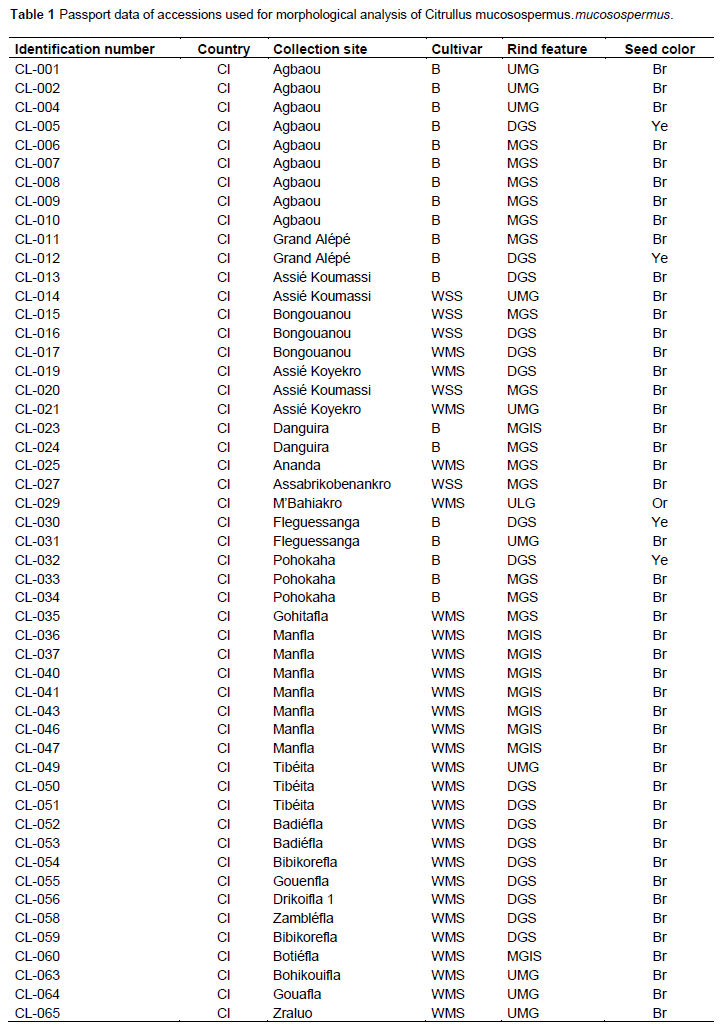
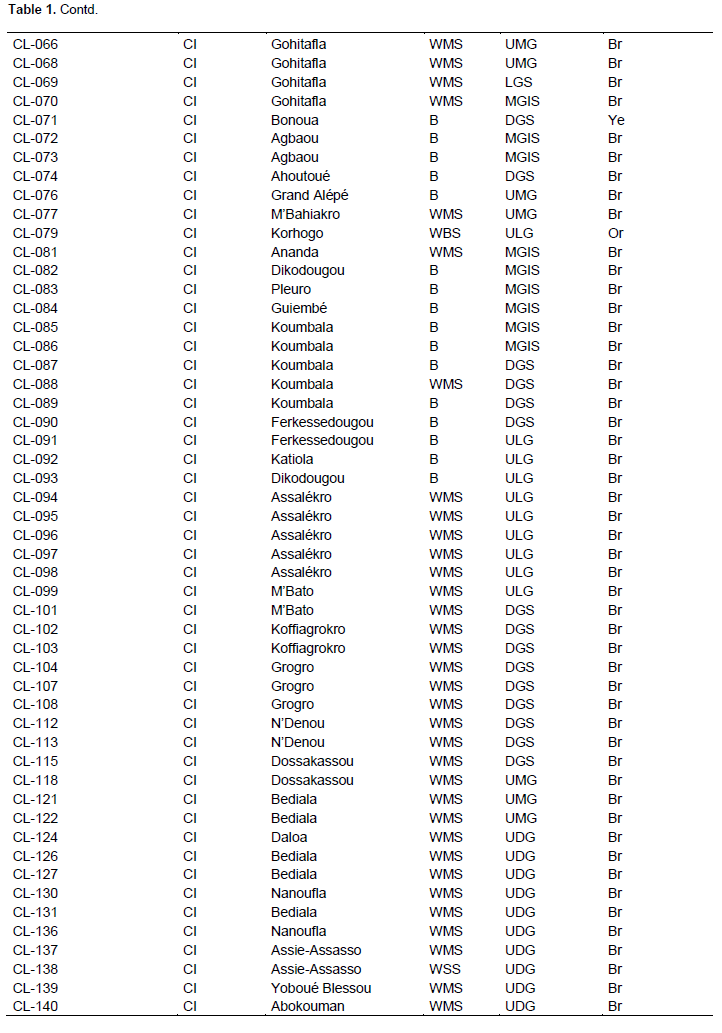
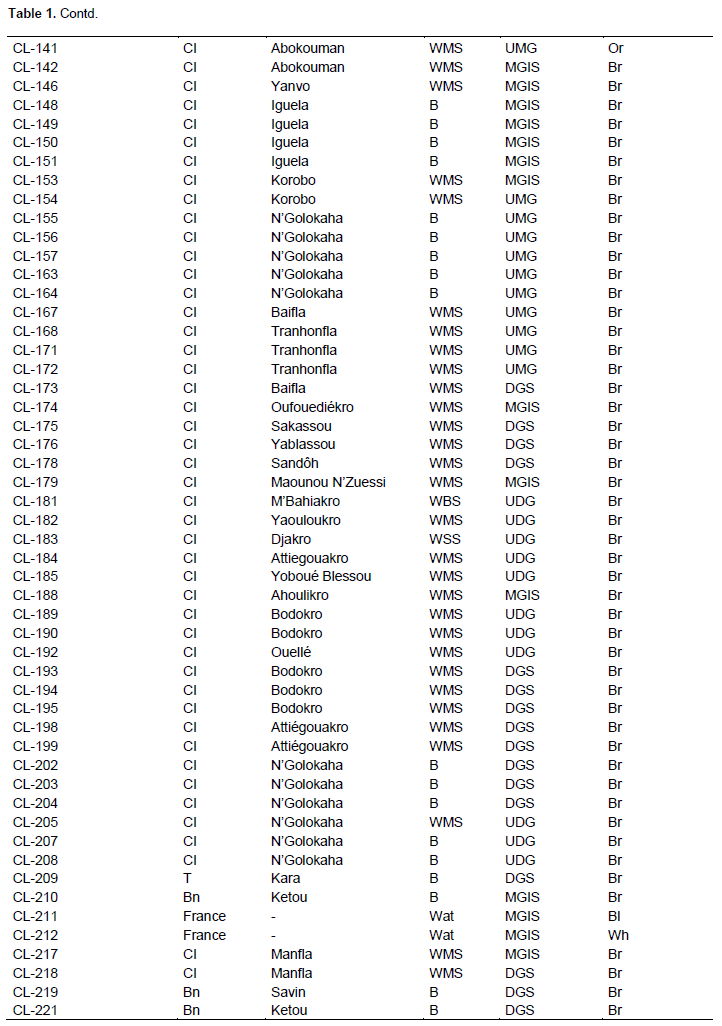
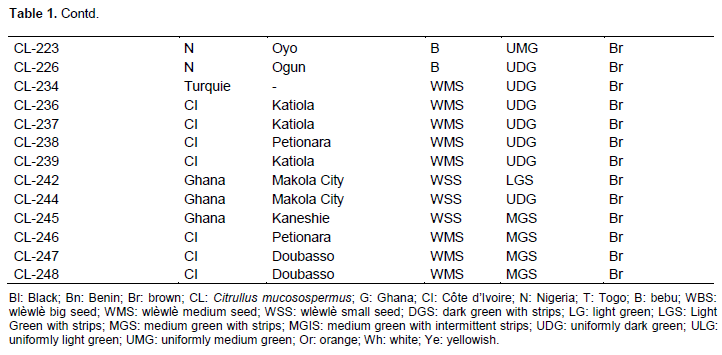
Full Length Research Paper

ABSTRACT
A hundred and seventy-one oleaginous watermelon accessions either collected from different countries or obtained from gene banks were evaluated and compared based on 11 quantitative morphological traits. Principal component analysis on 11 traits revealed 81.19% of the total variability and pointed out variations among accessions, mainly on the basis of fruit size and weight. The dendrogram and factorial discriminant analysis clustered the accessions in four groups. The multivariate analysis of variance showed a significant difference between the four groups and accessions of the group 2 had higher agronomic performances. The confusion matrix gave the details about accessions assignment and pointed out that breeders must have recourse to several multivariate analyses to have better agromorphological classification of accessions. The traits related to fruit size and weight were the most accession distinctive. Consequently, fruit morphological traits could be used for accession identification during collecting missions.
