Full Length Research Paper

ABSTRACT
The present investigation studied the effectiveness of ozone (O3) concentrations on epidermal cells of pepper (Capsicum frutescens L.) leaves and its response to ascorbic acid (AS). The plants were grown at two sites in Riyadh, King Saud University (KSU) Campus, and the second industrial city (SIC) under an average of 42.33 and 138.66 ppb of O3. Two groups grown at KSU site as a control; one of them was treated with tap water (TW) and the other was treated with TW+AS, while the remaining two groups were transferred to SIC site, treated as described previously. Treatment with 300 mg/L AS was performed once every 15 days until the end of the experiment. The plants grown under separately high concentrations of O3 and AS increased the stomatal numbers, dimensions and cell dimensions in both upper and lower epidermises of leaves in comparison to control plant leaves. Treatment with O3+AS significantly increased the length of the upper and lower epidermal cells, while it decreased the cell widths in comparison to plants grown under only O3. The AS might have a mitigating effect on the impacts of O3 on leaf epidermal cells of the pepper plant particularly, with respect to cell width.
Key words: Ozone, epidermal cell traits, pepper, ascorbic acid.
INTRODUCTION
Pepper (Capsicum frutescens L.) is an annual herb or shrub, and belongs to the Solanaceae family. It is one of the most important vegetables grown in parts of the humid and semi-arid tropics (Aliyu, 2000 ). The fruits are extensively used as a cooking condiment (Alabi, 2006). Pepper contains an excellent source of vitamins A and C as well as phenolic compounds, which are important antioxidant (Shotorbani et al., 2013). Pepper is also used for the prevention and treatment of cold and fever (Udoh et al., 2005), as it contains vitamin C (Osuna-García et al., 1998). In addition, capsaicin has been shown to have great potential as a chemotherapeutic agent against several cancers (Oyagbemi et al., 2010; Clark and Lee, 2016). The leaf surface of the plant is the major part that receives, absorbs, and accumulates air pollutants (Chauhan and Joshi, 2010). The gaseous pollutants enter the leaf through the stomata, which have the potential to alter the metabolic processes of the plant and react with the intercellular water to form reactive oxygen species (ROS) that act on the plasma membrane and cause oxidative stress in the mesophyll cells of the leaf (Bray, 2000; Roshchina and Roshchina, 2013; Iriti and Faoro, 2008), cellular damage in the leaves, reduce photo-synthesis, decrease carbon allocation to sink tissues, and affect plant biomass and radial growth (Wittig et al., 2009).
O3 is one of the gaseous pollutants that have impact on many aspects of the plants, such as the morphological, physiological, and anatomical characteristics. These effects vary with the intensity and duration of O3 exposure (Pasqualini, 2003). Exposure of alfalfa to high ozone concentrations (85-120 ppb) causes obvious effects on cell organelles such as chloroplasts, plastoglobules, nuclei, vacuoles and chromatin in leaf mesophyll tissue and stem cortex, and no clear effects of ozone were noted on starch grain shapes and the mitochondria in the leaf mesophyll and stem cortex cells (El-Zaidy et al., 2019). The stomatal density in the leaf epidermis of different plants was affected by elevated O3 concentration (Evans et al., 1996; Paakkonen et al., 1998; Frey et al., 1996; Lawson et al., 2002; Zouzoulas et al., 2009; Gostin, 2009; Wahid and Ahmad, 2003; Pedroso and Alves, 2008), and caused sluggishness stomata efficiency, and gradual loss in stomatal control over transpiration (Feng et al., 2018). However, few works indicated lack of significant impact on the stomatal density of certain plants when exposed to O3 stress (Giacomo et al., 2010; Riikonen et al., 2010; Dumont et al., 2014). Elevated O3 concentration was reported to affect the epidermal cell density of the plant leaves (Lawson et al., 2002; Wahid and Ahmad, 2003; Pedroso and Alves, 2008; Zouzoulas et al., 2009; Riikonen et al., 2010). Moreover, Riikonen et al. (2010)reported that high concentration of