Full Length Research Paper

ABSTRACT
INTRODUCTION
The Manila clam, Ruditapes philippinarum, is an economically-important marine bivalve species of the China aquaculture industry and is widely distributed in the coasts of China. The world production of this species was 3.9 million metric tons in 2013. China is the first largest country in the world in terms of production of the Manila clam, producing about 3.0 million metric tons annually, which accounts for about 90% of global production (Zhang and Yan, 2010). In aquaculture, genetic diversity is the fundamental resource on which stock improvements rely. However, many aquaculture practices, such as producing large numbers of offspring from a few parents, inbreeding and using broodstocks derived from hatchery seed, are likely to reduce genetic diversity and thereby diminish disease resistance and reduce the population's ability to adapt to new environments (Allendorf and Phelps, 1980). In recent years, the wild resources of R. philippinarum have experienced dramatic population declines due to over-exploitation and the deterioration of coastal environment.
The significant decline of wild R. philippinarum stocks makes people to pay close attention to its genetic variation and population structure which will provide essential information on maintenance and management of the clam resources (An et al., 2012; Mura et al., 2012; Xing et al., 2014).
Microsatellite or simple sequence repeat (SSR) markers, which are inherited in a Mendelian fashion as codominant markers, have been increasingly popular in genetic studies because of their high levels of allelic variability, wide dispersal and abundance throughout the genome (Chistiakov et al., 2006). Until recently, microsatellite markers have been developed in the R. philippinarum derived from both expressed sequence tag (EST) and anonymous genomic sequence (Yasuda et al., 2007; An et al., 2009; Nie et al., 2014). Molecular markers can be divided into type I (coding) markers which are associated with genes of known functions and type II (noncoding) markers which are associated with anonymous genomic sequences (O’Brien, 1991). As Type I markers that are associated with genes of known function, the EST-SSRs are superior to anonymous genomic SSR in functional diversity assessment and interspecific transferability (Pashley et al., 2006), but genomic SSRs usually are more polymorphic than EST-SSRs (Ellis and Burke, 2007).
About 60 microsatellite markers were developed in R. philippinarum until now (Yasuda et al., 2007; An et al., 2009; Hu et al., 2014), including 36 genomic SSRs and 25 expressed sequence tag derived SSRs (EST-SSRs). These markers provide sufficient information to evaluate wild and cultured genetic resources, but are still deficient for the development of genetic linkage map and marker-assisted selection (MAS). Therefore, the objective of the present study was to develop more polymorphic microsatellite markers using the transcriptome data derived from 454 pyrosequencing.
MATERIALS AND METHODS
Samples collection and DNA extraction
Thirty five individuals of R. philippinarum in total were collected from Jinzhou, Dalian, Liaoning province China. Genomic DNA of each specimen was isolated from muscle tissues following the standard phenol-chloroform method (Li et al., 2006) with some modifications. The adductor muscle was removed from fresh specimens and preserved in 100% ethanol until DNA preparation. Tissue was homogenized in 500 μL of extraction lyses buffer together with 0.5 μg/mL proteinase K and incubated at 55°C. Following phenol: chloroform: isoamyl alcohol (25:24:1) extractions, the supernatants were precipitated by the addition of 2 volumes of absolute ethanol. DNA was washed with 70% ethanol, dissolved in TE and stored at -20°C.
Microsatellite primer design and PCR
Microsatellite sequences were screened from transcriptome sequences derived from R. philippinarum in our laboratory. Microsatellite sequences were screened from a total of 9450 ESTs in the 454 database using the software SSRHUNTER 1.3 (Li and Wan, 2005). Microsatellite primers were designed using Primer Premier 5.0 software (http: //www. premierbiosoft.com/ primerdesign/). From 9450 sequences, 324 were identified with microsatellite motifs, and a set of 105 microsatellite primer pairs were designed and synthesized. The major parameters for primer design were set as follows: primer length from 19 to 25 nucleotides, the size of PCR product from 100 to 350 bp, and annealing temperature at 50-65°C. The primers were synthesized by Sangon Company (Shanghai). ESTs containing SSRs were then annotated using BLAST software as described by Maneeruttanarungroj et al. (2006). The BLAST results were classified into 3 groups: known gene products, hypothetical proteins and unknown genes.
