Full Length Research Paper

ABSTRACT
INTRODUCTION
In Ethiopia, among the oilseeds, Ethiopian mustard stands third next to niger seed and linseed in total production and area coverage (CSA, 2009). Its area and production are estimated to be 34,580 hectares and 47,721 tons, respectively, at private peasants holdings level, with an average productivity of 0.68 tones/ha (CSA, 2009). This
low production is attributed to a number of production constraints such as lack of high yielding, early maturing varieties, high erucic acid (C22:1) content in seed oil and high glucosinolate content in the meal (EARO, 2000).
Ethiopian mustard, Brassica carinata A. Braun (BBCC, 2n=4x=34) is believed to have originated in the Ethiopia plateau through natural hybridization of Brassica nigra (BB, 2n=2x=16) with Brassica oleracea (CC, 2n=2x=18), followed by the chromosome doubling of the hybrids, and has several agronomical important traits such as non-dehiscent siliques, much more developed and aggressive root system than B. napus. It is resistance to a wide range of diseases, pests and tolerant to many abiotic stresses (Katiyar et al., 1986; Malik, 1990), which makes it a suitable candidate as a food security crop in Ethiopia.
Plant transformation systems have been developed for many economically important species of the genus Brassica such as B. napus (Moloney et al., 1989), B. oleracea (Deblock et al., 1989), Brassica juncea (Barfield and Pua, 1991), Brassica rapa (Radke et al., 1992), Brassica nigra (Gupt et al., 1993) and B. carinata (Babic et al., 1998) which facilitates obtaining transgenic plants with modified agronomic traits. Many genetic improvements, such as herbicide tolerance, improved oil quality and production of pharmacological and industrial products, have been achieved by genetic transformation in the Brassica species. For example, in B. napus seeds, high levels of gamma-linoleic acid were obtained by the introduction of δ12-desaturase genes from the fungus Mortierella alpine (Liu et al., 2001). In addition B. carinata used to produce biodiesel, and their erucic acid contents are used as chemical additives in plastic, tannery and cosmetic industries (Bozzini et al., 2007), and also as luminant or lubricant in soap making (Greville, 2005).
Genetic improvement of Brassica spp. has been mainly achieved by conventional breeding methods. Recently, genetic engineering opened a new avenue for plant improvement (Hansen and Wright, 1999). Regeneration in Brassica is highly genotyped, age dependent and has been reported in several species (Guo et al., 2005). B. napus cultivar GSL-1 showed better regeneration efficiency than Westar (a standard cultivar for transformation) in a study by Phogat et al. (2000).
In vitro regeneration is one of a key factor in developing an efficient transformation method in plants. In Brassica spp. in vitro regeneration is highly genotype-dependent for B. napus (Ono et al., 1994; Phogat et al., 2000), B. rapa (Zhang et al., 1998) and B. oleracea (Sparrow et al., 2006). In addition, Dietert et al. (1982) compared 6 species of the genus Brassica for callus growth and plant regeneration and reported a high influence of the genotype in the in vitro culture. However, the available information on the genotype and explant variability for in vitro culture and shoot regeneration in B. carinata is limited to a small number of genotypes, thus a limiting factor for the application of genetic engineering to a wide number of genotypes. For that reason, it is important to identify highly regenerant genotypes that can be used in transformation via Agrobacterium tumefaciens.
However, no work has been done on the effects of various growth regulators on the callus formation from the hypocotyl and cotyledon explants of Yellow Dodola and Holleta-1 B. carinata genotypes. But there are a number of studies carried out in the direct regeneration to increase the regeneration frequency of Brassica spp. and remarkable progress has been achieved. Hence, the main objective of this study was to establish an in vitro regeneration and propagation protocol for the two rapseed (B. carinata) cultivars, Yellow Dodola and Holeta-1 using hypocotyl and cotyledon explants.
MATERIALS AND METHODS
The experiment was conducted in Holleta Agricultural Biotechnology Laboratory. Two varieties of B. carinata namely Holeta-1 and Yellow Dodola obtained from Highland Oil Crops Research Department of Holleta Agricultural Research Centre, were used for this study.