O3 increased the epidermal cell size with no obvious effect on cuticular striations and epicuticular wax crystallites. Meanwhile, ascorbic acid (AS) is a growth regulator that plays important roles in many physiological processes (Ejaz et al., 2012; Kim et al., 2008; Hathout, 1995; Mukherjee and Choudhuri, 1985). González-Reyes et al. (1998) reported the effective role of AS in stress resistance, whereas Veljovic-Jovanovic et al. (2001) found that AS concentration is low in the O3-sensitive plant tissues, which confirms its role in oxidative stress. AS increased the thickness of both the midrib and lamina of leaf blades, the size of the main vascular bundle of the midrib, and also increased the average diameter of the vessel in the leaves of tomato plants, Zea mays, and Mentha arvensis (Ali, 2001; Ali et al., 2015; Hendi and Boghdady, 2016). Leaf seedlings of pre-soaked seeds with AS increased the stomatal length and decreased the epidermis cell length on both the surfaces. Although AS application reduced the epidermis cell number on the upper surface, it had no effect on this feature on the lower surface (Cavusoglu and Bilir, 2015). Treatment with AS increased the mitotic divisions and cellular dimensions in the cell elongation region of the plant root (Kaviani, 2014). Despite the vast amount of data on the effects of O3 and AS on the physiological, biochemical, and molecular characteristics of plants; the effects of O3 and AS on the epidermal cell traits of C. frutescens L. have not been studied in detail. Because the epidermis is the protective layer and acts as a barrier between the outer environment and the internal structures of the plant body, this research was an initiative to study the impact of high concentration of O3 on the leaf epidermal cell traits, and its response to treatment by AS, which might assist in better understanding of the phenomena occurring in the leaves.
MATERIALS AND METHODS
Two sites were selected for this research in Riyadh city with different pollution levels; the first site was King Saud University campus for the control group (Cont.), and the second site was the second industrial city (SIC). Pepper seeds were obtained from a local market in Riyadh, Saudi Arabia. Seeds were sterilized with 1% sodium hypochlorite for 7 min, and then rinsed with sterilized double distilled water. Seeds were planted under natural environmental conditions in used plastic pots containing sterile sandy and alluvial soil (ratio 1:1). A fungicide was added to prevent the fungal growth; plants were left to grow until the generation of initial leaves, and then transferred to the study sites. The plants were divided into four groups: two control groups that were left to grow in King Saud University site; one of them was treated with tap water, and the other was treated with tap water and AS; while the remaining two groups were transferred to SIC site upon exposure with O3, where one of them was treated with tap water, and the other was treated with tap water and AS. Irrigation was performed once every 15 days using 300 mg/L AS until the end of the experiment. The leaf surfaces were cleaned with distilled water, followed by silicon rubber imprinting for studying the epidermal characteristics according to Lloyd (1908). The slides were then examined and photographed using Zeiss Photomicroscope III. Epidermal cell dimensions and the stomatal number and dimensions were captured at 40X. Twenty-five measurements were recorded for each parameter and the number of stomata was counted in a microscopic field area of 0.25 mm2. All measurements and descriptions were recorded at the vegetative growth end before flowering (after 90 days of sowing). Measurement of O3 concentrations was performed daily for three months at each of the study sites using a measuring device (AEROQUAL Series 200 with Monitor), average readings of ozone (O3) levels in control plants site (KSU Campus) was 42.33 ppm, while in polluted plant site second site (SIC) was 138.66 ppm. The data obtained were statistically analyzed using SAS version 8.2 (SAS 2002) in a completely randomized design (CRD) to test the differences among the treatment levels.