Polymorphism assessment for primers
The polymorphisms of microsatellite primers were tested in 35 individuals of R. philippinarum. Of the 105 potential microsatellite markers, 39 were not easily amplified, and 38 were found to be polymorphic among 8 individuals of R. philippinarum. Then, thirty eight microsatellite markers were selected to test polymorphic and genetic diversity of natural population of R. philippinarum in Jin Zhou, Dalian, China.
Polymerase chain reaction (PCR) was performed in 10-μl volumes containing 0.5 U easy Taq DNA polymerase (TransGen, Beijing), 1× PCR buffer, 0.2 mM dNTP, 0.4 μM of each primer set, 1.5 mM MgCl2, and about 25 ng template DNA. The reactions were performed using the following parameters: 3 min at 94°C, followed by 35 cycles of 45 s at 94°C, 45 s at the annealing temperature listed in Table 1 and 45 s at 72°C, then a final extension of 5 min at 72°C. Amplification products were resolved on a 8% polyacrylamide gel and visualized by silver staining.
Data analysis
The number of alleles, and observed (HO) and expected (HE) heterozygosities were estimated by MICROSATELLITE ANALYSER software (Dieringer and Schlötterer, 2003). Tests for linkage disequilibrium (LD) and deviations from Hardy-Weinberg equilibrium (HWE) were performed by GENEPOP 4.0 (Rousset, 2008). Sequential Bonferroni corrections (Rice, 1989) were applied for all multiple tests (P < 0.05). MICRO-CHECKER (Van Oosterhout et al., 2004) was employed to infer the most probable technical cause of HWE departures, including null alleles, mis-scoring due to stuttering and allelic dropout due to short allele dominance.
RESULTS AND DISCUSSION
