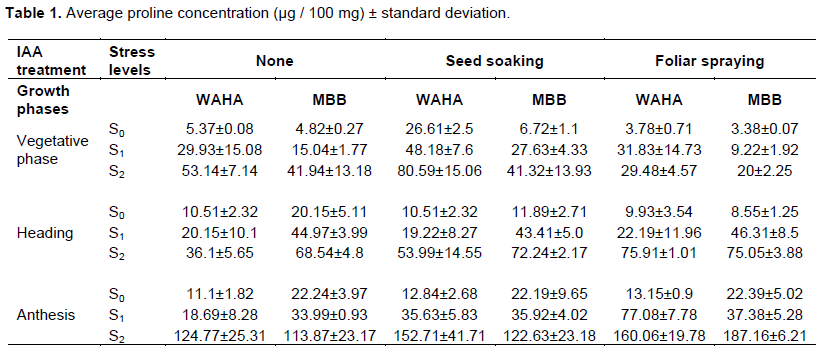

Analysis of variance (ANOVA) shows that the main effects, genotype, irrigation and hormone treatment are generally significant in all phases (Table 2). This indicates that the two genotypes have the capacity to modulate their endogenous proline to avoid water deficit conditions and that the application of the hormone significantly affects this response. First order interactions are also generally significant which indicates that the effect of each treatment differs from the levels of the others; thus the effect of the hormone on the accumulation of proline is dependent on the genotype and the intensity of the water stress. In fact, the accumulation of proline by each genotype is the result of the main effects and their interactions. The NEWMAN-KEULS test at the limit (α = 0.05%) shows the presence of homogeneous groups for the second order interaction between effects: genotype, irrigation and hormone treatment (Figure 2).
Sugars
Table 3 and Figure 3 show the effect of the genotype, hormone treatment and irrigation levels on the average leaf content of sugars during plant development. The accumulation of soluble sugars increased by increasing the duration of water stress. The two genotypes significantly increase their sugar content under stress conditions during anthesis, in particular. Note that genotype MBB accumulates more soluble sugar than WAHA during vegetative and heading phases. The opposite is noted in the anthesis. The effect of hormone treatment varied from one phase to another, where both seed soaking and foliar spraying by IAA have a positive effect on the accumulation of sugars for the two genotypes. During the heading phase this effect is less, whereas during the anthesis phase, spraying application has a strong positive effect on the two genotypes.

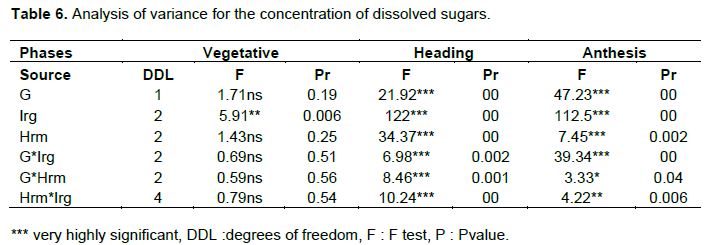
Analysis of variance (ANOVA) shows that the main effects, genotype, irrigation and hormone treatment as well as those first order interactions are generally significant (Table 4; Figure 4) suggesting that the two genotypes accumulate soluble sugars at different levels in response to water stress and that IAA is involved in this response. Interactions are not significant at the vegetative stage. This indicates that the soluble sugars synthesis is dependent only on main effects.