Surface sterilization and germination
Seeds of B.carinata genotypes Holeta-1 and Yellow Dodola were first washed by soaking in tap water for 5 min to remove dust and dirt. The seeds were then transferred to 70% ethanol for 1 min followed by 1% sodium hypochlorite (NaOCl) for 20 min. The surface sterilized seeds were then rinsed with sterile distilled water for 2 to 3 times. The pH of the culture medium was adjusted to 5.8 using 1.0 M HCl or 0.5 M NaOH before autoclaving and autoclaved at 15 psi at 121°C for 15 min. The seeds were plated for germination on the previously prepared hormone free full strength MS media containing 2% sucrose with 0.6% agar under sterile condition and were incu-bated at 25±2°C in a 16/8 h day/night photoperiodic regime under cool white fluorescent lights (2700 -Lux) µmol quanta m-2 s-1 for 5–7 days.
Cotyledon leaves and hypocotyls were excised from 5-7 days old seedlings under sterile condition and were cut into 0.5-1 cm pieces of explants excluding the meristematic axillary bud. Five explants were then placed horizontally in each magenta box with callus induction MS medium supplemented with different concentrations of 2,4-dichlorophenoxy acetic acid (2,4-D), naphthalene acetic acid (NAA) and 6-benzyl amino purine (BAP).
Callus induction medium
Explants were cultured on the basal MS-medium supplemented with three hormones 2,4-D, BAP and NAA at different concentrations (0.1, 0 .25, 0.5, 1.0 and 1.5 mg/L) to enhance callus initiation. All growth conditions were maintained with 3 replications in complete dark for one month. The explants were then sub-cultured every 4-5 weeks on the freshly prepared MS medium until callus was formed. The percentage of explants (%) that induced calli and their weights (gm) were recorded for each treatment.
Shoot induction medium
After about 30-48 days of callus culture, the calli were transferred aseptically onto a sterile Petri dish and were cut into convenient size by a sterile scalpel or blade, then transferred onto a freshly prepared media supplemented with 0, 1, 2 and 3 mg/L BAP. Each culture box contained 5 pieces of calli and all the treatments were performed with three replications and sealed with parafilm in a growth room under 16 h light at 2700 µmol quanta m−2 s−1 lux light intensity and 8 h dark cycle at 25 ±2°C. The explants were sub-cultured every 2-3 weeks intervals on the freshly prepared medium to obtain shoot regeneration. The parameters recorded were the percentage of explants producing shoots (%), the mean number of shoots produced per explants and shoot height attained (cm).
Root induction medium
Eight weeks later, regenerated shoots were carefully removed from the culture box and cultured in freshly prepared MS medium containing different hormonal supplements like IBA and NAA at different concentrations (0.0, 0.1, 0.3 and 0.5 mg/L) for root initiation. The MS media contained 2% sucrose, 0.6% agar and the pH was 5.8. Parameters recorded were the percentage of explants producing root (%), the mean number of roots produced per explants, root length attained (cm) and the fresh and dry weight of shoots attained (cm) after four weeks of culture. Plantlets with approximate height of 4-7.7 cm, with well-developed roots were transferred to a small acclimatization pot, containing sterilized soil mix with a proportion of 1:2:1 sand, red soil and compost, respectively.
Experimental design and statistical analysis
The experiments were arranged in a complete randomized design (RCD), with three replications and each replication per treatment contained 5 explants. Data were analyzed using analysis of variance (ANOVA) using statistical analysis system (SAS versions) software and the least significant differences among mean values were compared using DMRT at p≤0.05.
RESULTS AND DISCUSSION
The in vitro morphogenetic responses of cultured plants are affected by different components of the culture media and therefore, it is important to evaluate their effects on plant callus induction and regeneration (Gubis et al., 2004). The surface sterilized germinating seeds were used as sources of explants for callus induction. The explants were inoculated on MS medium with variable ranges of 2, 4-D, BAP, NAA.
It was observed that the explants showed an initial swelling followed by callus formation within 2 - 3 weeks of incubation, in treatment without plant growth regulators. Callus proliferation started from cut ends of hypocotyls and cotyledon on MS medium supplemented with different concentrations of 2, 4-D, BAP and NAA and eventually extended all over the explants. Callus initiation began from cut edges after 12 days in cotyledon explants and after 15-20 days in hypocotyl explants.
Callus produced from cotyledon explants grew faster during the first 7-30 days of culture in both varieties. Approximately, after 3 weeks of culture, the explants were almost completely converted into callus. Similar results have been described in other reports. Ullah et al. (2004) reported that on B. napus cv. Rainbow explants, callus proliferation started from the cut ends of the hypocotyls and cotyledon. Al–Naggar et al. (2010) reported different results in B. napus; 2,4-D induced callus production over the entire surface of the cotyledon and hypocotyls.