Microsatellites, which are inherited in a Mendelian fashion as codominant markers, have been increasingly popular in genetic studies because of their high level of heterozygosity, wide dispersal and abundance throughout the genome and transferability across different strains. For R. philippinarum, numerous microsatellites have been developed recently (Yasuda et al., 2007; An et al., 2009; Hu et al., 2014). But, the pace of development has been limited by the time-consuming and labor intensive requirement to construct, enrich and sequence genomic libraries (Edwards et al., 1996). Recently, identification of microsatellites from expressed sequences has been extensively used as an alternative strategy.
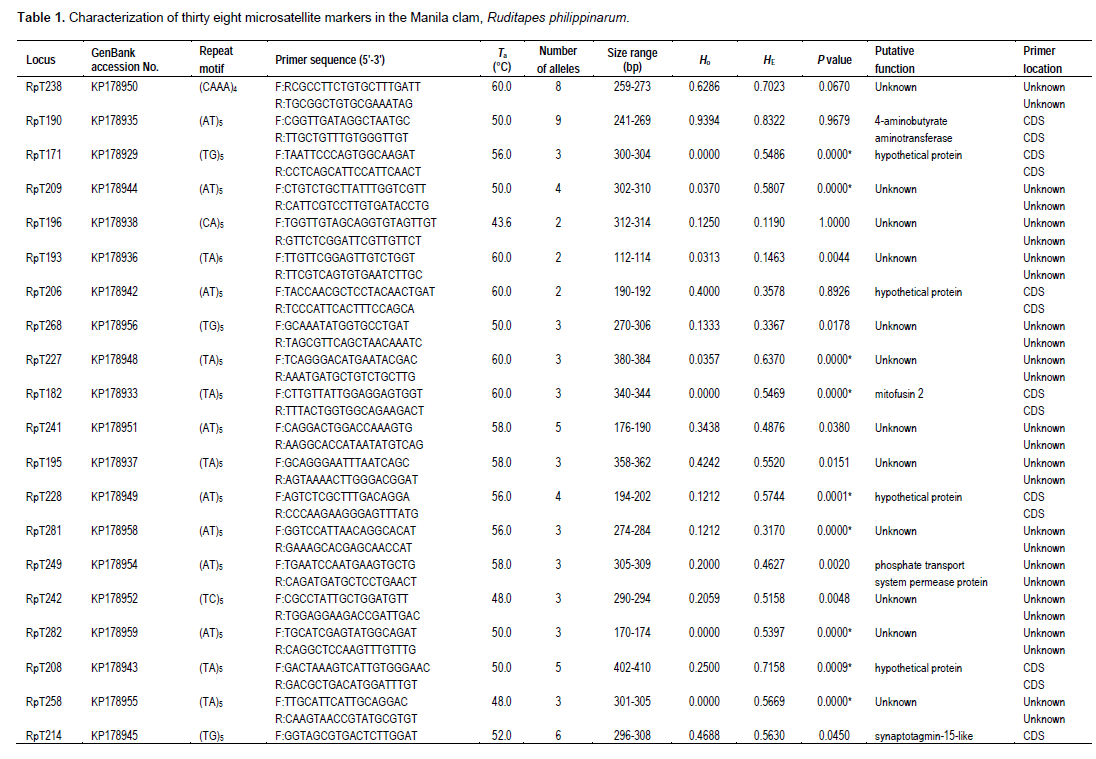
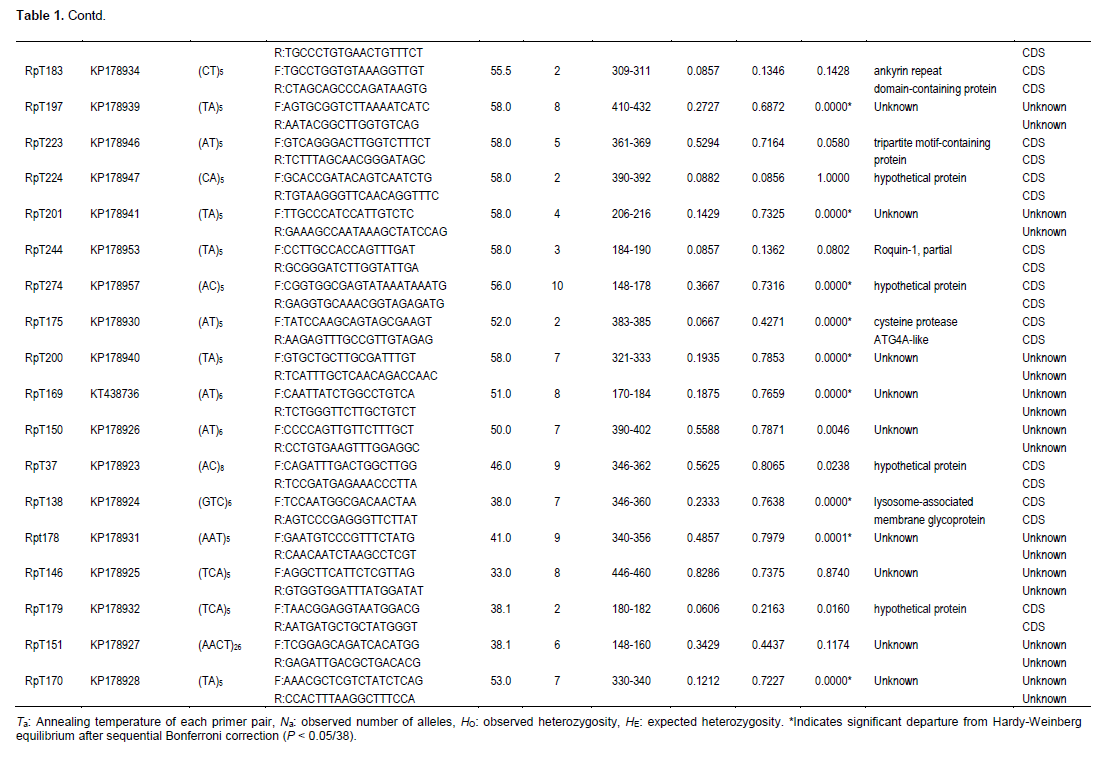
In addition to requiring less time and money to develop, expressed sequence tags (EST)-derived microsatellites have a number of intrinsic advantages. They tend to be more widely transferable between species, and even genera (Bouck and Vision, 2007). More importantly, because they represent genes, they serve as type I markers, which are more valuable for comparative gene mapping (Liu et al., 1999). In this study, a total of 324 microsatellite-containing sequences were identified from 9,450 transcriptome sequences in the R. philippinarum 454 database. Of the 324 sequences, 105 were selected for microsatellite marker optimization because of repetition times and flaking sequence priority. Of the 105 potential microsatellite markers, 39 were not easily amplified, 28 were monomorphic, and 38 were found to be polymorphic among 35 individuals of R. philippinarum.
