Full Length Research Paper

ABSTRACT
Eucalyptus cultivation has expanded considerably in Brazilian systems, leading to the current search for technologies to optimize growing conditions and the production of seedlings in nurseries. Based on the understanding that the development of tree species such as Eucalyptus sp. can be influenced by endophytic fungi that act directly as plant growth-promoting species, cloned seedlings of Eucalyptus grandis x Eucalyptus urophylla hybrids grown from minicuttings we stimulated with three species of endophytic fungi and the effects of inoculation on seedling growth was evaluated. Strains of Trichoderma sp., Fusarium sp. and Papulaspora sp. were forced to colonize the root system of the plants, which were continuously maintained under protected cultivation. Inoculation of the symbionts had positive effects on stem length, stem diameter and the fresh and dry biomass of the treated plants. Non-inoculated plants presented a shorter stem length than the plants treated with any of the endophytic species. The cloned seedlings inoculated with Trichoderma sp. exhibited the greatest stem measurements at 120 days after transplanting. The seedlings inoculated with Fusarium sp. displayed a greater number of leaves than the other seedlings as well as greater amounts of fresh and dry biomass. The authors also conducted quarterly evaluations of the increment in seedling growth promoted by the inoculants, which were more effective in the early stages, up to 60 days after transplanting.
Key words: Inoculants, microorganisms, minicutting, tree species.
INTRODUCTION
Many microorganisms, especially those associated with roots, have historically demonstrated the ability to promote plant growth and productivity (Chang et al., 1986; Kloepper et al., 1988). The numerous effects of such plant-microorganism associations include an increased N2 fixation capacity (Kuss et al., 2007; Lammel et al., 2013; Richardson et al., 2009) and the provision of solubilized nutrients, such as phosphates (Anzuay et al., 2013), where the solubilizing ability of many fungi has been associated with the release of organic acids and a decrease in pH. Gains in plant growth have also been related to facilitated absorption of iron promoted by certain microorganisms (Pii et al., 2015; Sadeghi et al., 2012) that synthesize siderophores (low molecular weight iron-chelating molecules) (Lemanceau et al., 2009; Miethke and Marahiel, 2007). Siderophores act as solubilizing agents of iron from minerals or organic compounds under iron-limited conditions (Rajkumar et al., 2010). The efficiency of siderophores from Trichoderma asperellum in donating Fe to plants grown under iron deficiency was demonstrated by De Santiago et al. (2009).
In addition to all of processes described above, some fungi have demonstrated the ability to synthesize phytohormones that are directly associated with plant development, such as auxins and gibberellins. In a study conducted in Arabidopsis thaliana, Contreras-Cornejo et al. (2011) found that Trichoderma virens produced compounds related to auxin, such as indole-3-acetic acid, indole-3-acetaldehyde and indole-3-ethanol. This phytohormone is known to act in the division and cell elongation process, since it activates enzymes that act on components of the bonds between cell wall cellulose microfibrils, increasing their plasticity (Castro et al., 2001). Consequently, water enters the cells more easily and rapidly increases their size (Taghavi et al., 2009).
There are few studies demonstrating the involvement of endophytes in promoting the growth of perennial plants such as those of the genus Eucalyptus. Cultivation of Eucalyptus has increased considerably in recent decades, associated with the use of various technologies and information on the edaphoclimatic diversity of cultivation areas (Gonçalves et al., 2014). Large forest stands of Eucalyptus, which is wood intended mainly for the civil construction and furniture industry, are distributed across all regions of Brazil, accounting for approximately 70% of the total planted forest area (ABRAF, 2013). Although, vegetative propagation of superior genotypes is employed in intensive clonal Eucalyptus forestry, this type of reproduction causes a number of problems, including variable results for the rooting index of cuttings and diseases acquired during this process (Díaz et al., 2009; Lombard et al., 2010). In this context, all technologies that optimize the growing conditions and production of ministumps directly favor the productive capacity of a nursery. Various agricultural strategies can be employed to improve the production of seedlings, including the use of bio-fertilizers, which are prepared organominerals rich in bacteria (Mehnaz, 2014; Sharma and Chaubey, 2015). Currently, the most widespread bacterial fertilizers are produced based on rhizobacteria (Dias et al., 2012; Melo et al., 2012; Vitorazi Filho et al., 2012). For species of the genus Eucalyptus, studies have related the incorporation of rhizobacteria in the substrate with increased rooting of minicuttings, growth after rooting and gains in dry mass (Alfenas et al., 2009; Díaz et al., 2009; Mafia et al., 2009), and these effects are significant when compared with those caused by the treatment of cuttings with indole butyric acid (IBA) (Teixeira et al., 2007). It is also known that there are many environmental and genetically effects such as nursery practice and seed source in tree seedling growth (Dilaver et al., 2015; Yazici et al., 2011; Yazici and Babalik, 2011, 2016).
Since eucalyptus plantations are located in low-fertility soils in Brazil (Gama-Rodrigues et al., 2005), studies that take into account fertilization and nutrient cycling techniques are important for increasing and maintaining forest production. Thus, this study evaluated the effect of inoculation with three endophytic fungi (Trichoderma sp., Fusarium sp. and Papulaspora sp.) on the growth of cloned seedlings of Eucalyptus grandis x Eucalyptus urophylla hybrids grown from minicuttings.