The type and quantity of callus and callogenesis efficiency depends on the duration of their exposure to hormone and on the type of explants (Bano et al., 2010; Khan and Rashid, 2002). In this experiment, the highest quantity of callus was formed in cotyledons after short induction on MS with 2,4-D. BAP and NAA required more time to produce callus and caused poor formation of compact calli, with many necrotic sites. Abellatef et al. (2008) and Dietert et al. (1982) obtained similar results in cotton and B. napus cv. Evitain on MS media supple-mented with 2,4-D and BAP, respectively. Highly significant difference in percentage of callus formation was observed in between two types of explants, PGRs and the genotypes used (Table 1). These results are in agreement with those reported by Sayed et al. (2010) and Khan et al. (2010).
2,4-D is among the most widely used auxin for in vitro callus induction in a wide range of plant species (Al-Naggar et al., 2010; Khan and Rashid, 2002). The highest percentage of callus (95%) was obtained on cotyledon explants cultured on MS medium supplemented with 2,4-D at 0.5 mg/L with 1.5 g mean weight (Table 1), followed by from the same explants on 1 mg/L 2,4-D (86%). However, for the hypocotyl explant, 80.7% of callus was obtained on MS medium supplemented with 2,4-D at 0.1 mg/L (Table 1), followed by using 0.25 mg/L 2,4-D (63.9%) within the genotype of Yellow-Dodola. In Holeta–1, The highest percentage of callus (88.9%) was obtained on cotyledon explants cultured on MS medium supplemented with 0.5 mg/L, 2,4-D followed by 80% from the same explants by using 1 mg/L of 2,4-D. However, for the hypocotyls explants, 66 and 73% of callus were obtained respectively with the same concentrations of 2,4-D as mentioned in cotyledon (Table 1). In line with the present results, Magdoleen et al. (2010) also reported that the most efficient hormone concentration depends on the genotype used and the type of explants. The cotyledon in particular was found to be the best for callus proliferation in both genotypes.
The Yellow Dodola genotype in particular was found to be the best for callus initiation in both explants types with 0.5 mg/L 2,4-D. It had the highest callus initiation frequency (CIF) and produced calli with more weight than the Holleta-1 within the different concentration levels of 2,4-D. These results are in harmony with those reported by Ali et al. (2007) in B. napus.
In study on the effects of different concentrations of BAP on callus initiation from hypocotyl and cotyledon, the results show that MS medium supplemented with 0.5 mg/L of BAP was the most effective in callus induction for both types of explants and genotype. 0.5 mg/L BAP was more effective in inducing callus from cotyledon explants than hypocotyls. A higher concentration of BAP (1.5 mg/L) inhibited callus proliferation. BAP at low concentrations were more effective for induction of callus. The present study showed that callus initiation frequency (CIF) was 80 and 50% for Yellow Dodola cotyledon and hypocotyls explant, supplemented with concentration of 0.5 mg/L of BAP respectively (Table 1). Whereas, Holetta–1 had CIF of 72 and 44% on cotyledon and hypocotyl explants supplemented with the same concentration of BAP, respectively (Table 1). Among the explants types, the cotyledon explants was more responsive than the hypocotyls.
Comparison of different explants types and lines in terms of callus production revealed a genotypic variation between the types of explants and lines with Holleta-1 being the least responsive while Yellow Dodola was the most productive. This finding correlated with the result of various research reports in B. napus (Datta and Conger, 1999; Fang et al., 2005; Khan et al., 2010; Pathirana and Eason, 2006, 2004; Rao et al., 2006.
Effects of NAA on callus induction
In study on the effects of different concentrations of NAA on callus initiation from hypocotyl and cotyledon, the results show that MS medium supplemented with 1.5 mg/L of NAA was the most effective in callus induction for both types of explants and genotype. 1.5 mg/L NAA was more effective in inducing callus in explants hypocotyls than cotyledon. The present study showed that callus initiation percentage was 66.8 and 52.8% for Yellow Dodola hypocotyls and cotyledon explants, supplemented with concentration of 1.5 mg/L NAA respectively (Table 1) whereas, in Holetta–1, the best callus induction was observed in cotyledonary explants and it was 73 and 69% at the concentration of 1.5 and 2.5 mg/L of NAA in MS medium, respectively. The second best callus was 58% at the concentration of 1.5 mg/L of NAA from the hypocotyls explants (Table 1). For the comparison of explants types, the cotyledon explants was more responsive than the hypocotyls (Table 1). These results are in agreement with those reported by Magdoleen et al. (2010) and Chamandosti et al. (2006). Comparison of different explants types and lines in terms of callus production revealed a genotypic variation between the types of explants and lines with Holleta-1 being the best responsive while Yellow Dodola was the least productive.