RESULTS AND DISCUSSION
The data indicate the differences in the O3 levels at the study sites, where O3 level at the first site (KSU Campus) was 42.33 ppb, which was within the global limits for air pollution by ozone in accordance to McCarthy and Lattanzio (2015); while it was 138.66 ppb in the second site (SIC) indicated higher pollution levels than the global limits for air pollution by ozone. Hence, we expected to detect some effects on the epidermal traits, such as stomatal number and its dimensions, and epidermal cell size.
The results shown in Figures 1 to 11 indicate the differences in the epidermis traits of the studied pepper leaves. The results revealed that there were variations in the stomatal densities (Figures 1 to 8) between the upper and lower epidermal cell of the pepper leaves at King Saud University Campus site (control plants); this trait differed from that of the other plant species (AbdulRahaman and Oladele, 2003). The stomatal responses to the environmental changes are important to maintain the movement of gases and water in and out of the leaves (Hetherington and Woodward, 2003). As O3 enters the plant leaves through the open stomata, the plant controls this process via stomatal closure or decreases stomatal conductance. Hence, the closure of stomata is a mechanism for controlling O3 diffusion into the stomatal chamber for decreasing the O3 concentrations in the mesophyll cells of the leaves (Madkour and Laurence, 2002). The results showed that the stomata of the plants that grew under high concentration of O3 were affected in comparison to control plants (Figures 5 and 6), where the stomatal number increased significantly in both the upper and lower epidermis of the leaf, which indicated that O3 might induce the increase in the stomatal numbers in both the epidermises. This result was consistent with the findings of the previous studies (Frey et al., 1996; Paakkonen et al., 1998; Zouzoulas et al., 2009). The stomatal dimensions decreased significantly in the lower epidermis of the leaf, while the stomatal length increased in the upper epidermis of the leaf of the plant grown under O3. This was in line with the results of different studies (Zouzoulas et al., 2009; Gostin, 2009; Dumont et al., 2014).
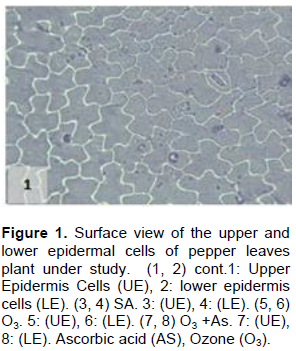
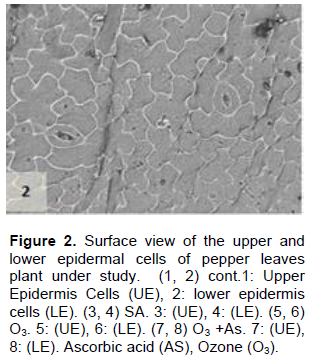