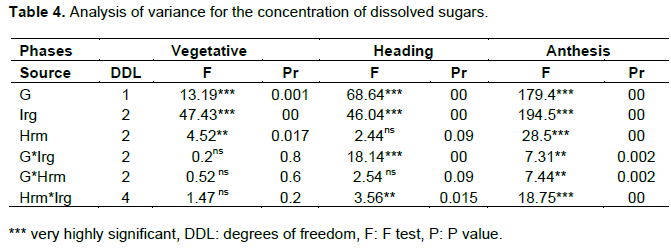
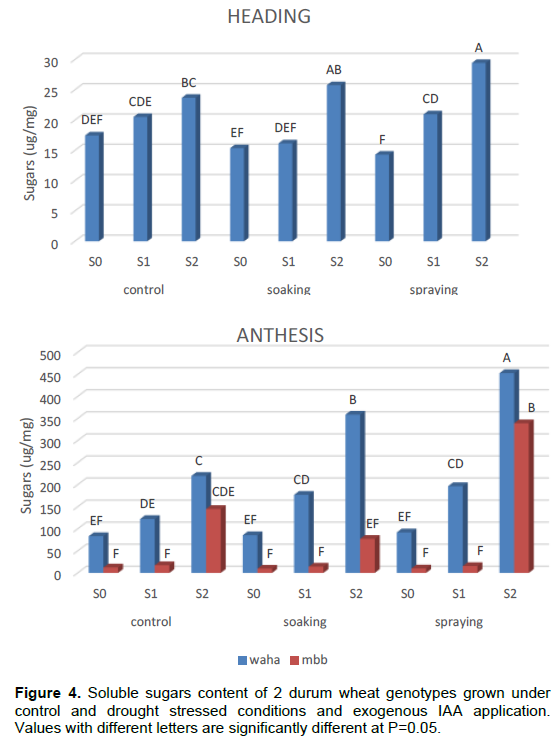
The NEWMAN-KEULS test at the limit (α = 0.05%) shows the presence of homogeneous groups with the smallest significant difference (LSD) for the first and second order interaction between effects: genotype, irrigation and hormone treatment (Figure 4).
Chlorophyll
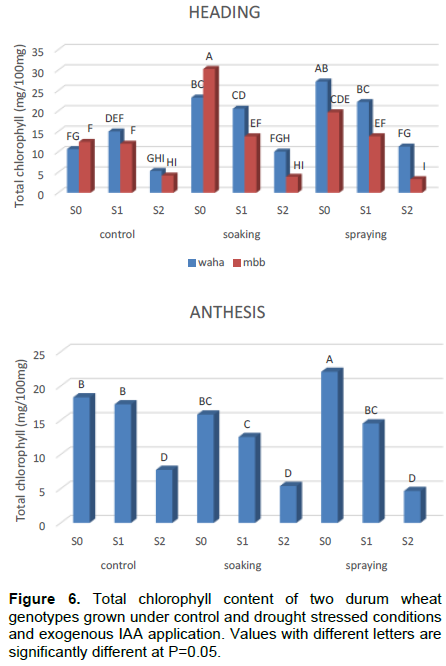
The results of the 3 stages of plant development (Table 5; Figure 5) show that the total chlorophyll content decreases significantly under stress conditions. Both hormonal treatments significantly improve the chlorophyll content of WAHA during the heading phase; and spraying application was shown to be the best.

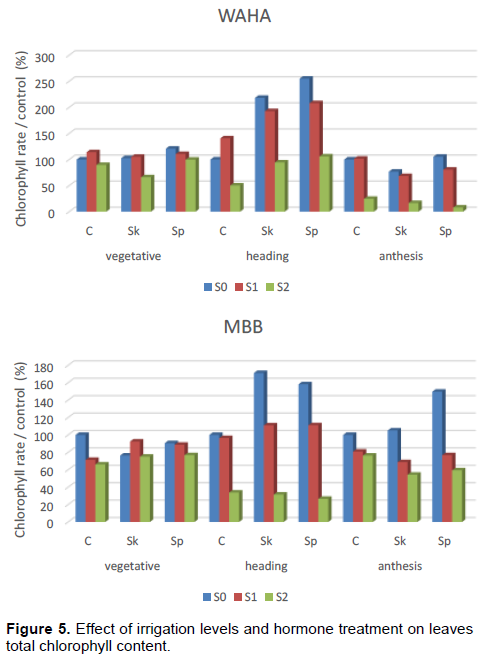
ANOVA shows that the main effects, genotype, irrigation and IAA treatment as well as their first order interactions are significant at the heading and the anthesis phases (Table 6). This indicates that the chlorophyll synthesis is affected by water stress and IAA treatment. The NEWMAN-KEULS test at the limit (α =0.05%) shows the presence of homogeneous groups with the smallest significant difference (LSD) for the second order interaction between effects: genotype, irrigation and hormone treatment (Figure 6).
