Full Length Research Paper

ABSTRACT
The variations in seed quality and scarcity vigor studies have been limiting factors in crop establishment at different regions of its Andes origin. The present study aimed to evaluate the physiological quality of quinoa seeds, from cultivars of Brazil, Colombia and Ecuador. The research was conducted in the Seeds Technology Laboratory of the University of Brasília, Brazil, through the tests of germination, hypocotyl and radicle length and accelerated aging test. Eighteen genotypes selected from different cultivars were evaluated using the F Test and Scott-Knotts. In addition, hierarchical cluster analysis was performed using the Ward method and Cluster analysis by k-means. The germination average was 80.81%; the hypocotyl and radicle length characteristics showed an average of 9.11 and 11.27 mm, respectively. The average germination after the accelerated aging test was 75.52%, with better results for the genotypes Piartal and Aurora. Quinoa genotypes showed variability in seed vigor, allowing determining differences in germination potential at high stress conditions. The application of methods developed for the analysis of quinoa seeds at 25°C and 4 days has been effective in screening for quality. Seed quality assessments in quinoa allow defining limits of tolerance to adverse conditions, facilitating selection in breeding programs.
Key words: Accelerated aging test, Chenopodium quinoa, germination, selection, vigor.
INTRODUCTION
Quinoa (Chenopodium quinoa Willd.) is a plant in the Amaranthaceae family, subfamily Chenopodioideae, as well as spinach and beets (Spehar et al., 2014). It has been cultivated for many centuries in the Andean countries, with greater predominance in Peru and Bolivia (Pereira et al., 2019). Due to its high content of vitamins (A, B2, E), minerals (Ca, Fe, Cu, Mg and Zn) and the stability of its amino acids, the cultivation of quinoa has expanded in different regions (Schabes and Sigstad, 2005). Currently, it is being cultivated in countries like Canada, China, England, Australia, India, Pakistan, United States, among others (Aziz et al., 2018).
The first attempts to introduce quinoa in Brazil, as a cultivated plant, began in the 1990s, evaluating hybrid lines between cultivars from the Andean altiplano, sensitive to the day length (Spehar et al., 2014). With the potential, quinoa germplasm was increased by introducing genotypes with higher genetic variability, coming from several countries, such as the Andes and the United States, including genotypes with large saponin-free seeds. The high rate of cross-pollination in the savanna allowed the recovery of hybrid vigor and the generation of progenies from individual plants, in which agronomic characteristics and nutritional quality were evaluated (Spehar et al., 2014).
These efforts resulted in the cultivars BRS Piabiru and BRS Syetetuba (Spehar et al., 2014). However, recently, these first cultivars showed seed quality problems (Souza et al., 2017a). Quinoa seeds are of the achene type, and have an external structure made of dead cells that surround the seeds (Burrieza et al., 2014), which can interfere with their germination (Souza et al., 2017a). Therefore, it is still necessary to assess seed germination and identification, to detect differences in the physiological potential, seeking to select high-performance materials for successful agricultural production (Marcos Filho, 2015). Climate change and population growth generate the need to use cultures and genotypes resistant to biotic and abiotic stress, with high yield and high nutritional properties (Ruiz et al., 2017). Thus, the cultivation of quinoa can be a solution to increase food for the population, since it has great resistance to adverse production conditions.
In laboratory conditions, at 20 and 30°C, the germination of quinoa seeds is optimized using absorbent paper, with an initial counting at two days and a final at four days. These standards have been used aiming at genotype characterization and selection in the tropical savannahs. They have been developed in conjunction with the accelerated ageing test, which comprises various molecular processes, causing degradation of large seed molecules related to germination and vigor (Roach et al., 2018). In general, vigorous seeds have shown resistance to the stress factors, exhibiting longer longevity and higher field emergence (Marcos Filho, 2015).
The aim of this study was to evaluate the physiological quality of quinoa seeds of the valley-type cultivars originated in Brazil, Colombia and Ecuador, with the purpose of finding variability in vigor characteristics, allowing to determine differences in germination potential under high stress conditions.