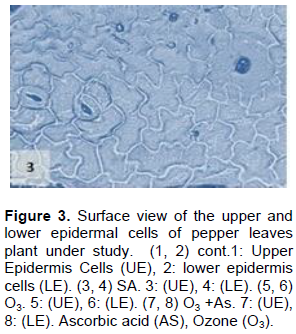
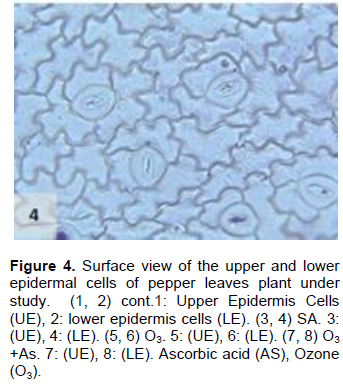
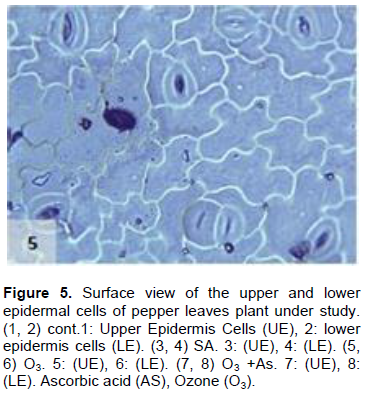
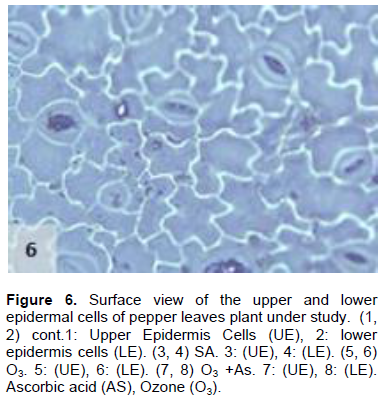
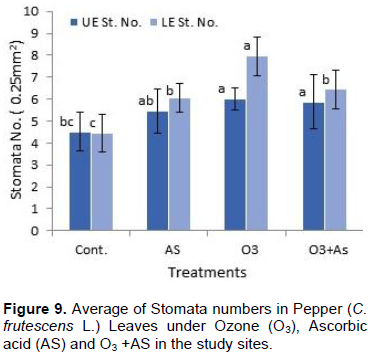
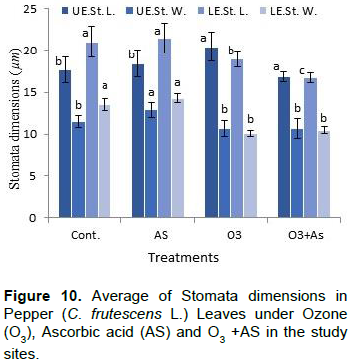
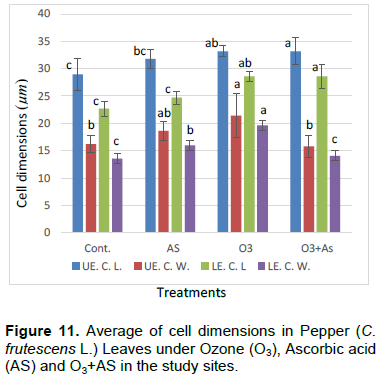
The cell dimensions of the upper and lower epidermises of the leaves of the plants grown under O3 were significantly increased (Figures 5 and 6) in comparison to control plant leaves; the mentioned changes may be due to the exposure to ozone. These observations were consistent with the results of some researchers, who revealed that high concentrations of O3 affected the epidermal cell dimensions (Lawson et al., 2002; Wahid and Ahmad, 2003; Riikonen et al., 2010). The results also showed that AS application increased the number of stomata on both the upper and lower epidermis of the leaves in comparison to the control plant leaves (Figures 3 and 4). However, the increase in the stomatal number was significant only in the lower epidermis; this result was in line with that obtained by Arafa et al. (2014) and Cavusoglu and Bilir (2015). The AS application may have a role in increasing the stomatal connectivity (Hinckley and Braatne, 1994; Dieter et al., 1995). Application of AS also caused an increase in the stomatal width and length in the upper and lower epidermis of the leaf, which was significant only in the upper epidermis. The increase in the stomatal width indicates that AS might have an impact on the guard cells by increasing in its size, thereby increasing the dimensions of the stomata. This finding is consistent with the results of previous research that indicated that AS influences the cell elongation in different parts of the plant body (De Gara et al., 1996; Tommasi et al., 1999; Horemans et al., 2000; Kaviani, 2014; Cavusoglu and Bilir, 2015). This could be due to the effect of AS on the crosslinking between the protein and polysaccharide in the cell wall that leads to loosening of the cell wall. Therefore, cell expansion and elongation (Padh, 1990; Smirnoff, 1996). Figures 1 and 3 show that the epidermal cell shapes were irregular, and the anticlinal cell walls were sinuous/undulate and appeared to be slightly changed, maybe because of the change in the dimensions of the cell.