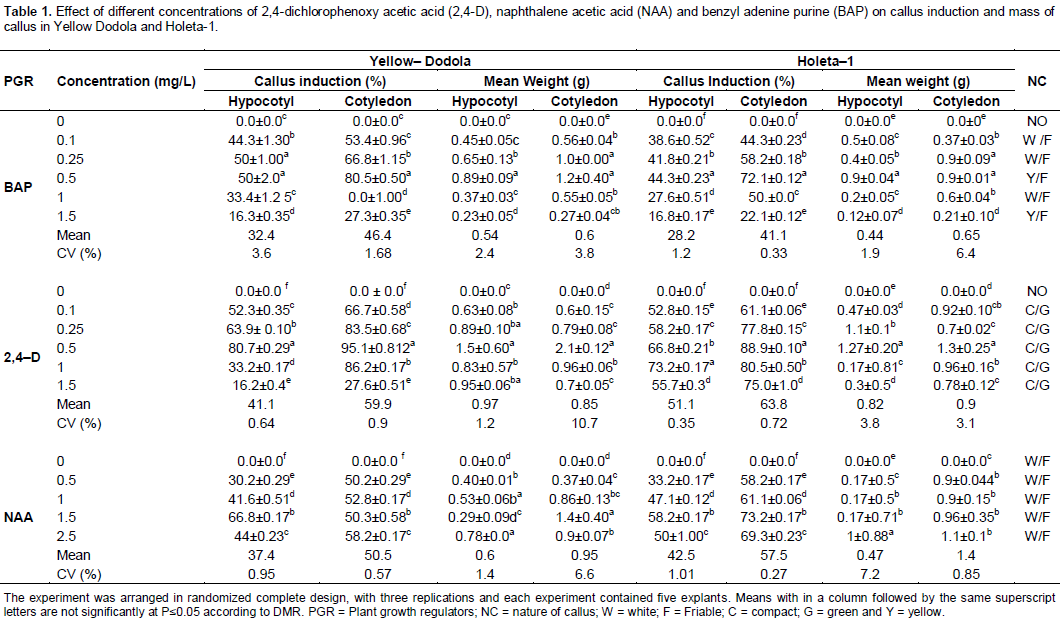
Two morphological types of calli were obtained after 30 days of culture; white friable and compact green. Calli derived from hypocotyls became soft or friable, sticky, yellow-white to cream colored and nodular type, and mostly consisted of large and translucent cells (Figure 1C and D). Whereas calli from cotyledons were nodular, green, organogenetic, compact green calli and non-friable, with small and green cells often developed on media containing 0.5 mg/L 2,4-D and formed round masses (Figure 1A and B). This result is in consistence with the result obtained for sugar beet by Kamal et al. (2007) and Chamandosti et al. (2006) which reported similar findings in B. napus; three morphologically distinct types of calli were induced by using different concen-trations of 2,4-D and BAP.
Shoot proliferation occurred through a callus phase. The development of shoots (Figure 2) from white friable and compact callus (regardless of the explants type) was observed after 3-4 weeks of culture. Shoot development was achieved at all concentrations of BAP except for the treatment without plant growth regulator (Table 2). The percentage of explants forming shoots varied between type of explants (hypocotyl and cotyledon) as well as between the genotypes, Holleta–1 and Yellow Dodola on MS media supplemented with different concen-trations of the BAP. These findings exhibited a varied response (22-90%) of shoot regeneration from both explants of both cultivars. Similar results were obtained by Moghaieb et al. (2006) and Kamal et al. (2007) in B. napus.

The explants developed as tiny patches of pale green callus bearing several hump-like structures within a period of 20-30 days. At the concentration of 0.5 mg/L 2, 4-D subsequently, they differentiated into shoot buds. Best shoot elongation was achieved when regenerating segment of the callus was cut into smaller pieces containing three or four shoots and sub cultured in the basal medium supplemented with 2 mg/L BAP. These results are correlated with those reported by Moghaieb et al. (2006), Zeynali et al. (2010) and Kamal et al. (2007) in B. napus. Results of the effect of BAP on shoot initiation in the two varieties of B. carinata are presented in Table 2 and Figure 2. The result reveals that all varieties respond to all treatments. Morphologically, best shoots were induced on the medium that contained 2 mg/L BAP (Figure 2). Highest percentage of survival rate of cultured explants was obtained at 2 mg/L of BAP. This holds true for both the varieties. Shoot regeneration frequency (SRF) was 90% for hypocotyl explants and 45% for cotyledonary explants of Yellow Dodola at 2 mg/L of BAP. In Holleta–1 there is a significance difference among the concentrations. The highest recorded SRF is 56% from hypocotyls explants and 34% from cotyledonary explants (Table 2) at 2 mg/L of BAP. Based on this result, 2 mg/L BAP concentration was used as an optimum PGRs for shoot initiation. Similar results have been reported by Ravanfar et al. (2009) in B. oleracea.
