Full Length Research Paper

ABSTRACT
The cactus pear is a widely cultivated plant in the Northeast of Brazil, contributing significantly to the feeding of livestock, especially in times of drought. Because of the high variation between phenotypic varieties grown in this region, it is essential to study the genetic diversity. The objectives of this study were to characterize the genetic diversity in seven varieties of cactus pear, genera Opuntia and Nopalea, through 19 morphological and behavioral characteristics, and to determine the phenotypic correlation and importance of these characteristics to the variability among genotypes, using multivariate analysis techniques. The study was conducted at the experimental station of Agronomic Institute of Pernambuco (IPA), located in city of Arcoverde, State of Pernambuco, Brazil using randomized block design with three replications. The materials IPA-100003, IPA-200016, IPA-200008, IPA-100004, IPA-200021, IPA-200205 and IPA-200149 were evaluated for 19 quantitative characteristics of the plants. The collected data were analyzed by analysis of variance by F test, and the means grouped by the Scott-Knott test (p<0.05). The broad-sense heritability and phenotypic correlation characteristics were estimated. The genetic diversity was estimated by multivariate methods (unweighted pair group method with arithmetic mean-UPGMA, Tocher, principal component and canonical variables). Analyses of variance and genetic diversity revealed significant differences among genotypes, with the possible formation of two, three or four genetically distinct groups. The heritability values ranged from 79.6 to 97.0% for all 19 quantitative characteristics. The water content and cladode fresh matter are the characteristics that