GenBank (BLAST) searches indicated that 17 of the 38 EST-SSRs matched genes of known functions at E values less than 10−4, whereas the other 21 had no significant matches to known genes (Table 1). Of the 105 primer pairs developed, 38 microsatellite loci (36.2%) showed polymorphism in the population of R. philippinarum (Table 1). The 38 polymorphic loci (36.2%) yielded 2 to 10 alleles per locus with an average of 4.8 (Table 1). The observed and expected heterozygosities ranged from 0.000 to 0.939 and from 0.086 to 0.832, with an average of 0.255 and 0.542, respectively (Table 1). Significant linkage disequilibrium was detected between 12 pairs of loci (RpT282/RpT258, RpT209/RpT282, RpT193/RpT197, RpT208/RpT223, RpT268/RpT201, RpT200/RpT249, RpT182/RpT150, RpT175/RpT150, RpT238/RpT178, RpT178/RpT146, RpT138/RpT179 and RpT258/RpT170) (P <0.01) before sequential Bonferroni correction for multiple tests (Rice, 1989); however, only 3 pairwise combinations of 6 loci (RpT209/RpT282, RpT178/RpT146 and RpT200/RpT249) were significant after Bonferroni correction.
Twenty loci conformed to HWE, while the remaining 18 loci showed significant deviation from HWE after Bonferroni correction at 5% significance level (Table 1). MICRO-CHECKER (Van Oosterhout et al., 2004) was used to estimate the most probable cause of departures from HWE. Micro-Checker analysis suggested that there was no evidence for scoring error due to stuttering and no evidence for large allele dropout. All the 18 loci were prone to null alleles (P < 0.01). Widespread null alleles have been reported for approximately 51.9% of loci in the Pacific oyster (Li et al., 2003) and 52.2% of loci in the Zhikong scallop (Zhan et al., 2009). A high percentage of primers containing variable nucleotides, such as base substitutions or deletions at the PCR-priming sites in the flanking region of the microsatellites, may be responsible for the widespread appearance of null alleles in bivalves (Hedgecock et al., 2004).
The results obtained in this study indicated that these SSRs developed from EST in the Manila clam will be a useful tool for the genetic research such as population variation, parentage analysis, stock enhancement eva-luation and the establishment of effective conservation strategy of R. philippinarum.
CONFLICT OF INTERESTS
The author has not declared any conflict of interest.
ACKNOWLEDGEMENTS
This study was supported by the National Natural Science Foundation of China (31302183), the Program for Liaoning Excellent Talents in University (LJQ2014076), the Cultivation Plan for Youth Agricultural Science and Technology Innovative Talents of Liaoning Province (2014004), the Modern Agro-industry Technology Research System (CARS-48) and the National High Technology Research and Development Program (2012AA10A410-2).