The same results have been reported by Khan and Rashid (2002) where they have shown that the important factors for shoot regeneration were explant type and genotype. This is also in agreement with Moghaieb et al. (2006), Yang et al. (1991) and Yu et al. (2005). Thus, it is clearly shown that the regeneration depends on explants type and genotype.
Among the four different BAP, 2 mg/L of BAP produced significantly maximum number of shoots for both varieties (Table 2). Yellow Dodola gave maximum mean number of shoots (7.7) followed by Holleta–1 (6) from the hypocotyl explants. The maximum shoot height attained at 2 mg/L of BAP for Yellow Dodola and Holleta–1 were 7.7 and 6.7 cm, respectively from the hypocotyl explants (Table 2). Medium supplemented with 2 mg/L of BAP was the best in many of the parameters recorded. However, there was a significant difference between the cultivars in number of shoots developed and shoots length. Yellow Dodolla was best in both cases (Table 2).
The responses of explants for shoots regeneration were different. Various authors (Munshi et al., 2007; Gubis et al., 2004) reported that hypocotyl explants were superior to cotyledon in shoot regeneration efficiency and shoot length. MS medium supplemented with 2 mg/L BAP was the most effective in adventitious shoot formation and shoot length in both explants and cultivars. This result agreed with those of George et al. (2008) who showed that one of the B. napus cultivars named RK–7 had low shoot regeneration (18%) from cotyledonary explants while the same cultivar had a higher (27%) shoot regeneration from the hypocotyls explants.
Genotypic effects
Genotypic effect is well established in tissue culture response. Different genotypes had different physiological requirement of plant growth regulators for in vitro shoot and root regeneration (Malik et al., 2005).
The response of various genotypes to callus formation varied considerably. Similarly, the effect of hormones on callus formation of the two genotypes was different (Table 1). A differential response in callus induction and survival was noticed when calli were cultured in media supplemented with auxins (2,4-D and NAA) and cytokinin (BAP) at different concentrations. The genotype, Yellow Dodola was found more responsive to callus production than Holleta–1 (Table 1). Such results can be attributed to the genetic differences in the two varieties. Genotypic influence on in vitro morphogenesis in Brassica has been documented previously (Khan and Rashid, 2002; Dietert et al., 1982; Fazekas et al., 1986; Khehra and Mathias, 1992). Such reports indicated that the developmental processes reflected by in vitro response are genetically controlled. Total number of shoots regenerated varied significantly among the two genotypes. During this study, a better consistent response and maximum number of shoots (90%) was produced from Yellow Dodola.
Root organogenesis
Effect of NAA and IBA on root induction
The rooting results reveal that plantlets cultured on rooting media induced roots in all media whether or not supplemented with NAA and IBA. The primary roots were observed after two weeks of culturing on MS medium supplemented by different concentrations of IBA and NAA. However, well developed and mature roots were obtained after a month. Rooting of plantlets was usually attained at half MS medium with low concentrations of auxins and sucrose. Similar result was reported by Khan and Rashid (2002). Among the different auxins, IBA was the best for root induction. The treatment containing 0.3 mg/L IBA and the control (MSO) gave the highest percentage of plantlets producing roots and mean number of roots per plantlet in both genotypes (Table 3). The highest percentage of explant producing roots (98%) and mean number of roots produced per explants (9) was observed on Yellow Dodela supplemented with IBA (0.3. mg/L) and the lowest frequency of response (41%) on Yellow Dodola fortified with NAA (0.5 mg/L). Whereas, Holleta–1 had the highest and lowest root formation at 92 and 38% on media fortified with 0.3 and 0.5 mg/L of IBA and NAA, respectively. NAA supplemented medium resulted in low root induction and shoots in the medium had a tender callus at the base of shoots and produced stumpy and thick roots. These results are in agreement with the findings of Ravanfar et al. (2009). Caboni and Tonalli (2002) reported that IBA is the most effective auxin for root induction in a wide range of plant species. IBA is superior in stability than NAA. The present study also proved that IBA was better than NAA for root induction. Similarly, Salman (2002) as well as Ali et al. (2007) reported a similar result by using 0.3 mg/L of IBA in B. oleracea and B. napus (Westar) respectively where 90% root formation was recorded in the latter.