MATERIALS AND METHODS
Acquisition of plant material
The plant material was obtained via the cloning of E. grandis x E. urophylla hybrids grown from minicuttings. Shoots were collected from a clonal minigarden grown in a sand bed and were approximately one year old. The shoots were maintained in Tri-Mix substrate for the adaptive period of two weeks in a protected environment. To set up the experiment, the seedlings were transferred to 3.5-L pots containing Tri-Mix that had previously been sterilized in an autoclave at 121°C (Figure 1A and D).

Acquisition of fungal isolates
Endophytic isolates were previously obtained from the roots of Hyptis marrubioides, a medicinal plant native to the Cerrado (Brazilian savanna) (Vitorino et al., 2012). These isolates were maintained in flasks containing nutrient agar culture medium and stored in the culture collection of the Laboratory of Agricultural Microbiology of the Federal Institute of Goias (IF Goiano), Rio Verde Campus. The isolates were activated in PDA (infusion of 200 g potato, 20 g dextrose, 15 g agar and water up to 1000 mL) (Figure 1C). Mycelial disks of approximately 0.5 cm in diameter were removed from the established colonies and carefully distributed inside the planting holes, where they were placed in contact with the root system of the seedlings at the time of transplantation to the plastic pots (Figure 1D).
Three species of endophytic fungi were tested (Trichoderma sp., Fusarium sp. and Papulaspora sp.), where the control treatment consisted of plants free of microorganism inoculation. The experimental design was completely randomized, and the four treatments (the three fungi and a control) were analyzed in seven replicates. Growth evaluations of stem length (SL), stem diameter (SD), number of leaves (NL), root length (RL), fresh biomass (FB), dry biomass (DB) and leaf area (LA) were performed. The biometric analyses were conducted using a caliper and an analytical balance. To calculate the leaf area, photographs were taken with a digital camera using a reference for size comparison. A white piece of paper with centimeter markings was employed as the reference, upon which the leaves of the plants were placed to take the photographs. The photographs were then analyzed using the software Sigma Scan Pro- V.S. O, Jandel Scientific.
The quarterly values for stem length and stem diameter were used to determine the percentage growth increase of the seedlings via the following equation:
Where, b = value of the variable on the assessment day; a = value of the variable in the previous assessment.
RL, FB and LA were measured only at 120 days, and mean DB values were obtained after six days of drying in a forced air oven at 65°C, when the samples reached constant weight. The growth data were compared with the values recorded for the seedlings on the day of transplantation. The means for the different treatments were subjected to analysis of variance (ANOVA) using the F-test, and when significant differences were found, the means corresponding to FB, DB and LA were compared using Tukey’s test at the 5% probability level, while the means for SL, SD and NL were compared through regression analysis. The data for SL obtained from the inoculation treatments were compared with the control treatment using a t test (inoculated plants x non-inoculated plants) for the different evaluation times. Statistical tests were performed with the aid of the R statistical package (R Core Team, 2016).
RESULTS
When evaluating the capacity of the three endophytic fungal isolates to promote the growth of cloned E. grandis × E. urophylla seedlings, analysis of variance of the means for the biometric variables revealed the significance of the treatments for SL, SD, FB and DB, while no difference was detected between the inoculation treatments and the control for LA. Additionally, differences were observed in relation to the evaluation times (0, 30, 60, 90 and 120 days after transplanting) for SL, SD and NL. The interaction between the different types of fungi used for inoculation and the evaluation times was not significant. This finding indicates independence of these variables, according to the F-test (Table 1).

For SL, the regression analysis was significant for time (R2 = 95.1) and treatment (R2 = 30.5). Breakdown of the time variable within the different treatments revealed significance for this growth parameter only at 90 and 120 days after transplanting (p = 0.0003 and p = 0.0000, respectively). For SD, the regression was also significant for time (R2 = 81.3) and treatment (R2 = 20.1), where the breakdown was significant at 30, 60 and 90 days (p = 0.0005, 0.0103 and 0.0001, respectively). Regarding NL, the regression analysis was significant for treatment (R2 = 86.6) and the interaction, where the breakdown showed significance for all time periods evaluated (p = 0.0093, p = 0.0067, p = 0.0072 and p = 0.0006).
The behavior of the different variables as a function of the treatments is shown in Figure 2. A linear effect was observed (a typical demonstration of a temporal increase in SL growth) in all treatments. For SD, linearity was also obtained for the data corresponding to the plants inoculated with Papulaspora sp. and Trichoderma sp.; however, a quadratic effect on the growth of the control plants as well as those inoculated with Fusarium sp. was detected. A quadratic effect was also found for the variable NL in all the treatments as well as the control.

The plants inoculated with the tested endophytic fungi exhibited better stem growth when compared with the control plants (Figure 2A), where the plants inoculated with Trichoderma sp. presented the greatest measurements at the end of the evaluation period. The results of the t test (Table 2) for SL indicated significant growth of plants inoculated with the endophytes when compared with non-inoculated plants, confirming the benefit of the plant-endophyte association as well as the growth-promoting character of the evaluated species.

The cloned seedlings inoculated with Fusarium sp. exhibited more leaves than the other seedlings (Figure 2C) and showed a trend towards high FB and DB values (Table 3).