REFERENCES
|
Allendorf FW, Phelps SR (1980). Loss of genetic variation in a hatchery stock of cutthroat trout. Trans. Am. Fish. Soc. 109:537-543. |
|
|
An HS, Kim EM, Park JY (2009). Isolation and characterization of microsatellite markers for the clam Ruditapes philippinarum and cross-species amplification with the clam Ruditapes variegate. Conserv. Genet. 10:1821-1823. |
|
|
An HS, Park WJ, Cho KC, Han HS, Myeong J-I (2012). Genetic structure of Korean populations of the clam Ruditapes philippinarum inferred from microsatellite marker analysis. Biochem. Syst. Ecol. 44:186-195. |
|
|
Bouck A, Vision T (2007). The molecular ecologist's guide to expressed sequence tags. Mol. Ecol. 16: 907-924. |
|
|
Chistiakov DA, Hellemans B, Volckaert FAM (2006). Microsatellites and their genomic distribution, evolution, function and applications: A review with special reference to fish genetics. Aquaculture 255:1-29. |
|
|
Dieringer D, Schlötterer C (2003). Microsatellite analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 3:167-169. |
|
|
Edwards KJ, Barker JH, Daly A, Jones C, Karp A (1996). Microsatellite libraries enriched for several microsatellite sequences in plant. Biotechniques 20:758-760. |
|
|
Ellis J, Burke J (2007). EST-SSRs as a resource for population genetic analyses. Heredity 99:125-132. |
|
|
Hedgecock D, Li G, Hubert S, Bucklin K, Ribes V (2004). Widespread null alleles and poor cross-species amplification of microsatellite DNA loci cloned from the Pacific oyster, Crassostrea gigas. J. Shellfish Res. 23:379-385. |
|
|
Hu GW, Yan XW, Zhu DP, Nie HT (2014). Isolation and characterization of fourteen polymorphic microsatellite loci in the Manila clam (Ruditapes philippinarum). Conserv. Genet. Resour. 6:251-253. |
|
|
Li G, Hubert S, Bucklin K, Ribes V, Hedgecock D (2003). Characterization of 79 microsatellite DNA markers in the Pacific oysters Crassostrea gigas. Mol. Ecol. Notes 3:228-232. |
|
|
Li Q, Wan JM (2005). SSRHUNTER: development of a local searching software for SSR sites. Hereditas 27:808-810. |
|
|
Li Q, Yu H, Yu RH (2006). Genetic variability assessed by microsatellites in cultured populations of the Pacific oyster (Crassostrea gigas) in China. Aquaculture 259:95-102. |
|
|
Liu ZJ, Tan G, Li P, Dunham RA (1999). Transcribed dinucleotide microsatellites and their associated genes from channel catfish, Ictalurus punctatus. Biochem. Biophys. Res. Commun. 259:190-194. |
|
|
Maneeruttanarungroj C, Pongsomboon S, Wuthisuthimethavee S, Klinbunga S, Wilson KJ, Swan J, Li Y, WhanV, Chu KH, Li CP, Tong J, Glenn K, Rothschild M, Jerry D, Tassanakajon A (2006). Development of polymorphic expressed sequence tag-derived microsatellites for the extension of the genetic linkage map of the black tiger shrimp (Penaeus monodon). Anim. Genet. 37:363-368. |
|
|
Mura L, Cossu P, Cannas A, Scarpa F, Sanna D, Dedola GL (2012). Genetic variability in the Sardinian population of the manila clam, Ruditapes philippinarum. Biochem. Syst. Ecol. 41:74-82. |
|
|
Nie HT, Zhu DP, Yang F, Zhao LQ, Yan XW (2014). Development and characterization of EST-derived microsatellite makers for Manila clam (Ruditapes philippinarum). Conserv. Genet. Resour. 6:25-27. |
|
|
O'Brien SJ (1991). Molecular genome mapping lessons and prospects. Curr. Opin. Genet. Dev. 1:105-111. |
|
|
Pashley CH, Ellis JR, McCauley DE, Burke JM (2006). EST Databases as a source for molecular markers: Lessons from Helianthus. J. Hered. 97(4):381-388. |
|
|
Rice RW (1989). Analyzing tables of statistical tests. Evolution 43:223-225. |
|
|
Rousset F (2008). GENEPOP'O007: a complete reimplementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 8:103-106. |
|
|
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P (2004). MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes. 4:535–538. |
|
|
Xing K, Gao ML, Li HJ (2014). Genetic differentiation between natural and hatchery populations of Manila clam (Ruditapes philippinarum) based on microsatellite markers. Genet. Mol. Res. 13(1):237-245. |
|
|
Yasuda N, Nagai S, Yamaguchi S, Lian CL, Hamaguchi M (2007). Development of microsatellite markers for the Manila clam Ruditapes philippinarum. Mol. Ecol. Notes 7:43-45. |
|
|
Zhan A, Hu J, Hu X, Hui M, Wang M, Peng W, Huang X, Wang S, Lu W, Sun C, Bao Z (2009) Construction of microsatellite-based linkage maps and identification of size-related quantitative trait loci for Zhikong scallop (Chlamys farreri). Anim. Genet. 40:821-831. |
|
|
Zhang GF, Yan XW (2010). Clam aquaculture. Science Press, Beijing, China. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