There was an increase in the dimensions of the upper and lower epidermal cells of the leaves of the plants treated with AS (Figures 3 and 4); however, this increase was not significant in comparison to the leaves of control plant. This result agrees to a certain extent with the findings of Ali (2001), Ali et al. (2015), and Hendi and Boghdady (2016), where they reported that AS induces some anatomical changes in the plants. In addition, the present result was in conformity with the results of previous studies, which reported that AS plays a role in the expansion and elongation of the cells (Padh, 1990; Wang and Faust, 1992; De Gara et al., 1996; Smirnoff and Pallanca, 1996; Tommasi et al., 1999; Horemans et al., 2000; Kaviani, 2014; Cavusoglu and Bilir, 2015).
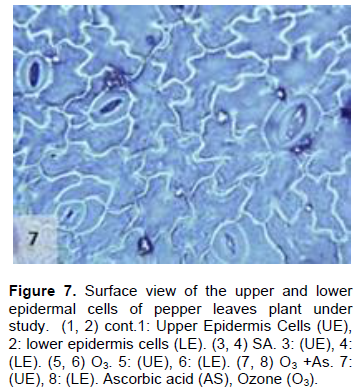
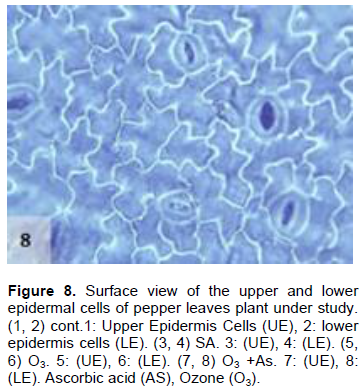
The results obtained also showed that AS application to the plants treated with O3 decreased the stomatal number and their dimensions in both the upper and lower epidermis in comparison to control plant leaves (Figures 7 and 8), which was significant in the lower epidermis. These results may indicate that the effect of AS contradicts with the effect of O3 in reducing the number of stomata in the leaves of the pepper plants. Moreover, the results were inagreement with the findings of previous studies, which reported that AS is a growth regulator and plays important roles in many physiological processes (Kim et al., 2008; Veljovic-Jovanovic et al., 2001; Ejaz et al., 2012). Additionally, AS improves plant tolerance and reduces the harmful effects of stress on plant growth (González-Reyes et al., 1998; Gadalla, 2009; Elwan and El-Hamahmy, 2009). AS also protects the plants from ROS, which are formed during periods of environmental stress associated with O3 exposure (Runeckles and Chevone, 1992; Smirnoff and Pallanca, 1996; Conklin and Barth, 2004; Burkey et al., 2006).
The present results further show that there was a significant increase in the length of the upper and lower epidermis cells in the plants grown under O3 and AS together (O3 + AS) (Figures 7 and 8). On the other hand, there was no significant decrease in the width of the upper and lower epidermis cells of the treated leaves in comparison to control leaves. Plants grown under O3 + AS had significantly increased length of the upper and lower epidermis cells, and significantly decreased width of the cells in comparison to the plants grown under only O3. It appeared that the width of epidermal cells was more responsive to AS in comparison to their length.