The second maximum frequency of root formation (91%) of Yellow Dodola was achieved on half strength MS medium without growth regulators along with maximum root length (4.7 cm). It is assumed that root formation on auxin free medium may be due to the availability of higher quantity of endogenous auxin in shootlets raised in vitro. Similar results were reported by Ali et al. (2007) in B. napus. Both treatments showed significant difference with respect to the percentage of shoots forming roots, mean number of roots produced per shoot, root length, fresh and dry weight. In this study, the highest increase in fresh and dry weight was observed in medium supplemented with 0.3 mg/L IBA. There was a significant difference between the root length attained with IBA and NAA treatment. NAA produced fewer roots than IBA. Similar results were obtained by various authors (Moghaieb et al., 2006; Chamandosti et al., 2006; Kamal et al., 2007).
The response of different genotypes to root induction varied considerably. Similarly, the effect of hormones on root formation of the two genotypes was different (Table 3) Variations in percentage and root length among the tested cultivars indicated that the differences in responses were due to the differences between the genotypes. Genotypic variations in in vitro culture system are frequently observed in B. carinata cultivars. During the present study, it was revealed that the varieties varied significantly for their regeneration capacity on the similar medium. This might be due to their genetic differences. Similarly, genotypic influence on in vitro morphogenesis in Brassica spp. has been documented previously (Dietert et al., 1982; Khan et al., 2010). After thorough washing of the well-developed roots with tap water, in vitro raised plantlets were then transferred to the pot containing sterilized soil mixed with a proportion of 1:2:1 sand, red soil and compost, respectively, at green house with 80% humidity, 25°C temperature and 16 h light of about 2700 lux of light intensity and 8 h dark. About 100% plantlets were successfully established.
CONCLUSION
The present study describes an effective indirect regene-ration protocol for in vitro propagation of two genotypes of B. carinata. Cotyledon explants gave higher callus initiation percentage than hypocotyls explants in both genotypes at the concentration of 0.5 mg/L 2, 4-D. The BAP and NAA took longer time to produce callus, and caused the formation of poor and compact calli with many necrotic sites. Successful shoot regeneration was achieved from white compact callus. 2 mg/L BAP is better for shoots induction in both genotypes. The highest percentage of shoots producing roots and mean number of roots per explants was achieved at the concentration of 0.3 mg/L of IBA. The survival rate of regenerated plantlet was 100%.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interest.
ACKNOWLEDGEMENTS
Special thanks go to the Plant Biotechnology Laboratory of the Holleta Agricultural Research Center, for their generous help and we wish to thank Dr. Adefris Teklewold from the Ethiopian Institute of Agricultural Research for providing the necessary facilities and encouragement.
REFERENCES
| Abellatef E, Khalafallah MM (2008). Influence of Growth Regulators on Callus Induction from Hypocotyls of Medium Staple Cotton (Gossypiumhirsutum L). Cultivar Barac B -67. J. Soil Nat. 2(1):17-22. | ||||
| Ali H, Zahir A, Haidar A, Mehmood S, Wiqar A (2007). In vitro regeneration of brassica napusl., cultivars (star, cyclone and westar) from hypocotyls and cotyledonary leaves. Pak. J. Bot. 39(4): 1251-1256. | ||||
| Al–Naggar AM, Shabana R, Rady MR, Ghanem SA, Saker MM, Reda AA (2008) . In Vitro callus initiation and Regeneration in some conala varieties. Int. J. Acad. Res. 2:36-89. | ||||
|