MATERIALS AND METHODS
The experiment was conducted in the Seed Technology Laboratory of the Faculty of Agronomy and Veterinary Medicine, University of Brasilia, Campus Darcy Ribeiro, Brasilia, DF, Brazil. Seeds of quinoa progenies selected in Brazil were grown in 2018, Departamento del Cauca, Comlombia Ecoaldea Fundamor, Santander de Quilichao, located at 1,100 m altitude, 3° 0 '30 "NL and 76° 29' 2" WL and Hacienda Prosperidad, Popayán, 1,850 m altitude, at 2° 27'NL and 76° 37' 18 " WL. The respective mean temperatures of the two locations were 21C and 19°C.
Plants harvested manually when seeds matured, contained approximately 20 to 30% wet base (w.b.) moisture and let dry at environment conditions until reached 12% w.b., being threshed and cleaned before storage in cold chamber at 10°C. Before proceeding with tests, seeds were immersed in 2% sodium hypochlorite solution for 10 min and rinsed with distilled water to prevent infection by possible seed borne fungi. A brief description of genotypes used in experiment is as follows: (a) Aurora, cultivar selected by the Facultad de Ciencias Agrícolas, Universidad de Nariño, Colombia, adapted to 2,300 to 3,000 m altitude, having short cycle (110 days to maturity) and average plant height of 1.15 m. Similar to BRS Syetetuba, the perigonium opens at maturity, exposing the white colored grains with diameter smaller than 2 mm (Sañudo et al., 2005). (b) Blanca de Jericó, selected in Boyacá, Colombia at 3,100 m altitude, average plant height of 1.70 m, yellow panicle at maturity, with white pericarp and low saponin content (Montes et al., 2018). (c) BRS Syetetuba, cultivar developed in Brazil at 1,100 m altitude, with an average plant height of 1.8 m and a yellow panicle at maturity, 3.0 g for the weight of 1,000 seeds, 18 g of protein per 100 g seeds, white pericarp and saponin absence; at maturity, the external structure of the perigonium that involves the fruits, opens and makes them visible at maturity (Spehar et al., 2014). (d) Piartal cultivar, selected in the Carchi province, Northern Ecuador, at 2,900 m altitude, with an average plant height of 2.0 m, purple panicle at maturity and white fruits with 2 mm in diameter, these being susceptible to downy mildew (Peronospora farinosa f.sp. chenopodii) (Alvarez et al., 1990).
Progenies of selected individual plants were obtained: BRS Syetetuba 8T1S, 8T2S, 8T1P, 8T2P, 4T1S, 4T2S, 4T1P, 4T2P and ST; Blanca de Jericó BLA; Piartal PT1S, PT2S, PT1P and PT2P; and Aurora AT2S, AT1P, and AT2P. All cultivars and progenies have typical quinoa shape, cylindrical and flat achene fruit which has been treated as seed.
Seeds of selected progenies were placed on blotter paper sheets, previously humidified with distilled water 2.5 times the paper dry weight in 11 × 11 × 3 cm plastic box (Gerbox), moved to biochemical oxygen demand chamber (BOD), set at 25°C, on 75° inclination angle, being kept for 48 h (Nakagawa, 1999; Souza et al., 2017a).
For the accelerated ageing test, 250 seeds taken for each progeny and distributed in a single layer arranged stainless-steel screen inserted in plastic boxes (Gerbox) covered with a lid, containing 40 mL of distilled water in BOD, regulated to 41 ± 1°C for 24 h (Barbosa et al., 2011). Each treatment was tested individually to avoid possible interference that could be caused by position in chamber. At the end of exposure, a sample of 200 seeds was taken for each progeny and divided in four subsamples for germination test.
The experimental design consisted of a complete randomized block design with four repetitions of 50 seeds each. The following characteristics were evaluated 48 h after the beginning: first count germination (G1), germination rate (% G - primary root emission), radicle length (mm) and e hypocotyl length (mm).
The data were subject to analysis of variance and significance of treatments assessed by F test (p ≤ 0.05 and p ≤ 0.01), while means were compared by Scott-Knott (p ≤ 0.05). The statistical analyses were conducted in SISVAR 5.3 program (Ferreira, 2011). Similarities among genotypes were assessed by hierarchical cluster (Ward, 1963) and clusters compared by k-means (Mac Queen, 1967).
RESULTS AND DISCUSSION
The progenies were significantly different for germination in both standard and accelerated ageing test, radicle length and hypocotyl length as shown by the F test from the analysis of variance (Table 1).