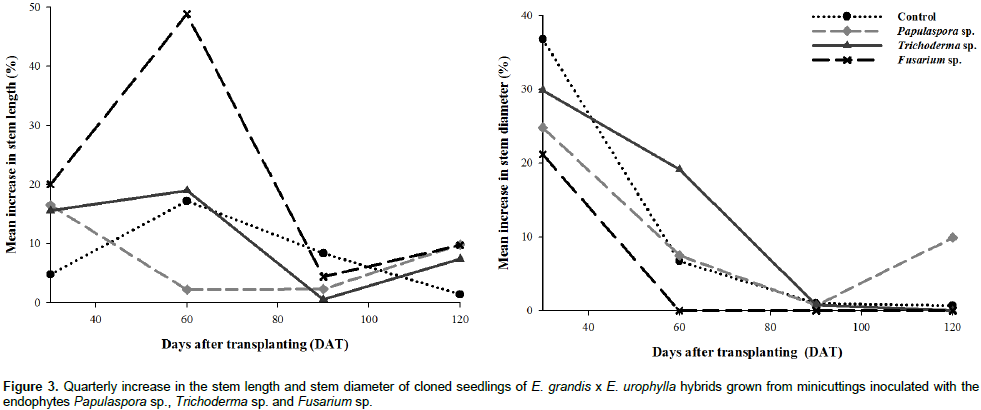
When evaluating the quarterly increase in the SL of cloned of E. grandis x E. urophylla seedlings, the highest percentages were obtained 60 days after transplanting (DAT) in association with the endophytes Fusarium sp. and Papulaspora sp. (Figure 3). Therefore, these fungi (especially Fusarium sp.) were effective in stimulating the cloned seedlings in the early stage of their growth. There have been few studies associating the genus Papulaspora with growth promotion, and the endophytes of this genus are best known for the biosynthesis of enzymes (Tuppad and Shishupala, 2014) and other metabolites with antimicrobial (Ramos et al., 2010) and/or cytotoxic (Gallo et al., 2014) activity. With respect to SD, the largest increases were also obtained in the early stages, where at 60 DAT, the plants inoculated with the fungus Trichoderma sp. exhibited a considerable increase in SD growth; however, in the final assessment at 120 DAT, the plants inoculated with Papulaspora sp. exhibited a greater increase in SD development than the plants inoculated with the other fungi (Figure 3).
DISCUSSION
Endophytic microorganisms are currently being evaluated as growth promoters for use in the production of seedlings of various species. It has been suggested that endophytic fungi can act symbiotically with plants, increasing the absorption of nutrients as well as the response to pathogens and stress conditions, which translates into gains in growth (Mandyam and Jumpponen, 2014). In this study, a linear temporal increase was observed for SL in all treatments. In contrast, for the variable NL, the control plants and those subjected to the inoculation treatments exhibited quadratic behavior. The same pattern was detected in the control plants and those inoculated with Fusarium sp. with respect to SD. This observation may be explained by a reduction in the values of these variables that occurred at the final evaluation times, possibly related to exhaustion of the nutritional content of the available substrate, or even an absence of symbiotic microflora in the autoclaved substrate, specifically in the case of the control plants for the latter scenario. Symbionts mediate the transfer of nutrients from the soil to plants (Barretti et al., 2008; Behie and Bidochka, 2014), and the presence of endophytic fungi has been linked to mechanisms such as the mineralization of available organic matter (Van Hecke et al., 2005), alteration of the chemical and biological properties of the soil and changes in hydraulic characteristics and aggregate stability (Hosseini et al., 2015). Filamentous fungi, such as those used in this work, release enzymes through their hyphae that interact with the organic matter in the substrate (Chigineva et al., 2011) and transform nutrients, such as nitrogen, into forms that are assimilable by plants (Chen et al., 2013). It is possible that endophytic fungi such as mycorrhizae also alter the permeability of roots, facilitating hydraulic conductivity and the absorption of nutrients, which are fundamental processes for the promotion of growth (Ruiz-Lozano and Azcón, 1995).
The cloned seedlings inoculated with Fusarium sp. produced more leaves than the control plants or those inoculated with the other fungal species tested. Endophytes of the genus Fusarium have attracted the interest of a large number of researchers, who have reported their ability to synthesize bioactive secondary metabolites, including exopolysaccharides (Mahapatra and Banerjee, 2012), naphthoquinones (Kornsakulkarn et al., 2011) and cytotoxic alkaloids (Musavi et al., 2015; Venugopalan and Srivastava, 2015). When the potential of this symbiont as a growth promoter was evaluated, its action was found to result in the highest FB and DB values in the cloned E. grandis x E. urophylla seedlings (Table 3). In Annona squamosa, a Fusarium sp. strain endophytic to Annona spp. was also described as a growth promoter due to increase in the DB of the shoots of seedlings cultivated in a greenhouse (de Oliveira Silva et al., 2006). Similar results were observed in seedlings of Passiflora edulis f. flavicarpa Deg, in which this fungus stood out among other endophytes based on promoting increases in shoot FB (Luz et al., 2006). According to Magalhães et al. (2003), the dry matter production rate is strongly affected by leaf area, which is influenced by factors such as environment and management. The amount of biomass produced by the plant can therefore be defined by a simple physiological relationship, based on the amount of radiation intercepted and the efficiency of its conversion into dry matter (Charles-Edwards, 1982). Therefore, the increase in the shoot dry biomass of cloned E. grandis x E. urophylla seedlings may indicate an improvement in the quality of seedlings, which would allow a reduction in the time spent maintaining them in nurseries.