CONCLUSION
The results of the present study revealed that high concentration of O3 or AS increased the stomatal numbers and their dimensions, and the cell dimensions in both the upper and lower epidermises of the leaves of the pepper plant (C. frutescens L.) in comparison to the leaves of the control plant. Plants exposed to high concentration of O3 and treated with AS had significantly increased length of the upper and lower epidermal cells. Therefore, we can hypothesize that ascorbic acid may have a mitigating effect on the impact of O3 on the epidermal cell elongation of pepper leaves.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
AbdulRahaman A, Oladele F (2003). Stomatal complex types, size, density and index in some vegetable species in Nigeria. Nigerian Journal of Botany16:144-150. |
|
|
Alabi D (2006). Effects of fertilizer phosphorus and poultry droppings treatments on growth and nutrient components of pepper (Capsicum annuum L). African Journal of Biotechnology 5(8):671-677. |
|
|
Ali Z (2001) Ascorbic acid induced anatomical changes in the leaves and stems of tomato plants. Bulletin of the National Research Centre 26(3):371-382. |
|
|
Ali Z, Hussein M, El-Taher A (2015). Effect of antioxidants on some morphological and anatomical features of maize grown under salinity conditions. International Journal of ChemTech Research 8(6):389-400. |
|
|
Aliyu L (2000). Effect of Organic and Mineral Fertilizers on Growth, Yield and Composition of Pepper (Capsicum annuum L.). Biological Agriculture and Horticulture 18(1):29-36. |
|
|
Arafa A, Khafagy M, Abo- El Kheer A, Fouda R, El-Banna M (2014). Stomatal Density in the Leaves of Sweet Pepper Plant as Affected by Certain Bio-Stimulants Under Salt Stress Conditions. Journal of Plant Production 5(4):649-662. |
|
|
Bray EA (2000) Responses to abiotic stresses. Biochemistry and Molecular Biology of Plants pp. 1158-1203. |
|
|
Burkey KO, Neufeld HS, Souza L, Chappelka AH, Davison AW (2006). Seasonal profiles of leaf ascorbic acid content and redox state in ozone-sensitive wildflowers. Environmental Pollution 143(3):427-434. |
|
|
Cavusoglu K, Bilir G (2015). Effects of ascorbic acid on the seed germination, seedling growth and leaf anatomy of barley under salt stress. Journal of Agricultural and Biological Science 10:124-129. |
|
|
Chauhan A, Joshi P (2010). Effect of ambient air pollutants on wheat and mustard crops growing in the vicinity of urban and industrial areas. New York Science Journal 3(2):52-60. |
|
|
Clark R, Lee SH (2016). Anticancer properties of capsaicin against human cancer. Anticancer Research 36(3):837-843. |
|
|
Conklin PL, Barth C (2004). Ascorbic acid, a familiar small molecule intertwined in the response of plants to ozone, pathogens, and the onset of senescence. Plant, Cell and Environment 27(8):959-970. |
|
|
De Gara L, De Pinto M, Paciolla C, Cappetti V, Arrigoni O (1996). Is ascorbate peroxidase only a scavenger of hydrogen peroxide. Plant peroxidases: biochemistry and physiology University of Geneva, Geneva pp. 157-162. |
|
|
Dieter V, Matyssek R, Herppich W (1995).Experimentielle Pflanzenoekologie. Grundlagen und Anwendungen. |
|
|
Dumont J, Cohen D, GÉRard J, Jolivet Y, Dizengremel P, Le Thiec D (2014). Distinct responses to ozone of abaxial and adaxial stomata in three Euramerican poplar genotypes. Plant, Cell and Environment 37(9):2064-2076. |
|
|
Ejaz B, Sajid ZA, Aftab F (2012). Effect of exogenous application of ascorbic acid on antioxidant enzyme activities, proline contents, and growth parameters of Saccharum spp. hybrid cv. HSF-240 under salt stress. Turkish Journal of Biology 36(6):630-640. |
|
|