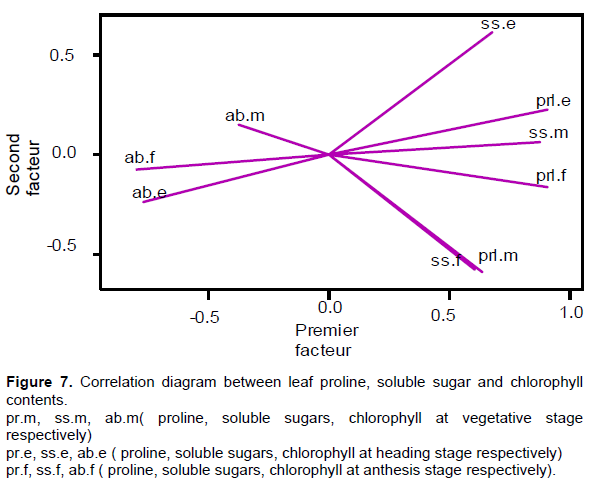
To examine the relationships between the different measurements made, we performed a PCA (Figure 7). The percentage of information for axis 1 was 56.5% and 13.5% for axis 2, that is, 70%. Prol.e / prl.f, prl.e / ss.m and prl.f / ss.f where the correlation coefficient was (0.80), 0.78) and (0.76) respectively, noting that the ss.e / prl.m and ss.e / ss.f correlations were weak. Correlations between chlorophyll ratios were low except for ab.e / ab.f which were significant (0.54). The correlations between proline and dissolved sugar concentrations and total chlorophyll concentrations were negative and significant, the most important being: (-0.75), (-0.73) and (-0.70) for ab.f / prl. e, ab.f / ss.mÙˆ ab / prl.e, respectively.
DISCUSSION
The concentration of proline and soluble sugars in leaves of the two wheat genotypes significantly increased at the different growth stages in stressed plants. The highest levels of accumulation were in the anthesis phase. These results suggest that the osmotic adjustment is a common response of plants under drought conditions irrespective of the growth stage of the plant. Proline, a compatible osmolyte, is known to accumulate under abiotic stress (Sandhya et al., 2010). Osmotic adjustment through the accumulation of cellular solutes, such as proline, has been suggested as one of the possible means for overcoming osmotic stress caused by water loss (Caballero et al., 2005). According to Umezawa et al. (2006), plants have the ability to accumulate non-toxic compounds such as proline which protects cell damage due to low water potential of cells, which is a way of plant adaptation to drought stress tolerance. The accumulation of glucose in different wheat varieties was observed under the conditions of water deficit (El-jaafari, 1993; Brinis, 1995). Several studies have also shown a positive correlation between the accumulation of proline and sugars and the severity of water stress (Berllinger et al., 1991; Gorham, 1993).
The results show that the IAA treatment mostly led to a greater accumulation of both proline and sugars in the two varieties in the anthesis and heading stages in particular, suggesting that IAA had a positive effect in improving drought tolerance of wheat. Studies have shown that plants pre-treated with IAA exhibited enhanced drought resistance (Almazroue, 2014). Foliar spraying with growth regulators (IAA and GA3 ) showed significant effect on plant, to the extent of reducing the hurt effect of salinity on the vegetative measurements and some physiological components of plant (Gherroucha et al., 2011). Shalaby and Kishk (1986) confirmed the increase of proline accumulation in the presence of growth regulators; stimulating hormones, especially quinine, regulate the synthesis and accumulation of dissolved sugars in growing plants in saline media. Chlorophyll content decreased significantly under water stress in both wheat genotypes; various reports have noted that the water deficit decreases considerably, depending on the intensity and duration of the stress, the total chlorophyll content in different plant species (Mafakheri et al., 2010; Gholamin and Khatnezhad, 2011; Din et al., 2011; Mouradi et al., 2016). Exogenous IAA appears to have significant beneficial effects on chlorophyll content; Zhang et al. (2020), in a study on white clover, noted that the chlorophyll content of leaves increased significantly when treated with exogenous IAA. Negative and significant correlations between proline and dissolved sugar concentrations and total chlorophyll concentrations were found. These results are in accordance with other researches (Schonfeld et al., 1988; Bayoumi et al., 2008; Rad et al., 2012).