The low value coefficient of variation demonstrates experimental precision in controlled conditions. Germination rate mean for all progenies was 80.81%, although seeds of progenies BLA and 4T2S had considerably lower germination values (Table 2).
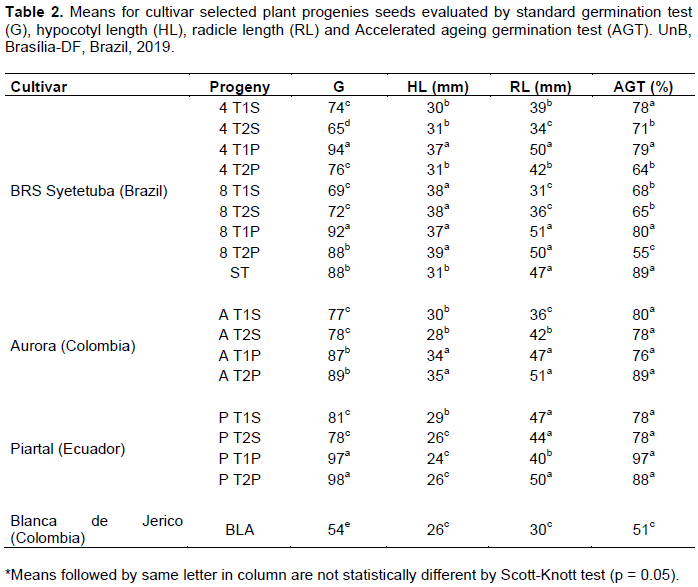
The comparatively lower germination rate of genotypes BLA and 4T2S may have been related to high moisture field conditions in Santander de Quilichao at their seed maturity. The field is located at 1,100 m altitude, with mean temperatures of 21°C. Therefore, under lower mean temperature, seed quality is expected to be high, confirmed by the accelerated ageing test. Quinoa seeds are liable to variations in temperature and moisture, relating to rapid deterioration (Spehar et al., 2014).
Immediately after harvesting, drying, and cleaning, seeds had grain moisture lowered to 12%, before being stored in cold chamber at 10°C (Souza et al., 2017b). This procedure justifies the high germination rates higher than 70% for most progenies. Moreover, the lower germination rate for the seeds submitted to accelerate ageing test indicates that some progenies are more liable to lose germinability under moisture and temperature stress at maturity. This has been shown by statistical differences picked up by Scott-Knott test of means. Germination rate was an indication of germinability not always associated to seedling vigor, as measured by hypocotyl length and radicle length.
The progenies PT2P, PT1P, 4T1P, 8T1P and AT2P had the best germination rate, confirmed by the accelerated ageing test. The storage at 10°C assured their seeds kept the quality associated with cropping, harvesting, drying, and processing conditions (Souza et al., 2017b). In addition, progeny selections PT2P and PT1P from cultivar Piartal of Northern Ecuador with germination rates of 97 and 96% and respectively 97 and 88% after being exposed to AGT. When seeds are mature under low moisture in the field, it has been found that they keep high germination and vigor (Spehar et al., 2014). This was confirmed by radicle length, whereas the hypocotyl length was relatively smaller than progenies derived from BRS Syetetuba and Aurora cultivars. This is an indication of their common origin as BRS Syetetuba was first selected progeny 4.5 from quinoa population of Northern Ecuadorian valleys (Spehar and Rocha, 2009). Similar results were obtained with five batches of BRS Syetetuba (Souza et al., 2017a). The Colombian Aurora had also high germination rates AT1P: 86.50% and AT2P: 89% (Table 2). This cultivar was selected in Nariño, on similar climatic conditions as neighboring Ecuador, and described as having high quality seed quality (Montes et al., 2018). Thus, hypocotyl length used to define seed quality in quinoa (Anzooman et al., 2018) does not hold true for the progenies of this experiment.
Evaluations on the fourth day of experiment, confirmed as the best for discriminating for seed quality (Souza et al., 2017a). Genotypes originated in high moisture stress environment have shown best results for standard germination test at the fourth day (Makinen et al., 2014). The high imbibition capacity of seeds leads to rapid radicle protrusion, between 6 and 12 h, after being exposed to moisture (Makinen et al., 2014; Souza et al., 2017b). Quinoa originated, evolved, and was domesticated in environment of low temperature and moisture. Therefore, its adaptation to high moisture and temperature depends on selection for seed quality under these conditions (Spehar et al., 2014).