El-Zaidy M, Al Musalim A, Al Sahli A, Doaigey A, Yakout SM, Arif IA, Saleh IA (2019). Effects of ozone on cell organelles of alfalfa (Medicago sativa L.) seedlings. Saudi Journal of Biological Sciences. In press. |
|
|
Elwan MWM, El-Hamahmy MAM (2009). Improved productivity and quality associated with salicylic acid application in greenhouse pepper. Scientia Horticulturae 122(4):521-526. |
|
|
Evans LS, Albury K, Jennings N (1996). Relationships between anatomical characteristics and ozone sensitivity of leaves of several herbaceous dicotyledonous plant species at Great Smoky Mountains National Park. Environmental and Experimental Botany 36(4):413-420. |
|
|
Feng Z, Li P, Xu Y, Bo S (2018). The response of water use efficiency in poplar to elevated ozone In: International Conference on Ozone and Plant Ecosystems, 21-25 May, 2018. Florence, Italy. |
|
|
Frey B, Scheidegger C, Gunthardt-Goerg MS, Matyssek R (1996). The effects of ozone and nutrient supply on stomatal response in birch (Betula pendula) leaves as determined by digital image-analysis and X-ray microanalysis. New Phytologist 132(1):135-143. |
|
|
Gadalla S (2009). The role of antioxidants in inducing wheat flag leaf osmotic adjustment under salinity stress. Journal of Agricultural Science - Mansoura University 34(11):10663-10685. |
|
|
Giacomo B, Forino LMC, Tagliasacchi AM, Bernardi R, Durante M (2010) Ozone damage and tolerance in leaves of two poplar genotypes. Caryologia 63(4):422-434. |
|
|
González-Reyes JA, Córdoba F, Navas P (1998) Involvement of Plasma Membrane Redox Systems in Growth Control of Animal and Plant Cells. Plasma Membrane Redox Systems and their Role in Biological Stress and Disease. Springer Netherlands. |
|
|
Gostin IN (2009) Air pollution effects on the leaf structure of some Fabaceae species. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 37(2):57. |
|
|
Hathout T (1995) Diverse effects of uniconazole and nicotinamide on germination, growth, endogenous hormones and some enzymic activities of peas. Egyptian Journal of Physiological Sciences (Egypt). 19(1-2):77-95. |
|
|
Hendi D, Boghdady M (2016). Response of Spearmint Plants Grown under Sandy Soils Condition to some Growth Stimulators. International Journal of PharmTech Research 9(4):92-101. |
|
|
Hetherington AM, Woodward FI (2003) The role of stomata in sensing and driving environmental change. Nature 424(6951):901-908. |
|
|
Hinckley T, Braatne J (1994) Stomata. Plant-environment Interactions pp. 323-355. |
|
|
Horemans N, Foyer HC, Potters G, Asard H (2000). Ascorbate function and associated transport systems in plants. Plant Physiology and Biochemistry 38(7-8):531-540. |
|
|
Iriti M, Faoro F (2008) Oxidative stress, the paradigm of ozone toxicity in plants and animals. Water, Air, and Soil Pollution 187(1-4):285-301 |
|
|
Kaviani B (2014). Effect of ascorbic acid concentration on structural characteristics of appical meristems on in vitro Aloe barbadensis Mill. Acta Scientiarum Polonorum Hortorum Cultus 13:3. |
|
|
Kim JE, Jin DH, Lee SD, Hong SW, Shin JS, Lee SK, Jung DJ, Kang JS, Lee WJ (2008) Vitamin C inhibits p53-induced replicative senescence through suppression of ROS production and p38 MAPK activity. International journal of molecular medicine 22(5):651-655 |
|
|
Lawson T, Craigon J, Black CR, Colls JJ, Landon G, Weyers JDB (2002) Impact of elevated CO2 and O3 on gas exchange parameters and epidermal characteristics in potato (Solanum tuberosum L.). Journal of Experimental Botany 53(369):737-746. |
|
|
Lloyd F (1908). The physiology of stomata, vol . 82. Carnegie Institution of Washington, Washington. |
|
|
Madkour SA, Laurence JA (2002). Egyptian plant species as new ozone indicators. Environmental Pollution 120(2):339-353. |
|
|
McCarthy JE, Lattanzio RK (2015) Ozone Air Quality Standards: EPA's 2015 Revision. Congressional Research Service 29. |