contributed most to the genetic divergence among the materials. Moreover, these characteristics are significantly and positively correlated with dry matter, width, length and cladode area. Thus, the genetic variability among the studied varieties of cactus pear and their potential use in breeding programs are confirmed. The uni and multivariate methods used for the genetic divergence differ and gather genotypes in two, three or four groups.
Key words: Brazilian semiarid, forage, genetic distance, grouping, multivariate analysis.
INTRODUCTION
MATERIALS AND METHODS
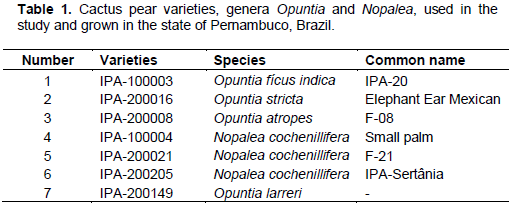
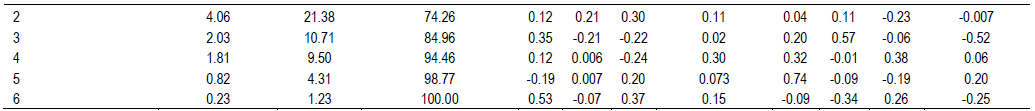
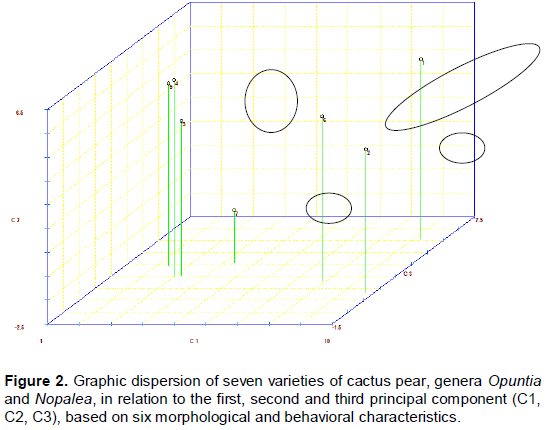
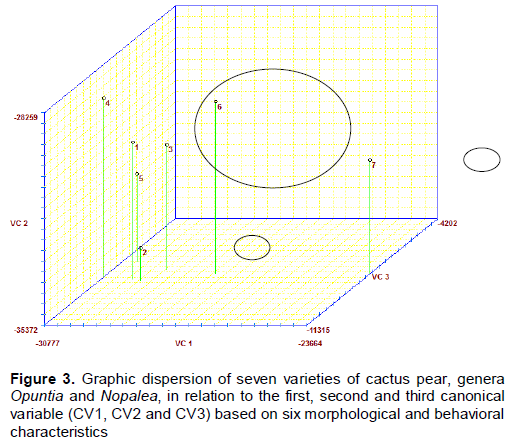
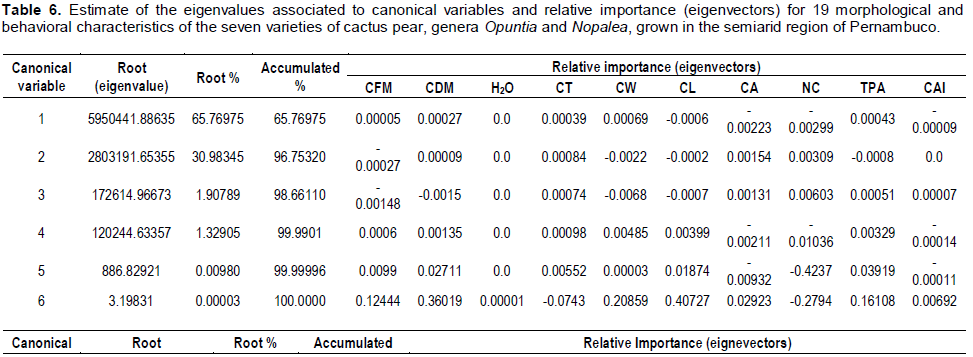
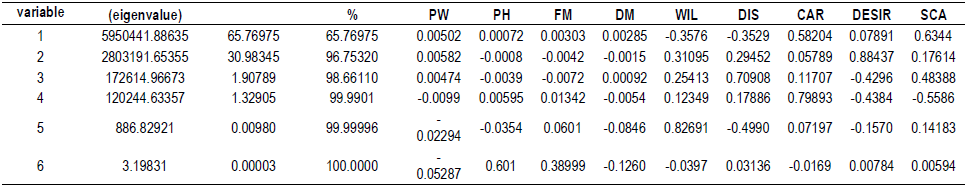
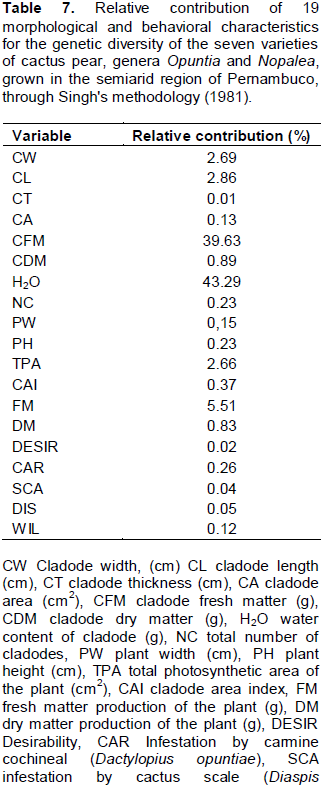
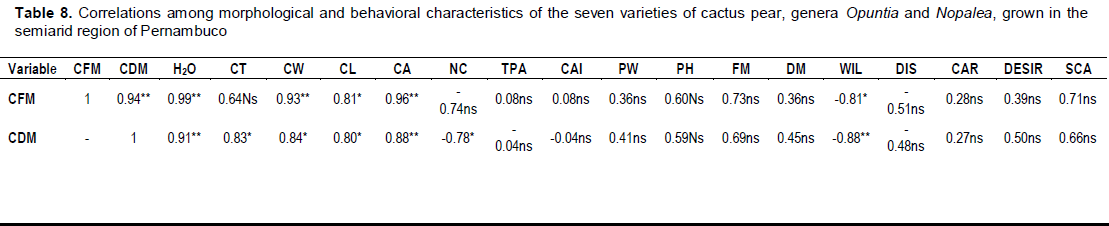
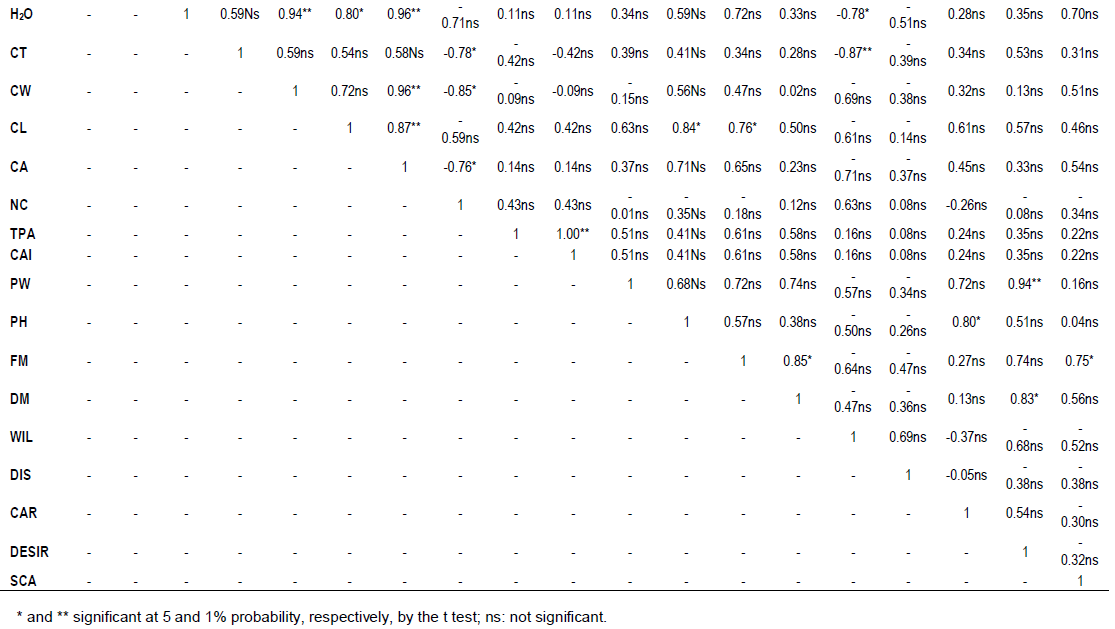
RESULTS AND DISCUSSION
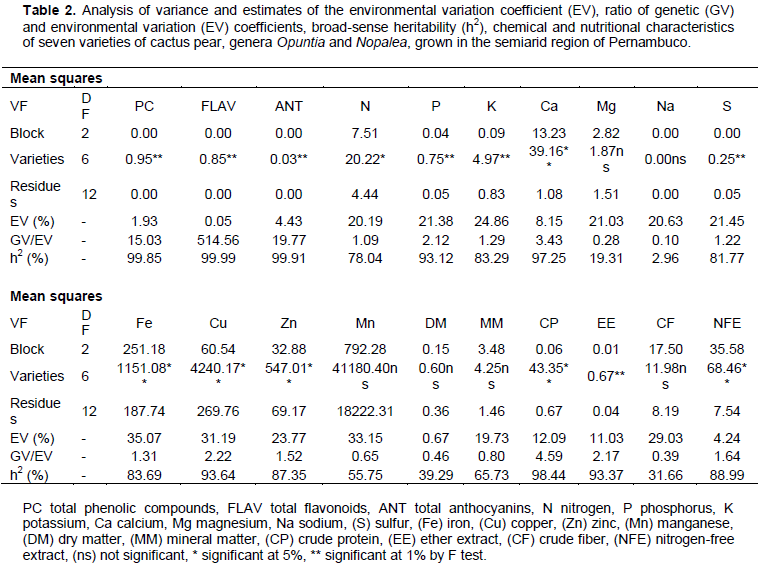
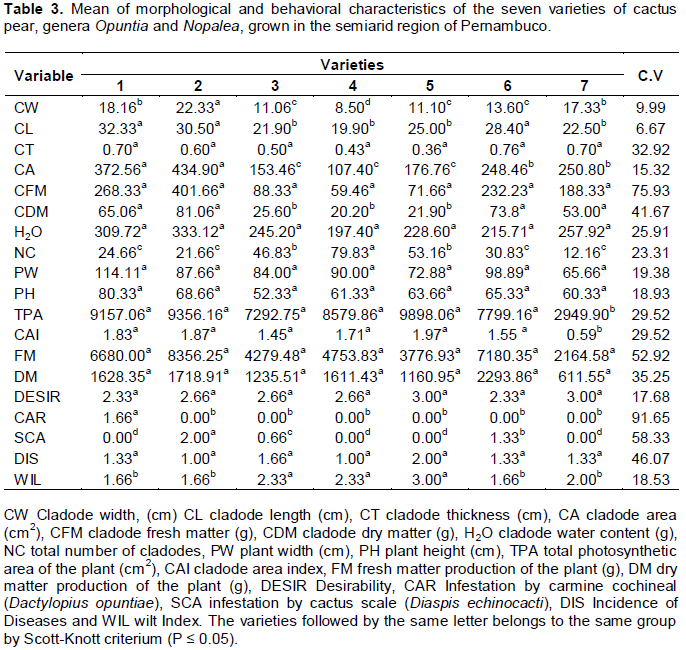


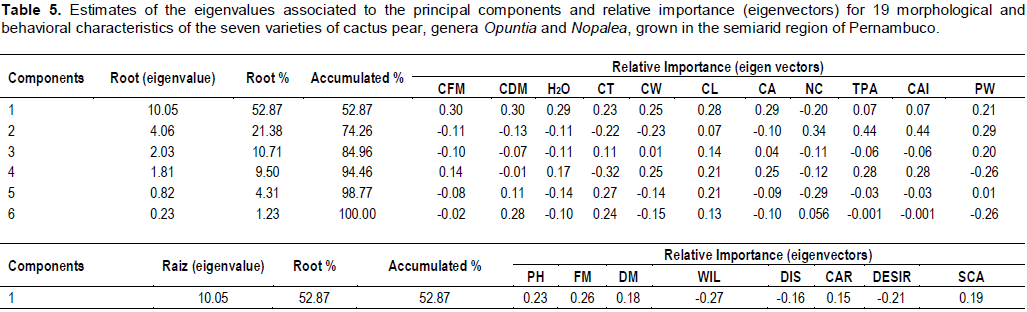

CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Alves FAL, Santos DC dos, Pereira VLA, Silva M da C, França LTV, Amorim PMD de (2013). Estimativa de produção de massa fresca, seca e acúmulo de água em palma forrageira dos gêneros Opuntia e Nopalea. In: Congresso Nordestino de Produção Animal, 8, Fortaleza, Brasil pp. 1-4. |
|
|
Alves JCS, Peixoto JR, Vieira JV, Boiteux LS (2006). Herdabilidade e correlações genotípicas entre caracteres de folhagem e sistema radicular em famílias de cenoura, cultivar Brasília. Hortic. Bras. 24:363-367. |
|
|
Amorim EP, Souza JC (2005). Híbridos de milho inter e intrapopulacionais obtidos a partir de populações S0 de híbridos simples. Bragantia 64:561-567. |