Babic V, Datla RS, Scoles GJ, Keller WA (1998). Development of an Efficient Agrobacterium–Mediated Transformation System for Brassica carinata. Plant Cell Rep. 17:183-188. Crossref |
||||
| Bano RM, Khan H, Rashid H, Swati Z (2010). Development of an efficient regeneration protocol for three genotypes of brassica juncea. Pak. J. Bot. 42(2):963-969. | ||||
|
Barfield DG, Pua EC (1991). Gene Transfer in Plants of Brassica juncea using Agrobacterium tumefaciens Mediated Transformation. Plant Cell Rep. 10(6-7):308-314. Crossref |
||||
| Bozzini A, Calcagno F, Soare T (2007). "Sincron", a new Brassica carinata cultivar for biodiesel production. HELIA 46:207-214. | ||||
|
Caboni E, Tonalli MG (2002). Effect of 1, 2-Benzisoxazole-3 acetic acid on adventitious shoot regeneration and in vitro rooting in apple. Plant Cell Rep. 18:985-988. Crossref |
||||
|
Chamandosti F, Majd A, Mehrabian S(2006). In vitro plant regeneration from callus of cotyledons in canola (Brassica napus L). Pak. J. Biol. Sci. 9 (2)302-306. Crossref |
||||
| CSA (Central Statistical Authority) (2008/09). Report on land utilization: Private peasant holdings, 'Meher' season. Statistical Bulletin 302. Addis Ababa, Ethiopia | ||||
|
Datta GS, Conger BV(1999). Somatic embryogenesis and plant regeneration from suspension cultures of Switch grass. Crop Sci. 39: 243-247. Crossref |
||||
|
Deblock M, Debrouwer D, Tenning P (1989).Transformation of Brassica napusand Brassica oleracea Using Agrobacterium tumefaciens and the Expression of the bar and neoGenes in the Transgenic Plants. Plant Physiol. 91:694-701. Crossref |
||||
|
Dietert MF, Barron SA, Yoder OC (1982). Effects of genotype on in vitro culture in the genus Brassica. Plant Sci. Lett. 26:233–240. Crossref |
||||
| EARO (Ethiopian Agricultural Research Organization) (2000). Crop Research Directorate, High land oil crops research strategy, Addis Ababa, Ethiopia. | ||||
|
Fang Yu, Zhang D, Fengwu B, Lijia A (2005). The accumulation of Isocamptothecin A and B in suspension cell cultures of Camptotheca acuminate. Plant Cell Tissue Organ Cult. 81:159-163. Crossref |
||||
| George EF, Hall MA, Klerk GJD (2008). Plant Propagation by Tissue Culture: Volume 1. Third Edition, Springer Publisher: Dordrecht; London. | ||||
| Greville LTD(2005). Plants for a Future, Blagdon Cross, Ashwater, Beaworthy, Devon, EX215DF,UK. | ||||
| Gubis J, Lajchová Z, Faragó J, Jureková Z (2004). Effect of growth regulators on shoot induction and plant regeneration in tomato (LycopersiconesculentumMill.). Biol. Bratislava 59(3):405-408. | ||||
| Guo D P, Zhu Z J, Hu XX, Zheng SJ(2005). Effects of cytokinins on shoot regeneration from cotyledon and leaf segment of stem mustard Brassica juncea var. Tsatsai. Plant Cell Tissue Org. Cult. 83:123-127. | ||||
| Gupt V, Sita GL, Shaila MS Jagannathan V (1993). Genetic Transformation of Brassica nigraby Agrobacterium Based Vector and Direct Plasmid Uptake. Plant Cell Rep. 12(7-8):418-421. | ||||
|
Hansen G, Wright MS(1999). Recent advances in the transformation of plants. Trends Plant Sci. 4:226-231. Crossref |
||||
| Kamal GB, Illich KG, Asadollah A (2007). Effects of genotype explant type and nutrient medium components on canola (Brassica napus L.) shoot in vitro organogenesis. Afr. J. Biotechnol. 6 (7): 861-876. | ||||
|
Katiyar RK, Saran G, Giri G (1986). Evaluation of Brassica carinataas a New Oilseed Crop in India. Exp. Agric. 22(1):67-70. Crossref |
||||
| Khan MR, Rashid H, Quraishi A (2010). Increased regeneration efficiency of Brassica napusL. cultivars Star, Westar and Cyclone from hypocotyle and cotyledonary explants. Plant Tissue Cult.12: 131-138. | ||||
|
Khan MR, Rashid H (2002). Effects of various growth regulators on callus formation and regeneration in Brassica napuscv. Oscar. Pak. J. Biol. Sci. 5(6):693-695. Crossref |
||||
|
Khehra GS, Mathias RJ (1992). The interaction of genotype, explant and media on the regeneration of shoots from complex explants of Brassica napus L. J. Exp. Bot. 43:1413-1418. Crossref |