CONCLUSION
From the present experiment, it can be concluded that water deficit significantly increases the leaf content in proline and soluble sugars and decreases their chlorophyll content in both genotypes used in this study. The IAA application considerably counteracts these effects by improving the biosynthesis of these components. The effect of hormone treatment varies from one phase to another where the leaf spraying treatment was shown to be the best.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Almazroue HA (2014). Identification, cloning, and expression of tobacco responsive to dehydration like protein (RD22), SBIP-355 and its role in SABP2 mediated SA pathway in plant defense [D]. |
|
|
Anjum SA, Ashraf U, Tanveer M, Khan I, Hussain S, Shahzad B, Zohaib A, Abbas F, Saleem MF, Ali I, Wang LC (2017). Drought Induced Changes in Growth, Osmolyte Accumulation and Antioxidant Metabolism of Three Maize Hybrids. Front. Plant Science 8:69. |
|
|
Arabzadeh N (2012). The effect of drought stress on soluble carbohydrates (Sugars) in two species of Haloxylonpersicum and Haloxylonaphyllum. Asian Journal of Plant Science 11(1):44-51. |
|
|
Basu S, Ramegowda V, Kumar A, Pereira A (2016). Plant adaptation to drought stress. F1000Res 5:F1000 faculty Rev-1554. |
|
|
Bates LS, Waldren RP, Teare ID(1973). Rapid determination of free proline for water-stress studies. Plant Soil 39:205-207. |
|
|
Bayoumi TY, Eid MH, Metwali EM (2008). Application of physiological and biochemical indices as a screening technique for drought tolerance in wheat genotypes. African Journal of Biotechnology 7:2341-2352. |
|
|
Berllinger Y, Bensaoud A, Larher F (1991). Physiology significance of proline accumulation , a J.P. Eds Physiology breeding of winter cereals for stressed Mediterranean environment, Montpelier (France), July 3-6 1989. 55:449-458. |
|
|
Brinis L (1995). Effet du stress hydrique sur quelques mécanismes morphophysiologiques et biochimiques de traits d'adaptation et déterminisme génétique chez le blé dur (Triticum durum Desf). Thèse de Doctorat. Univ. Annaba. |
|
|
Caballero JI, Verduzco CV, Galan J (2005). Proline accumulation as a symptom of drought stress in maize: A tissue differentiation requirement. Journal of Experimental Botany 39:889-897. |
|
|
Chen TH, Murata N (2002). Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Current Opinion in Plant Biology 5:250-257. |
|
|
Din J, Khan S, Ali I, Gurmani A (2011).Physiological and agronomic response of canola varieties to drought stress. Journal of Animal and Plant Sciences 21:78-82. |
|
|
Dubois M, Gilles KA, Hamilton JK, Rebers P T, Smith F (1956). Colorimetric method for determination of sugar and related substances. Analytical Chemistry 28(3):350-356. |
|
|
El-Jaafari S (1993). Contribition à l'étude des mécanismes biophysiques et biochimiques de la resistance à la sècheresse chez le blé. Thèse de doctorat d'état pp. 3-34. |
|
|
Gherroucha H , Fercha A, Ben Mekhlof A (2011). Foliar application of Indol Acitic Acid (IAA) and Gebirilic acid (GA3) as well as interaction effect on growth yield and some physiological compositions of Triticum plant grown under salinity conditions. Agriculture and Biology Journal of North America 2:512-521. |
|
|
Gholamin R, Khayatnezhad M (2011). The effect of end season drought stress on the chlorophyll content, chlorophyll fluorescence parameters and yield in maize cultivars. Scientific Research and Essays 6:5351-5357. |
|
|
Giri J (2011). Glycinebetaine and abiotic stress tolerance in plants. Plant signaling and behavior 6:1746-1751. |
|
|
Gorham J (1993). Stress tolerance and mechanisms behind tolerance in barley. In: Agronomical and physiological characterization of different barley genotypes to salt stress. Settat 1993. Meeting. |
|
|
Gulnaz A, Iqbal J, Farooq S, Azam F (1999). Seed treatment with growth regulators and crop productivity. I. 2,4-D as an inducer of salinity-tolerance in wheat (Triticum aestivum L.). Plant Soil 210:209-217. |
|
|
Inskeep WP, Bloom PB (1985). Extinction coefficients of chlorophyll a and chlorophyll b in N, N- dimethyl formamide and 80% acetone. Plant physiology 77(2):483-485. |
|
|
Iqbal M, Ashraf M (2007). Seed treatment with auxins modulates growth and ion partitioning in salt-stressed wheat plants. Journal of Integrative Plant Biology 49(7):1003-1015. |