|
|
Amorim PL de (2011). Caracterização morfológica e produtiva em variedades de palma forrageira. (Dissertação de Mestrado). Universidade Federal de Alagoas, Rio Largo. 54 p. |
|
|
Araújo LF, Oliveira LSC, Perazzo-Neto A, Alsina OLS, Silva FLH (2005). Equilíbrio higroscópico da palma forrageira: Relação com a umidade ótima para fermentação sólida. Bras. Eng. Agríc. Ambient. 9:379-384. |
|
|
Arriel NHC, Di-Mauro AO, Di-Mauro SMZ, Bakke AO, Unêda-Trevisoli SH, Costa MM, Capeloto A, Corrado AR (2006). Técnicas multivariadas na determinação da diversidade genética em gergelim usando marcadores RAPD. Pesqui. Agropecuária Bras. 41:801-809. |
|
|
Barbera G (2001). História e importância econômica e agroecológica. In: G Barbera (Ed.), Agroecologia, cultivo e usos da palma forrageira. Roma: Food Agric. Organ. United Nations/Braz. Support Serv. Micro Small Entreprises of Paraiba. pp. 1-11. |
|
|
Barrios EP, Mu-oz-Urias A (2001). Domesticação das Opuntias e variedades cultivadas. In: G Barbera (Ed.), Agroecologia, cultivo e usos da palma forrageira. Roma: Food Agric. Organ. United Nations/Braz. Support Serv. Micro Small Entreprises of Paraiba. pp. 58-64. |
|
|
Cruz CD (2001). Programa GENES: Aplicativo computacional em genética e estatística. Viçosa: UFV 648 p. |
|
|
Cruz CD, Regazzi AJ, Carneiro PCS (2012). Modelos biométricos aplicados ao melhoramento genético. (4. ed.). Viçosa: UFV. 514 p. |
|
|
Falconer DS (1987). Introdução à genética quantitativa. (1. ed.). Viçosa: UFV 279 p. |
|
|
Ferreira CA, Ferreira RLC, Santos DC dos, Santos MVF dos, Silva JAA da, Lira M de A, Molica SG (2003). Utilização de Técnicas Multivariadas na Avaliação da Divergência Genética entre Clones de Palma Forrageira (Opuntia ficus-indica Mill.). Rev. Bras. Zoot. 32:1560-1568. |