Endophytic fungi have only recently been recognized for their importance in improving the overall fitness of host plants. Despite the prevalence and diversity of plant-endophytic fungus associations, studies have sought to document the impact of using these fungi on plants of agronomic or medical interest; however, such research is still in its incipient stages when compared with the number of studies on growth promotion conducted with bacteria, especially rhizobacteria, such as Rhizobium, Pseudomonas, Bacillus, Azotobacter and Azospirillum (Ahemad and Kibret, 2014; Bashan et al., 2014; Dutta and Khurana, 2015; Egamberdieva and Lugtenberg, 2014; Sharma et al., 2015). Studies using endophytic fungi as growth promoters have thus far been restricted to investigation of their antagonist activity against pathogens (Aktar et al., 2014; Parmar et al., 2015a) and have been much more restrictive than studies conducted with mycorrhizae and non-root rhizosphere fungi. However, new endophytic fungi are frequently isolated and identified as possible inoculants, such as Penicillium funiculosum, Sordariomycetes sp. and Fusarium spp., which were shown to stimulate growth in Glycine max L. (Khan et al., 2011), Oryza sativa L. (Li et al., 2012) and Hordeum vulgare (Maciá-Vicente et al., 2009), respectively.
Fungal strains of the genus Fusarium can be found in symbiotic associations with the internal tissues of most plants (Demers et al., 2015; Singh et al., 2015). However, these strains can also act as pathogenic fungi (Sobowale et al., 2005), triggering Fusarium wilt, or fusariosis, which is characterized by xylem hypertrophy (Pinto et al., 2010) and wilting followed by death of the affected plants (Costa et al., 2010). The high pathogenicity of Fusarium strains in some plants of agronomic interest (Gásperi et al., 2003; Nascimento et al., 2014), together with the great economic losses triggered by fungi of this type in stored fruits (Dantas et al., 2003) or grains (Barros et al., 2005; Ramos et al., 2014), has limited the number of studies attempting to use Fusarium strains as species that promote plant growth. Based on the absence of symptoms caused by infestation with pathogenic Fusarium strains as well as the growth response observed in the plants evaluated in this study, it can be concluded that the tested Fusarium strain is, in accordance with strict criteria, a plant growth-promotingendophytic strain. Many strains of the genus Fusarium and other endophytic fungi can produce phytohormones (Nassar et al., 2005) that directly influence plant development. It is well established that some Fusarium strains are capable of producing the plant hormone gibberellin (GA) (Troncoso et al., 2010; Tudzynski, 2005). Working with strains of Fusarium proliferatum that are symbiotic with orchids, Tsavkelova et al. (2008) identified an isolate that is a significant producer of GAs, and strains symbiotic with Physalis alkekengi have been shown to synthesize a wide range of GAs, including GA3 (Rim et al., 2005). GAs are ubiquitous substances that elicit several important metabolic functions during plant growth (Bomke et al., 2008), which could potentially represent the means by which Fusarium sp. stimulated the cloned E. grandis x E. urophylla seedlings, promoting their growth.
The plants inoculated with Trichoderma sp. exhibited the greatest mean SL at the end of the evaluation period. In E. urograndis, Paz et al. (2012) tested the EUCB 10 strain of the endophytic bacterium Bacillus sp., which was also shown to be efficient in promoting height gains in the evaluated plants, in addition to increasing the dry mass of the roots and shoots. With respect to the increases in stem length and diameter, the seedlings subjected to the isolate tested in the present study responded positively only in the early stages of development (60 DAT). Many studies have associated Trichoderma sp. with growth promotion, though this effect is almost always related to the ability of these fungi to act as an antagonist of plant pathogens (Druzhinima et al., 2011; Kumar et al., 2015; Parmar et al., 2015b), which was not the case in the present study. However, strains of Trichoderma used in biocontrol may stabilize in the rhizosphere, subsequently stimulating plant growth and eliciting plant defense reactions against pathogens (Harman et al., 2011). Trichoderma spp. have been shown to induce growth and control Sclerotinia sclerotiorum in seedlings of the bean plant cv. Carioca grown in nurseries (Aguiar et al., 2012). However, in seedlings of Eucalyptus benthamii, it was indicated that this fungus shows antagonistic potential for the biological control of Botrytis cinerea (Sbravatti Júnior et al., 2013). Research involving species of this genus as well as other fungi that are considered to be potential plant growth promoters is still in the exploratory stages, and studies associating endophytic microorganisms with Eucalyptus species have been conducted with the sole intention of isolation and characterization (Kaewkla and Franco, 2011; Taylor et al., 2009). Therefore, a greater number of studies will be required to improve the understanding and application of these microorganisms in agriculture on a large scale as well as in other areas that rely on biotechnological advances
CONCLUSIONS
From the study, it can be concluded that: 1. plants inoculated with endophytic fungi exhibited better stem growth than that of control plants; 2. cloned E. grandis x E. urophylla seedlings inoculated with Fusarium sp. presented a greater number of leaves as well as higher FB and DB values when compared with the other plants; 3. endophytic fungi were effective in stimulating the growth of cloned E. grandis x E. urophylla seedlings in the early development stage (60 DAT).
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors wish to thank the Research Foundation of the State of Goiás (Fundação de Amparo à Pesquisa do Estado de Goiás - FAPEG) for the Ph.D. scholarship and the National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq).