|
|
IturbeOrmaetxe I, Escuredo PR, Arrese-Igor C, Becana M (1998). Oxidative damage in pea plants exposed to water deficit or paraquat. Plant Physiol. 116:173-181. |
|
|
Khan MIR, Fatma M, Pe TS, Anjum NA, Khan NA (2015). Salicylic acid-induced abiotic stress tolerance and underlying mechanisms inplants. Frontiers in Plant Science 6:462 |
|
|
Mafakheri AB, Siosemardeh PC, Bahramnejad Y, Struik T, Sohrabi S (2010). Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Australian Journal of Crop Science 4: 580-585. |
|
|
Maggio A, Miyazaki S, Veronese P, Fujita T, Ibeas JI, Damsz B, Narasimhan ML, Hasegawa PM, Joly RJ, Bressan RA (2002). Does proline accumulation play an active role in stress-induced growth reduction. Plant Journal 31:699-712. |
|
|
Manavalan LP, Guttikonda SK, Phan Tran LS, Nguyen HT (2009). Physiological and molecular approaches to improve drought resis-tance in soybean. Plant and Cell Physiology 50:1260-1276 |
|
|
Manivannan P, Abdul Jaleel C, Sankar B, Kishorekumar A, Somasundaram R, Lakshmanan GMA, Panneerselvam R (2007). Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids and Surfaces B: Biointerfaces 59:141-149. |
|
|
Mouradi M, Farissi M, Bouizgaren A, Makoudi B, Kabbadj A, Very AA, Sentenac H, Qaddourya A, Ghoulam C (2016). Effects of water deficit on growth, nodulation and physiological and biochemical processes in Medicago sativa-rhizobia symbiotic association. Arid Land Research and Management 30. |
|
|
Ommen OE, Donnelly A, Vanhoutvin S, Van Oijen M, Manderscheid R (1999). Chlorophyll content of spring wheat flag leaves grown under elevated CO2 concentrations and other environmental stresses within the ESPACE-wheat project European Journal of Agronomy 10:197-203. |
|
|
Rad RN, Kadir MA, Jaafar HZ, Gement DC (2012). Physiological and biochemical relationship under drought stress in wheat (Triticum aestivum). African Journal of Biotechnology 11(24):1574-1578. |
|
|
Sah SK, Reddy KR, Li J (2016). Abscisic acid and abiotic stress tolerancein crop plants. Frontiers in Plant Science 7:571. |
|
|
Sandhya V, Ali SZ, Venkateswarlu B, Reddy G, Grover M (2010). Effect of osmotic stress on plant growth promoting Pseudomonas spp. Archives of Microbiology 192:867-876. |
|
|
Schonfeld MA, Johnson RC, Carver BF, Mornhinweg BW (1988). Water relation in winter wheat as drought resistance indicators. Crop Science 33:287-294. |
|
|
Shalaby A, Kishk E (1985). A comparative study between foliar and seed-soaking applications of B9 (N-Dimethyl Amino Succinamic Acid) for increasing salt tolerance of Sudan grass plants. The Desert Institute Bulletin (Egypt). |
|
|
Singh VP, Prasad SM, Munné-Bosch S, Müller M (2017). Phytohormones and the regulation of stress tolerance in plants: cur-rent status and future directions. Frontiers in Plant Science 8:1871 |
|
|
Ullah A, Sun H, Yang X, Zhang X (2017). Drought coping strategies incotton: increased crop per drop. Plant Biotechnology Journal 15:271-284. |
|
|
Umezawa T, Fujita M, Fujita Y, Yamaguchi-Shinozaki K, Shinozaki K (2006). Engineering drought tolerance in plants: discovering and tailoring genes to unlock the future. Current Opinion in Biotechnology 17:113-122. |
|
|
Vishwakarma K, Upadhyay N, Kumar N, Yadav G, Singh J, Mishra RK, Kumar V, Verma R, Upadhyay RG, Pandey M, Sharma S (2017). Abscisicacidsig-nalling and abiotic stress tolerance in plants: a review on currentknowledge and future prospects. Frontiers in Plant Science 8:161. |
|
|
Watanabe S, Kojima K, Ide Y, Sasaki S (2000). Effects of saline and osmotic stress on proline and sugar accumulation in Populus euphratica in vitro. Plant Cell, Tissue and Organ Culture 63(3):199-206 |
|
|
Xoconostle-Cazares B, Ramirez-Ortega FA, Flores-Elenes L, Ruiz-Medrano R (2010). Drought tolerance in crop plants. American Journal of Plant Physiology 5:241-256. |
|
|
Zhang Y, Li Y, Hassan MJ, Li Z, Peng Y (2020). Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC plant biology 20:1-12. |
|
|
Zholkevich VN, Pustovoitova TN (1993). Growth and Phytohormone Content in Cucumis sativus L. Leaves under Water Deficiency, Fiziol. Rast (Moscow), 40:676-680 (Russian Journal of Plant Physiology the English-language translation). |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