|
|
Gonçalves PS, Martins ALM, Bortolleto N, Tanzizi MR (1996). Estimates of genetic parameters and correlations of juvenile characters based on open-pollinated progenies of Hevea. Braz. J. Genet. 19:105-111. |
|
|
Guimarães CM, Stone LF (2008). Métodos de avaliação das condições hídricas das plantas. Comunicado Técnico 161, Goiás, Embrapa Arroz e Feijão 8 p. |
|
|
Inmet - Instituto Nacional de Meteorologia (2015). Dados climáticos da Estação de Arcoverde: série histórica de 1961 a 2014. Banco de dados do Instituto Nacional de Meteorologia. Disponível em. View |
|
|
López-García JJ, Fuentes-Rodríguez JM, Rodríguez RA (2001). Production and use of Opuntia as forage in Northern Mexico. In: C Mondragón-Jacobo (Ed.), Cactus (Opuntia spp.) as forage. Roma: Food Agric. Organ. United Nations. pp. 29-36. |
|
|
Neder DG, Costa FR da, Edvan RL, Souto-Filho LT (2013). Correlations and path analysis of morphological and yield traits of cactus pear accessions. Crop Breed. Appl. Biotechnol. 13:203-207. |
|
|
Paixão SL (2012). Avaliação morfológica de clones e progênies de palma forrageira. (Tese de Doutorado). Universidade Federal Rural de Pernambuco, Recife. 70p. |
|
|
Pereira VLA, Santos DC dos, Silva M da C, Alves FAL, Soares G, Costa AF da, Mesquita FLT de (2014). Selection of genotypes of cactus pear adapted to semiarid conditions of Pernambuco state. In: Reunião Anual da Sociedade Brasileira de Zootecnia, 51, Barra dos Coqueiros, Brasil. 1 p. |
|
|
Rao CR (1952). Advanced statistical methods in biometric research. New York: John Wiley & Sons. 390 p. |
|
|
Rêgo ER do, Rêgo MM do, Cruz CD, Finger FL, Casali VWD (2011). Phenotypic diversity, correlation and importance of variables for fruit quality and yield traits in Brazilian peppers (Capsicum baccatum). Gen. Resour. Crop Evol. 58:909-918. |