The significant higher value for hypocotyl length in genotypes derived from BRS Syetetuba (Table 2), confirmed results in other study (Souza et al., 2017). This contrasts with results of native Argentinian native accessions, for which hypocotyl length varied between 13.93 and 22.15 mm, much lower than the ones of this experiment (Curti et al., 2010).
As for radicle length, there were statistical differences within and among progenies (Table 2). In all progenies, the radicle had ≥ 2 mm length, similar to the obtained in experiment using LED and fluorescent light (Sanoubar, 2018).
Progenies derived from BRS Syetetuba had higher radicle length as T1P 51.35 mm, 8T2P 50.15 mm and 4T1P 50.17 mm, followed by progenies of Colombian cultivar Aurora as AT2P 50.47 mm and Ecuadorin Piartal PT2P 49.62 mm. Their possible common genealogy would explain the similar behavior (Spehar and Rocha, 2009; Alvarez et al., 1990). Differences in physiological seed quality have been detected by complimentary testing as accelerated ageing method (Marcos Filho, 2015). A lower mean value was expected as reported here (Table 2). This has been used to assess quinoa seed vigor (Soares and Broges, 2017). In these tests, some genotypes showed resilience, responding equally
in standard germination test and accelerated ageing test, performing above the mean (75.52%). Significant differences among progenies of cultivars indicate the possibility to select quinoa to adverse field conditions, where seeds may gain and lose water before ripening (Sigstad and Garcia, 2001).
The hierarchical clustering allowed grouping the progenies for similarities considering the characteristics measured in the testing (Figure 1). The group division was related to Euclidean distance among detected differences (Freddi et al., 2008). The progenies were grouped in four clusters, following the Euclidean distance of 5 (Hair et al., 2005). Cluster 1 was formed by 5 progenies, 3 of BRS Syetetuba (4T1P, 8T1P and 8T2P) and two of Aurora (AT1P and AT2P). Their similarity confirmed results with germination and vigor tests. Cultivar BRS Syetetuba has shown seed quality in the Brazilian Savannah when plant senescence occurs in dry conditions, desirable for commercial production (Spehar et al., 2014).

In cluster II, there were three genotypes progenies, one of BRS Syetetuba (ST) and two of Piartal (PT1S and PT2S). These consisted of a cluster of progenies that had lower seed quality relative to the ones of cluster I.
The cluster III had 7 progenies, three of which derived from BRS Syetetuba (4T1S, 4T2S and 4T1P), two of Piartal (PT1S and PT2S) and two of Aurora (AT1S and AT2S). This cluster showed lower physiological seed quality than clusters I and II.
The cluster IV had the three remaining progenies, two of BRS Syetetuba (8T1S and 8T2S) and the Blanca de Jerico (BLA). These progenies represented the extreme of low seed quality of the progenies.
In the grouping by k-means, each measurement was weighed taking the means for standard germination test, hypocotyl length, radicle length and accelerated ageing germination test.
The lower the p-value the higher the capacity of differentiating the groups for a given set of variables (Coelho and Toledo, 2017). In this study, standard germination test, radicle length and accelerated ageing germination test were the key characters to segregate the clusters (Table 3). Accelerated ageing test has successfully been used to discriminate progenies of turnip in response to high temperature (Oliveira et al., 2014).

Genotypes of cluster I had mean germination value of 90%, 50 mm radicle length and 76% in accelerated ageing germination test (Figure 2).

Germinability must be considered in progeny selection for seed quality (Spehar et al., 2014). The apparent variability in the progenies of this study indicates they have a common genetic background based on parental genealogy (Spehar and Rocha, 2009; Alvarez et al., 1990). Under growing conditions of the Brazilian Savannahs, seed quality is important trait in grain crops to turn them viable (Carvalho and Nakagawa, 2012; Oliveira et al., 2014).