REFERENCES
|
ABRAF (Associação Brasileira de Produtores de Florestas Plantadas) (2013). Anuário estatístico da ABRAF 2013. Ano base 2012, Brasília: ABRAF. 148 p. |
|
|
Aguiar AR, Machado DFM, Paranhos JT, Silva ACF (2012). Seleção de isolados de Trichoderma spp. na promoção de crescimento de mudas do feijoeiro cv. carioca e controle de Sclerotinia sclerotiorum. Ciênc. Nat. 34(2):47-58. |
|
|
Ahemad M, Kibret M (2014). Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud. Univ. Sci. 26:1-20. |
|
|
Aktar MT, Hossain KS, Bashar MA. (2014. Antagonistic potential of rhizosphere fungi against leaf spot and fruit rot pathogens of brinjal. Bangladesh J. Bot. 43(2):213-217. |
|
|
Alfenas AC, Zauza EAV, Mafia RG, Assis TF (2009). Clonagem e doenças do eucalipto, Viçosa: Universidade Federal de Viçosa. 500 p. |
|
|
Anzuay MS, Frola O, Angelini JG, Luduena LM, Fabra A, Taurian T (2013). Genetic diversity of phosphate-solubilizing peanut (Arachis hypogaea L.) associated bacteria and mechanisms involved in this ability. Symbiosis 60:143-154. |
|
|
Barretti PB, Souza RM, Pozza AAA, Pozza EA, Carvalho JG, Souza JT (2008). Aumento da eficiência nutricional de tomateiros inoculados com bactérias endofíticas promotoras de crescimento. R. Bras. Ci. Solo 32:1541-1548. |
|
|
Barros RG, Barrigossi JAF, Costa JLS (2005) Efeito do armazenamento na compatibilidade de fungicidas e inseticidas, associados ou não a um polímero no tratamento de sementes de feijão. Bragantia 64(3):459-465. |
|
|
Bashan Y, De-Bashan LE, Prabhu SR, Hernandez JP (2014). Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspectives (1998–2013). Plant Soil 378(1-2):1-33. |
|
|
Behie SW, Bidochka MJ (2014) Nutrient transfer in plant–fungal symbioses. Trends Plant Sci. 19(11):734-740. |
|
|
Bomke C, Rojas MC, Gong F, Hedden P, Tudzynski B (2008). Isolation and characterization of the gibberellin biosynthetic gene cluster in Sphaceloma manihoticola. Appl. Environ. Microbiol. 74:5325-5339. |
|
|
Castro PRC, Cato SC, Vieira EL (2001) Aplicação de reguladores vegetais na agricultura tropical, Guaíba: Livraria e Editora Agropecuária.132 p. |
|
|
Chang Y-C, Chang Y-C, Baker R, Kleifeld O, Chet I (1986). Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 70:145-148. |
|
|
Charles-Edwards DA (1982). Physiological determinants of crop growth, London: Academic Press. 161 p. |
|
|
Chen Y, Ren C-G, Yang B, Peng Y, Dai C-C (2013). Priming effects of the endophytic fungus Phomopsis liquidambari on soil mineral N transformations. Microb. Ecol. 65:161-170. |
|
|
Chigineva NI, Aleksandrova AV, Marhan S, Kandeler E, Tiunov AV (2011). The importance of mycelial connection at the soil–litter interface for nutrient translocation, enzyme activity and litter decomposition. Appl. Soil Ecol. 51:35-41. |
|
|
Contreras-Cornejo HA, Macías-Rodriguez L, Beltrán-Pe-a E, Herrera-Estrella A, López-Bucio J (2011). Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungi Botrytis cinerea. Plant Signal. Behav. 6(10):1554-1563. |
|
|
Costa MD, Lovato PE, Sete PB (2010). Micorrização e indução de quitinases e β-1,3-glucanases e resistência à fusariose em porta-enxerto de videira. Pesq. Agropec. Bras. 45(4):376-383. |
|
|
Dantas SAF, Oliveira SMA, Michereff SJ, Nascimento, LC, Gurgel LMS, Pessoa WRLS (2003). Doenças fúngicas pós-Colheita em mamões e laranjas comercializados na central de abastecimento do Recife. Fitopatol. Bras. 28(5):528-533. |
|
|
de Oliveira Silva RL, Luz JS, da Silveira EB, Cavalcante UM (2006). Fungos endofíticos em Annona spp.: isolamento, caracterização enzimática e promoção do crescimento em mudas de pinha (Annona squamosa L.). Acta Bot. Bras. 20(3):649-655. |
|
|
De Santiago A, Quintero JM, Avilés M, Delgado A (2009). Effect of Trichoderma asperellum strain T34 on iron nutrition in white lupin. Soil Biol. Biochem. 41:2453-2459. |
|
|
Demers JE, Gugino BK, Jiménez-Gasco MM (2015). Highly diverse endophytic and soil Fusarium oxysporum populations associated with field-grown tomato plants. Appl. Environ. Microbiol. 81:81-90. |
|
|
Dias PC, Pereira MSF, Magumikasuya MC, Paiva HN, Oliveira LS, Xavier A (2012). Micorriza arbuscular e rizóbios no enraizamento e nutrição de mudas de angico-vermelho. Rev. Árvore 36(6):1027-1037. |
|
|