|
|
Rufino ER, Siqueira WJ, Marques MOM, Colombo CA, Chiorato AF, Azevedo-Filho JA, Lourenção AL, Yamamoto PY, Martins ALM (2010). Estimativas de parâmetros genéticos de caracteres relacionados ao vigor de estacas em Lippia alba cuttings. Bragantia 69:779-785. |
|
|
Sales AT, Andrade AP, Silva DS, Leite MLMV, Viana BL, Ramos JPF (2003). Taxa de crescimento relativo de palma forrageira (Nopalea cochenillifera Salm-Dyck). Pesqui. Agropecuária Bras. 34:340-346. |
|
|
Sales AT, Leite MLMV, Alves AQ, Ramos JPF, Nascimento JP (2013). Crescimento vegetativo de palma forrageira em diferentes densidades de plantio no Curimatú Paraibano. Technol. Ciênc. Agropecuária 7:19-24. |
|
|
Santos DC, Farias I, Lira MA, Santos MVF, Arruda GP, Coelho RSB, Dias FM, Melo JN (2006). Manejo e utilização da palma forrageira (Opuntia e Nopalea) em Pernambuco. Recife: IPA. 48p. |
|
|
Scott AJ, Knott M (1974). A cluster analysis method for grouping means in the analysis of variance. Biometrics 30:507-512. |
|
|
Silva FASE, Azevedo CAV (2006). A New Version of the Assistat-Statistical Assistance software. In: World Congress on Computers in Agriculture, 4, Orlando-FL-USA, Orlando: Am. Soc. Agric. Biol. Eng. pp. 393-396. |
|
|
Silva JAG da, Schwertner DV, Carbonera R, Kruguer CAMB, Crestani M, Gaviraghi F, Schiavo J, Arenhardt EG (2011). Distância genética em genótipos de girassol. Rev. Bras. Agrociênc. 17:326-337. |
|
|
Singh D (1981). The relative importance of characters affecting genetic divergence. Indian J. Gen. Plant Breed. 41:237-245. |
|
|
Viana JPG (2013). Diversidade genética em alho (Allium sativum L.). (Dissertação de Mestrado). Universidade Federal do Piauí, Teresina 56 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