The distance between cluster points for each seed characteristic showed high germination rate was associated with physiological quality of seeds (Figure 2). The opposite was found for the low seed progenies cluster, with emphasis on germination and accelerated ageing. In fact, a quality seed is expected to possess germinability and vigor to generate homogenous plant population in the field (França Neto et al., 2010; Wendt et al., 2014). From the results presented here it is inferred that quinoa can be improved for seed quality by exploiting variability in genotypes originated from high moisture and temperature environmental stresses. The differences could be picked up in progenies aiming to acquire improved genotypes for seed quality (Spehar et al., 2014).
It should be stated that quinoa crop has a relatively short story in the Brazilian Savannah commercial production. The scientific studies have been limited to genotype introduction and evaluation, with little investment on artificial hybridization and selection. The identification of genetic sources of seed quality shall ensure identification of progenies to compose of a breeding program. The seed characteristics common to Brazilian, Colombian, and Ecuadorian cultivars identified here offers opportunity to cooperative breeding to reach the purpose and adapt the crop to less-favorable climatic conditions.
CONCLUSIONS
Progenies obtained from individual plants selected in quinoa cultivars adapted to environments of higher moisture and temperature show variability in seed germination and vigor and differences in quality.
The quality seed progenies have the potential to germinate and establish when submitted to high stress and to adapt in similar environments.
Seed quality evaluation in quinoa is useful to identify limits of tolerance to adverse climatic conditions and to select progenies for genetic improvement.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENT
Universidade de Brasília (UnB) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) were acknowledged in providing resources to accomplish the work.
REFERENCES
|
Alvarez M, Pavón J, Von Rütte S (1990). Caracterización. In: WAHLI, C. Quinua: hacia su cultivo comercial pp. 5-30. |
|
|
Anzooman M, Dang YP, Christopher J, Mumford M H, Menzies NW, Kopittke PM (2018). Greater emergence force and hypocotyl cross sectional area may improve wheat seedling emergence in sodic conditions. Plant Science 277:188-195. |
|
|
Aziz A, Akram N A, Ashraf M (2018). Influence of natural and synthetic vitamin C (ascorbic acid) on primary and secondary metabolites and associated metabolism in quinoa (Chenopodium quinoa Willd.) plants under water deficit regimes. Plant Physiology and Biochemistry123:192-203. |
|
|
Barbosa RM, Costa DS, Sá ME (2011). Envelhecimento acelerado em sementes de alface. Ciência Rural 1:1899-1902. |
|
|
Burrieza HP, López FMP, Maldonado S (2014). Analogous reserve distribution and tissue characteristics in quinoa and grass seeds suggest convergent evolution. Frontiers in Plant Science 5:546. |
|
|
Carvalho NM, Nakagawa J (2012). Sementes: ciência, tecnologia e produção. 5.ed. Jaboticabal: FUNEP 590 p. |
|
|
Coelho RP, Toledo JC (2017). Programas para segurança na indústria de alimentos para animais: caracterização e benefícios percebidos com a implantação. Gestão e Produção 24:704-718. |
|
|
Curti RN, Bertero D, Andrade A, Belasquez B, Bramardi S (2010). Caracterización morfo-fenológica preliminar del germoplasma nativo de quinoa (Chenopodium quinoa Willd.) del noroeste argentino (NOA). |
|
|
Ferreira DF (2011). Análises estatísticas por meio do Sisvar para Windows versão 4.0. In: 45a Reunião Anual da Região Brasileira da Sociedade internacional de Biometria 45(2000):235. |
|
|
França Neto JB, Krzyzanowski FC, Henning AA (2010). A importância do uso de sementes de soja de alta qualidade. Informativo Abrates, Londrina 20:37-38. |
|
|
Freddi OS; Ferraudo AS, Centurion JF (2008). Análise multivariada na comparação de um latossolo vermelho cultivado com milho. Revista Brasileira de Ciência do Solo 32:953-961. |
|
|