Díaz K, Valiente C, Martínez M, Castillo M (2009). Sanfuentes E Root-promoting rizobacteria in Eucalyptus globules cuttings. World J. Microbiol. Biotechnol. 25:867-873. |
|
|
Dilaver M, Seyedi N, Bilir N (2015). Seedling Quality and Morphology in Seed Sources and Seedling Type of Brutian Pine (Pinus brutia Ten.). World J. Agric. Res. 3(2):83-85. |
|
|
Druzhinima IS, Seidl-Seiboth V, Herrera-Estrella A, Horwitz BA, Kenerley CM, Monte E, Mukherjee PK, Zeilinger S, Grigoriev IV, Kubicek C (2011). Trichoderma: the genomics of opportiunistic success. Nat. Rev. Microbiol. 9:749-759. |
|
|
Dutta S, Paul Khurana SM (2015). Plant growth-promoting rhizobacteria for alleviating abiotic stresses in medicinal plants. In. PGPR and Medicinal Plants Soil Biology. Springer International Publishing. 42:167-200. |
|
|
Egamberdieva D, Lugtenberg B (2014). Use of plant growth-promoting rhizobacteria to alleviate salinity stress in plants. In. Use of Microbes for the Alleviation of Soil Stresses. Springer New York 1: 73-96. |
|
|
Gallo MBC, Falso MJS, Balem F, Menezes D, Rocha N, Balachandran R, Sturgeon TS, Pupo MT, Day BW (2014). The anti-promyelocytic leukemia mode of action of two endophytic secondary metabolites unveiled by a proteomic approach. Planta Med. 80:473-481. |
|
|
Gama-Rodrigues EF, Barros NF, Gama-Rodrigues AC, Santos GA (2005). Nitrogênio, carbono e atividade da biomassa microbiana do solo em plantações de eucalipto. Rev. Bras. Cienc. Solo 29(6):893-901. |
|
|
Gásperi AC, Prestes AM, Costamilan LM (2003) Reação de cultivares de soja à podridão vermelha da raiz causada por Fusarium solani f. sp. glycines. Fitopatol. Bras. 28(5):544-547. |
|
|
Gonçalves JLM, Alvares CA, Behling M, Alves JM, Pizzi GT, Angeli A (2014). Produtividade de plantações de eucalipto manejadas nos sistemas de alto fuste e talhadia, em função de fatores edafoclimáticos. Sci. For. 42(103):411-419. |
|
|
Harman GE (2011). Multifunctional fungal plant symbionts: new tools to enhance plant growth and productivity. New Phytol. 189:647-649. |
|
|
Hosseini F, Mosaddeghi MR, Hajabbasi MA, Sabzalian MR (2015). Aboveground fungal endophyte infection in tall fescue alters rhizosphere chemical, biological, and hydraulic properties in texture-dependent ways. Plant Soil 388:351-366. |
|
|
Kaewkla O, Franco CMM (2011). Flindersiella endophytica gen. nov., sp nov., an endophytic actinobacterium isolated from the root of Grey Box, an endemic eucalyptus tree. Int. J. Syst. Evol. Microbiol. 61:2135-2140. |
|
|
Khan AL, Hamayun M, Kim YH, Kang SM, Lee IJ (2011). Ameliorative symbiosis of endophyte (Penicillium funiculosum LHL06) under salt stress elevated plant growth of Glycine max L. Plant Physiol. Biochem. 49:852-861. |
|
|
Kloepper JW, Hume DJ, Scher FM, Singleton C, Tipping B, Laliberte M, Frauley K, Kutchaw T, Simonson C, Lifshitz R, Zaleska I, Lee L (1988). Growth-promoting rhizobacteria on canola (rapeseed). Plant Dis. 72:42-46. |
|
|
Kornsakulkarn J, Dolsophon K, Boonyuen N, Boonruangprapa T, Rachtawee P, Prabpai S, Kongsaeree P, Thongpanchang C (2011). Dihydronaphthalenones from endophytic fungus Fusarium sp. BCC14842. Tetrahedron 67(39):7540-7547. |
|
|
Kumar V, Shahid M, Srivastava M, Singh A, Pandey S, Maurya MK (2015). Screening of Trichoderma species for virulence efficacy on seven most predominant phytopathogens. Microbiol. Res. 9(11):793-799. |
|
|
Kuss AV, Kuss VV, Lovato T, Flores ML (2007) Fixação de nitrogênio e produção de ácido indolacético in vitro por bactérias diazotróficas endofíticas. Pesq. Agropec. Bras. 42(10):1459-1465. |
|
|
Lammel DR, Cruz LM, Carrer H, Cardoso EJBN (2013). Diversity and symbiotic effectiveness of beta-rhizobia isolated from sub-tropical legumes of a Brazilian Araucaria Forest. World J. Microbiol. Biotechnol. 29:2335-2342. |
|
|
Lemanceau P, Expert D, Gaymand F, Bakker PAHM, Briat JF (2009). Role of iron in plant–microbe interactions. Adv. Bot. Res. 51:491-549. |
|
|
Li X, Bu N, Li Y, Ma L, Xin S, Zhang L (2012). Growth, photosynthesis and antioxidant responses of endophyte infected and non-infected rice under lead stress conditions. J. Hazard. Mater. 213:55-61. |
|
|
Lombard L, Zhou XD, Crous PW, Wingfield BD, Wingfield MJ (2010). Calonectria species associated with cutting rot of Eucalyptus. Persoonia 24:1-11. |
|
|
Luz JS, Silva RLO, Silveira EB, Cavalcanti UMT (2006). Atividade enzimática de fungos endofíticos e efeito na promoção do crescimento de mudas de maracujazeiro-amarelo. Caatinga 19(2):128-134. |