|
|
Mukherjee SP, Choudhuri MA (1985). Implication of hydrogen peroxide - ascorbate system on membrane permeability of water stressed vigna seedlings. New Phytologist 99(3):355-360. |
|
|
Osuna-García JA, Wall MM, Waddell CA (1998). Endogenous Levels of Tocopherols and Ascorbic Acid during Fruit Ripening of New Mexican-Type Chile (Capsicum annuum L.) Cultivars. Journal of Agricultural and Food Chemistry 46(12):5093-5096. |
|
|
Oyagbemi AA, Saba AB, Azeez OI (2010). Capsaicin: A novel chemopreventive molecule and its underlying molecular mechanisms of action. Indian Journal of Cancer 47(1):53. |
|
|
Paakkonen E, Vahala J, Pohjola M, Holopainen T, Karenlampi L (1998). Physiological, stomatal and ultrastructural ozone responses in birch (Betula pendula Roth.) are modified by water stress. Plant, Cell and Environment 21(7):671-684. |
|
|
Padh H (1990). Cellular functions of ascorbic acid. Biochemistry and Cell Biology 68(10):1166-1173. |
|
|
Pasqualini S (2003). Ozone-Induced Cell Death in Tobacco Cultivar Bel W3 Plants. The Role of Programmed Cell Death in Lesion Formation. Plant Physiology 133 (3):1122-1134. |
|
|
Pedroso ANV, Alves ES (2008). Anatomia foliar comparativa das cultivares de Nicotiana tabacum L. (Solanaceae) sensível e tolerante ao ozônio. Acta Botanica Brasilica 22(1):21-28. |
|
|
Riikonen J, Percy KE, Kivimäenpää M, Kubiske ME, Nelson ND, Vapaavuori E, Karnosky DF (2010). Leaf size and surface characteristics of Betula papyrifera exposed to elevated CO2 and O3. Environmental Pollution 158(4):1029-1035. |
|
|
Roshchina VV, Roshchina VD (2013). Ozone and plant cell. Springer Science and Business Media 267 p. |
|
|
Runeckles V, Chevone B (1992). Crop responses to ozone. Surface-level ozone exposures and their effects on vegetation. Lefohn, A.S. (Ed.) edn. Lewis Publishers, Chelsea, Michigan |
|
|
SAS (2002) User's Guide, Statistics (Release 8.02). SAS Inst. Inc, Cary, NC, USA. |
|
|
Shotorbani NY, Jamei R, Heidari R (2013). Antioxidant activities of two sweet pepper Capsicum annuum L. varieties phenolic extracts and the effects of thermal treatment. Avicenna Journal of Phytomedicine 3(1):25. |
|
|
Smirnoff N (1996). Botanical briefing: The function and metabolism of ascorbic acid in plants. Annals of Botany 78(6):661-669. |
|
|
Smirnoff N, Pallanca JE (1996) Ascorbate metabolism in relation to oxidative stress. Biochemical Society Transactions 24(2):472-478. |
|
|
Tommasi F, Paciolla C, Arrigoni O (1999). The ascorbate system in recalcitrant and orthodox seeds. Physiologia Plantarum 105(2):193-198. |
|
|
Udoh DJ, Ndon BA, Asuquo PE, Ndaeyo N (2005). Crop production techniques for the tropics. Ibadan: Concept Publications Limited. |
|
|
Veljovic-Jovanovic SD, Pignocchi C, Noctor G, Foyer CH (2001). Low Ascorbic Acid in the vtc-1 Mutant of Arabidopsis Is Associated with Decreased Growth and Intracellular Redistribution of the Antioxidant System. Plant Physiology 127(2):426-435. |
|
|
Wahid A, Ahmad S (2003). Environmental pollution and its effects on chilli (Capsicum annuum L.) grown in fields of Lahore [Pakistan]. Biologia (Pakistan). |
|
|
Wang SY, Faust M (1992). Ascorbic acid oxidase activity in apple buds: relation to thidiazuron-induced lateral budbreak. HortScience 27(10):1102-1105. |
|
|
Wittig VE, Ainsworth EA, Naidu SL, Karnosky DF, Long SP (2009). Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis. Global Change Biology 15(2):396-424. |
|
|
Zouzoulas D, Koutroubas SD, Vassiliou G, Vardavakis E (2009). Effects of ozone fumigation on cotton (Gossypium hirsutum L.) morphology, anatomy, physiology, yield and qualitative characteristics of fibers. Environmental and Experimental Botany 67(1):293-303. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