Hair JF, Anderson RE, Tatham RL, Black W (2005). Análise Multivariada De Dados. Porto Alegre, Bookman editora. |
|
|
Mac Queen JB (1967). Some Methods for classification and analysis of multivariate observations. Proceedings of 5th Berkeley Symposium on Mathematical Statistics and Probability. 1. University of California Press. pp. 281-297. MR 0214227. Zbl 0214.46201. |
|
|
Makinen OE, Hager AS, Arendt EK (2014). Localisation and development of proteolytic activities in quinoa (Chenopodium quinoa) seeds during germination and early seedling growth. Journal of Cereal Science 60(3):484-489. |
|
|
Marcos FJ (2015). Seed vigor testing: an overview of the past, present and future perspective. Scientia Agricola 72(4):363-374. |
|
|
Montes RC, Burbano CGA, Munoz CEF, Calderón Y (2018). Descripción del ciclo fenológico de cuatro ecotipos de (Chenopodium quinua Willd.), en Puracé-Cauca, Colombia. Biotecnología en el Sector Agropecuario y Agroindustrial, Popayán- Colombia 16(2):26-37. |
|
|
Nakagawa J (1999). Testes de vigor baseados no desempenho das plântulas. Vigor de sementes: conceitos e testes. Londrina: ABRATES 1:1-24. |
|
|
Oliveira SSC, Martins CC, Cruz SJS, Silva CJ (2014). Seleção de progênies de nabo-forrageiro para germinação sob altas temperaturas. Ciência Rural, Santa Maria 44(2):217-222. |
|
|
Pereira E, Encina ZC, Barros L, Gonzales BU, Cadavez V, Ferreira ICFR (2019). Chemical and nutritional characterization of Chenopodium quinoa Willd (quinoa) grains: A good alternative to nutritious food. Food Chemistry 280:110-114. |
|
|
Roach T, Nagel M, Börner A, Eberle C, Kranner I (2018). Changes in tocochromanols and glutathione reveal differences in the mechanisms of seed ageing under seedbank conditions and controlled deterioration in barley. Environmental and Experimental Botany 156:8-15. |
|
|
Ruiz KB, Rapparinib F, Bertazzab G, Silva H, Torrigianid T, Biondia S (2017). Comparing salt-induced responses at the transcript level in a salares and coastal-lowlands landrace of quinoa (Chenopodium quinoa Willd). Environmental and Experimental Botany 139:127-142. |
|
|
Sanoubar R, Calone R, Noli E, Barbanti L (2018). Data on seed germination using LED versus fluorescent light under growth chamber conditions. Data in Brief 19:594-600. |
|
|
Sañudo B, Arteaga G, Betancourth C, Zambrano J, Burbano J (2005). Perspectivas de la quinua dulce para la región andina de Nariño. Pasto: Unigraf, Colombia. |
|
|
Schabes FI, Sigstad EE (2005). Calorimetric studies of quinoa (Chenopodium quinoa Willd.) seed germination under saline stress conditions. Thermochimica Acta 428:71-75 |
|
|
Sigstad EE, Garcia CLA (2001). Microcalorimetric analysis of quinoa seeds with different initial water content during germination at 25 °C. Thermochimica Acta 366(2):149-155. |
|
|
Soares VN, Borges CT (2017). Produção de Sementes de Quinoa no Sul do Rio Grande do Sul. Embrapa Clima Temperado-Comunicado Técnico (INFOTECA-E). |
|
|
Souza FFJ, Souza JEA, Souza NOS, Spehar CR, Jesus TF (2017a). Standardizing germination tests for quinoa seeds. African Journal of Agricultural Research 12:155-160. |
|
|
Souza FFJ, Spehar CR, Souza NOS, Fagioli M, Souza RTG, Borges SRS (2017b). Accelerated ageing test for the evaluation of quinoa seed vigour. Seed Science and Technology. Seed Science and Technology 45(1)212-221. |
|
|
Spehar CR, Rocha JES (2009). Effect of sowing density on plant growth and development of quinoa, genotype 4.5, in the Brazilian savannah highlands. Bioscience Journal 25(4):53-58. Spehar C, Rocha JES, Ribeiro JWQ, Santos RDB, Ascheri J, Souza FDJ (2014). Avances y desafíos de la producción y utilización de la quinua en Brasil. In: BAZILE, D., et al. Estado del arte de la quinua en el mundo en 2013. FAO (Santiago de Chile) y CIRAD, (Montpellier, Francia) pp. 681-706. |
|
|
Ward JH (1963), Hierarchical Grouping to Optimize an Objective Function. Journal of the American Statistical Association 58(301):236-244. |
|
|
Wendt L, Gomes JFG, Zorato MF, Moreira GC (2014). Avaliação do potencial fisiológico de sementes de soja por meio de imagens. Pesquisa Agropecuária 44(3):280-286. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