|
|
Maciá-Vicente JG, Rosso LC, Ciancio A, Jansson H-B, Lopez-Llorca LV (2009). Colonisation of barley roots by endophytic Fusarium equiseti and Pochonia chlamydosporia: Effects on plant growth and disease. Ann. Appl. Biol. 155(3):391-401. |
|
|
Mafia RG, Ferreira EM, Binoti DHB, Mafia MV, Mounteer AH (2009). Root colonization and interaction among growth promoting rhizobacteria isolates and Eucalyptus species. Rev. Árvore 33(1):1-9. |
|
|
Magalhães PC, Durães FOM, Rodrigues JAS (2003). Fisiologia da Planta de Sorgo, Sete Lagoas: EMBRAPA/ CNPMS. P 4. |
|
|
Mahapatra S, Banerjee D (2012). Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr. Polym. 90(1):683-689. |
|
|
Mandyam K, Jumpponen A (2014). Unraveling the dark septate endophyte functions: insights from the Arabidopsis model. In. Advances in Endophytic Research 1:115-141. |
|
|
Mehnaz S (2014). Azospirillum: A biofertilizer for every crop. In. Plant Microbes Symbiosis: Applied Facets, Springer India. 1:297-314. |
|
|
Melo LC, Oliveira CV, Manfredi C, Baldani VLD, Ferreira JS (2012). Efeito de bactérias na promoção do enraizamento em clone de eucalipto. Enciclopédia Biosfera 8(15):736-747. |
|
|
Miethke M, Marahiel MA (2007). Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 71:413-451. |
|
|
Musavi SF, Dhavale A, Balakrishnan RM (2015). Optimization and kinetic modeling of cell-associated camptothecin production from an endophytic Fusarium oxysporum NFX06. Prep. Biochem. Biotechnol. 45(2):158-172. |
|
|
Nascimento DM, Vieira FEC, Batista TB, Koyanagui M, Bardiviesso EM, Vieira GHC (2014). Controle in vitro do Fusarium sp. causador da fusariose na soja. Cadernos Agroecol. 9(4):1-11. |
|
|
Nassar AH, Ei-Tarabily KA, Sivasithamparam K (2005). Promotion of plant growth by an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (Zea mays L.) roots. Biol. Fertil. Soils 42(2):97-108. |
|
|
Parmar HJ, Bodar NP, Lakhani HN, Patel SV, Umrania VV, Hassan MM (2015b). Production of lytic enzymes by Trichoderma strains during in vitro antagonism with Sclerotium rolfsii, the causal agent of stem rot of groundnut. Afr. J. Microbiol. 9(6):365-372. |
|
|
Parmar HJ, Hassan MM, Bodar NP, Umrania VV, Patel SV, Lakhani HN (2015a). In vitro antagonism between phytopathogenic fungi Sclerotium rolfsii and Trichoderma strains. Int. J. Appl. Sci. Biotechnol. 3(1):16-19. |
|
|
Paz ICC, Santin RCM, Grimarães AM, Rosa OPP, Dias ACF, Quecine MC, Azevedo JL, Matsumura ATS (2012). Eucalyptus growth promotion by endophytic Bacillus spp. Genet. Mol. Res. 11(4):3711-3720. |
|
|
Pii Y, Mimmo T, Tomasi N, Terzano R, Cesco S, Crecchio C (2015). Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. Biol. Fertil. Soils 51(4): 403-415. |
|
|
Pinto Z, Bettiol W, Morandi MAB (2010). Efeito de casca de camarão, hidrolisado de peixe e quitosana no controle da murcha de Fusarium oxysporum f. sp. chrysanthemi em crisântemo. Trop. Plant Pathol. 35(1):16-23. |
|
|
R Core Team (2016). R: A language and environmental for statistical computing. R Foundation for Statistical Computing, Viena, Austria. URL: http://www.R-project.org/. |
|
|
Rajkumar M, Ae N, Prassad MNV, Freitas H (2010). Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 28(3):142-149. |
|
|
Ramos DP, Barbosa RM, Vieira BGTL, Panizzi RC, Vieira RD (2014). Infecção por Fusarium graminearum e Fusarium verticillioides em sementes de milho. Pesq. Agropec. Trop. 44(1):24-31. |
|
|
Ramos HP, Braun GH, Pupo MT, Said S (2010). Antimicrobial activity from endophytic fungi Arthrinium state of Apiospora montagnei Sacc. and Papulaspora immersa. Braz. Arch. Biol. Technol. 53(3):629-632. |
|
|
Richardson AE, Barea J-M, Mcneill AM, Prigent-Combaret C (2009). Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321:305-339. |
|
|
Rim SO, Lee JH, Choi WY, Hwang SK, Suh SJ, Lee IJ, Rhee IK, Kim JG (2005). Fusarium proliferatum KGL0401 as a new gibberellin-producing fungus. J. Microbiol. Biotechnol. 15:809-814. |
|
|
Ruiz-Lozano JM, Azcón R (1995). Hyphal contribution to water uptake in mycorrhizal plants as affected by the fungal species and water status. Physiol. Plant. 95(3):472-478. |
|
|