||||
|
Liu JW, DeMichele S, Bergana S, Bobik M, Hastilow EC, Chuang L, Mukerji TP, Huang Y S(2001). Characterization of Oil Exhibiting High Gamma–Linolenic Acid from a Genetically Transformed Canola Strain. J. Am. Oil Chem. Soc. 78(5):489-493. Crossref |
||||
| Magdoleen G, Elsadig O, Elhadi A, Mutasim M (2010). Callus formation and organogenesis of tomato (LycopersiconesculentumMill, C.V. Omdurman) induced by thidiazuron. Afr. J. Biotechnol. 9(28):4407-4413. | ||||
|
Malik RS (1990). Prospects for Brassica carinataas an Oilseed Crop in India. Exp. Agric. 26(1):125-129. Crossref |
||||
|
Malik SK, Chaudhury R, Rajwant K(2005).Rapid in vitro multiplication and conservation of Garciniaindica: A tropical medicinal tree species Sci. Hortic. 106:539-53. Crossref |
||||
| Moghaieb REA, El–Awady MA, Mergawy RG, Youssef SS,El-SharkawyAM(2006). A reproducible protocol for regeneration and transformation in canola g (Brassica napus L). Afr. J. Biotechnol. 5(2): 143-148. | ||||
|
Moloney MM, Walker JM, Sharma KK (1989). High Efficiency Trans-formation of Brassica napus Using Agrobacterium Vectors. Plant Cell Rep. 8(4):238-242. Crossref |
||||
|
Ono Y, Takahata Y, Kaizuma N(1994). Effect of genotype on shoot regeneration from cotyledonary explants of rapeseed (Brassica napus L.). Plant Cell Rep. 14:13-17. Crossref |
||||
| Pathirana R, Eason JR(2006). Establishment and characterization of a rapidly dividing diploid cell suspension culture of Arabidopsis thaliana suitable for cell cycle synchronization. Plant Cell Tissue Org. Cult. 85: 125-136. | ||||
|
Phogat SK, Burma PK, Penta lD(2000). High frequency regeneration of Brassica napus varieties and genetic transformation of stocks containing fertility restorer genes of two cytoplasmic male sterility systems. J. Plant Biochem. Biotechnol. 9:73-79. Crossref |
||||
|
Radke SE, Turner JC, Facciotti D(1992). Transformation and Regeneration of Brassica rapa Using Agrobacteriumtumefaciens. Plant Cell Rep. 11(10):499-505. Crossref |
||||
| Rao AQ, Hussnain SS, Shahzad MS, Bokhari S, Yassir AHT, Rizuddin S(2006). Somatic embrogenesis in wild relatives of cotton (Gossipiumspp.). J. Zhejiang 291-298. | ||||
|
Salman MN (2002). Establishment of callus and cell suspension cultures from Gypsophila paniculata leaf segments and study of the attachments of host cells by Erwiniaherbicolapv. gypsophilae. Plant Cell Tissue Organ Cult. 69(2): 189-196. Crossref |
||||
| Sayed AI, Sh H, El-MorsyI, El-Said RA (2010).in vitro culture of somatic explants and anther cultures of canola (Brassica napus L.). J. Agric. Chem. Biotechnol. 1(1):23-39 | ||||
| Sparrow AC, Snape JW, Dale PJ, Irwin JA (2006). The Rapid Identification of B. napusGenotypes, for High-Throughput Trans-formation, Using Phenotypic Tissue Culture Markers. Acta Hortic. 706:239-247. | ||||
|
Ullah I, Rashid H, Khan MR (2004). Establishment of tissue culture protocol in Brassica (Brassica napusL.). Pak. J. Biol. Sci. 7:277-278. Crossref |
||||
|
Yang MZ, Jia SR, Pua EC (1991). High frequency of plant regeneration from hypocotyls explants of Brassica carinataA.Br. Plant Cell Tissue Organ Cult. 24:79-82. Crossref |
||||
|
Yu F, Zhang D, Bai F, An L (2005). The accumulation of isocamptothecin A and B in suspension cell culture of Camptotheca acuminata. Plant Cell.Tissue Organ Cult. 81:159-163. Crossref |
||||
| Zeynali M, Bahram EA, Mohammad MN, Aghajari SM (2010). Influence of genotype and plant growth regulator on somatic embryogenesis in rapeseed (Brassica napus L.). Afr. J. Biotechnol. 9(26):4050-4055. | ||||
|
Zhang FL, Takahata Y, Xu JB (1998). Medium and genotype factors influencing shoot regeneration from cotyledonary explants of Chinese cabbage (Brassica campestris L. ssp. pekinensis). Plant Cell Rep. 17:780-786. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