Sadeghi A, Karimi E, Dahaji PA, Javid MG, Dalvand Y, Askari H (2012). Plant growth promoting activity of an auxin and siderophore producing isolate of Streptomyces under saline soil conditions. World J. Microbiol. Biotechnol. 28:1503-1509. |
|
|
Sbravatti Junior JÁ, Auer CG, Pimentel IC, Santos AF, Schultz B (2013). Seleção in vitro de fungos endofíticos para o controle biológico de Botrytis cinerea em Eucalyptus benthamii. Floresta 43(1):145-152. |
|
|
Sharma A, Chaubey OP (2015). Biotechnological approach to enhance the growth and biomass of Tectona grandis Linn. F. (Teak) seedlings. J. BioSci. Biotechnol. 7(1):19-28. |
|
|
Sharma S, Singh V, Kumar V, Devi S, Shukla KP, Tiwari A, Singh J, Bisht S (2015). Plant growth-promoting rhizobacteria (PGPR): emergence and future facets in medicinal plants. In. Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants Soil Biology. Springer International Publishing 42:109-131. |
|
|
Singh AK, Rathod V, Singh D, Ninganagouda S, Kulkarni P, Mathew J, Haq MU (2015). Bioactive silver nanoparticles from endophytic fungus Fusarium sp. isolated from an ethanomedicinal plant Withania somnifera (Ashwagandha) and its antibacterial activity. J. Nanomater. Biostruct. 5(1):15-19. |
|
|
Sobowale AA, Cardwell KF, Odebode AC, Bandyopadhyay R, Jonathan SG (2005). Growth inhibition of Fusarium verticillioides (Sacc.) Nirenberg by isolates of Trichoderma pseudokoningii strains from maize plant parts and its rhizosphere. J. Plant Prot. Res. 45(4):249-265. |
|
|
Taghavi S, Garafola C, Monchy S, Newman L, Hoffman A, Weyens N, Barac T, Vangronsveld J, Van Der Lelie DD (2009). Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of popular trees. Appl. Environ. Microbiol. 75:748-757. |
|
|
Taylor K, Barber PA, Hardy GES, Burgess TI (2009). Botryosphaeriaceae from tuart (Eucalyptus gomphocephala) woodland, including descriptions of four new species. Mycol. Res. 113:337-353. |
|
|
Teixeira DA, Alfenas AC, Mafia RG, Ferreira EM, Siqueira L, Maffia LA, Mounteer AH (2007). Rhizobacterial promotion of eucalypt rooting and growth. Braz. J. Microbiol. 38:118-123. |
|
|
Troncoso C, González X, Bömke C, Tudzynski B, Gong F, Hedden P, Rojas MC (2010). Gibberellin biosynthesis and gibberellin oxidase activities in Fusarium sacchari, Fusarium konzum and Fusarium subglutinans strains. Phytochemistry 71:1322-1331. |
|
|
Tsavkelova EA, Bomke C, Netrusov AI, Weiner J, Tudzynski B (2008). Production of gibberellic acids by an orchid-associated Fusarium proliferatum strain. Fungal Genet. Biol. 45:1393-1403. |
|
|
Tudzynski B (2005). Gibberellin biosynthesis in fungi: genes, enzymes, evolution, and impact on biotechnology. Appl. Microbiol. Biotechnol. 66:597-611. |
|
|
Tuppad DS, Shishupala S (2014). Evaluation of endophytic fungi from Butea monosperma for antimicrobial and enzyme activity. J. Med. Plants Stud. 2(4):38-45. |
|
|
Van Hecke MM, Treonis AM, Kaufman JR (2005). How does the fungal endophyte Neotyphodium coenophialum affect tall fescue (Festuca arundinacea) rhizodeposition and soil microorganisms? Plant Soil 275:101-109. |
|
|
Venugopalan A, Srivastava S (2015). Enhanced camptothecin production by ethanol addition in the suspension culture of the endophyte, Fusarium solani. Bioresour. Technol. 188:251-257. |
|
|
Vitorazi Filho JÁ, Lima KB, Freitas MSM, Martins MA, Olivares FL (2012). Crescimento de mudas de maracujazeiro-doce inoculadas com fungos micorrízicos arbusculares e bactérias diazotróficas sob diferentes doses de fósforo. Rev. Bras. Frutic. 34(2):442-450. |
|
|
Vitorino LC, Silva FG, Soares MA, Souchie EL, Costa AC, Lima WC (2012). Solubilization of calcium and iron phosphate and in vitro production of Indoleacetic acid by Endophytic isolates of Hyptis marrubioides Epling (Lamiaceae). Int. Res. J. Biotechnol. 3:47-54. |
|
|
Yazici N, Babalik AA (2011). Determination of suitable irrigation interval for Anatolian Black pine (Pinus nigra Arn. subsp. Pallasiana (Lamb.) Holmboe.) seedlings. J. Bartin For. Fac. 13(19): 100-106. |
|
|
Yazici N, Babalik AA (2016). Effect of Irrigation Density on Seedling Morphology in Taurus Cedar (Cedrus libani A. Rich.). Int. J. Sci.: Basic Appl. Res. 27:211-218. |
|
|
Yazici N, Ozhan S, Babalik AA (2011). Determination of water consumption of crimean juniper (Juniperus excelsa Bieb.) seedlings and its relation with meteorological. SDU Fac. For. J. 12:84-88. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