Key words: Citrullus mucosospermus, morphological characterization, multivariate analysis, oleaginous cucurbit.
INTRODUCTION
Cucurbits are present in both the New and Old World and are among the most important plant families that supply human with edible products and useful fibers. Cucurbits are divided into five sub-families: Fevilleae, Melothrieae, Cucurbitaceae, Sicyoideae, and Cyclanthereae.The genus Citrullus Schrad. ex Eckl et Zeyh placed closest to a clade formed by Peponium and Lagenaria, the latter including the bottle gourd (Lagenaria siceraria), conï¬rming previous ï¬ndings(Schaefer and Renner, 2011). Citrullus, member of the cucurbit family, includes seven species: (1) Citrullus lanatus (Thunb.) Matsum. et Nakai (2n = 22), found in tropical and subtropical climates worldwide; (2) Citrullus amarus Schrad. 1836 syn. C. caffer Schrad. 1838, also known as C. lanatus var. caffrorum (Alef.) Fosb. or C. lanatus var. citroides (L.H. Bailey) Mansf.; it is the preserving melon grown in Southern Africa and called tsamma melon (Whitaker and Bemis, 1976) is used to make jams since at least the ï¬fteenth century (Bailey, 1930); (3) Citrullus mucosospermus Fursa, the so-called egusi melon, largely referred to as a subspecies of C. lanatus by many authors (including recently(Hammer and Gladis, 2014)), but which was earlier raised at specific rank(Fursa, 1972, 1981, 1983); (4) Citrullus colocynthis (L.) Schrad. (2n = 22), a perennial species growing in sandy areas throughout Northern Africa and adjacent Asia; (5) Citrullus ecirrhosus Cogn., another perennial wild species (Meeuse, 1962); (6) Citrullus rehmii De Winter, an annual wild species (de Winter, 1990), and (7) Citrullus naudinianus (Sond.) Hook.f., from the Namib-Kalahari region.
Watermelon (Citrullus vulgaris) (Cucurbitaceae) includes several other economically important species such as cucumber (Cucumis sativus), melon (Cucumis melo L), squash (Cucurbita pepo), calabash (L. siceraria), and pumpkin (Citrullus maxima). C. lanatus is an annual species, which has wild, cultivated, and feral forms.
In the ï¬nding that the watermelon and its sister species are West African plants suggests that the natural range of watermelon may have extended into Libya or into Egypt during more humid periods of the Pleistocene and Holocene(Schulz, 1987, 1991). Alternatively, watermelon seeds may have been traded from West Africa to Northern Africa. The illustrations found in Egyptian tombs of watermelon served on a tray suggest that these fruits were eaten raw, perhaps as a dessert. Seeds found in ancient Egyptian tombs, including that of Thutankhamun (Hepper, 1990) should ideally be studied using ancient DNA approaches.
Genetically, the cultivated watermelon is closest to plants from West Africa that represents the gene pool from which watermelon was domesticated (Chomicki and Renner, 2014). The sister species of watermelon is C. mucosospermus, the egusi melon from Nigeria to Senegal and described by Fursa (1983), Che et al. (2003), and Dane and Liu (2007). In consideration of the findings by Fursa (1983)and Chomicki and Renner (2014), the egusi-type watermelon is recognized as C. mucosospermus throughout this paper.
The "egusi" watermelon of West Africa is cultivated for its oleaginous and nutritious seeds that are important in the social and cultural live of several peoples (Burkill, 1985; Oyulu, 1977; van der Vossen et al., 2004; Achigan-Dako et al., 2008). For example, dried slightly toasted and ground seeds are used as soup thickener. The "egusi" seeds are reported to be rich in nutrients ~60% lipids and ~30% proteins (Loukou et al., 2007). Edible oil can also be extracted from the seeds. Commonly found in many traditional cropping systems, the plant is well adapted to extremely divergent agro-ecosystems and various cropping systems characterized by minimal inputs. C. lanatus thus represents an excellent plant model for which improved cropping systems implementation can insure the economic prosperity of rural women that are the main producers in tropical Africa. The west African watermelon is generally considered as allogamous, monoecious, entomophilous, and protandrous(Gusmini and Wehner, 2003). This species is named as C. lanatus var. lanatus or as C. lanatus subsp. Mucosospermus (Fursa, 1972) or C. colocynthis(Hutchinson and Dalziel, 1954).
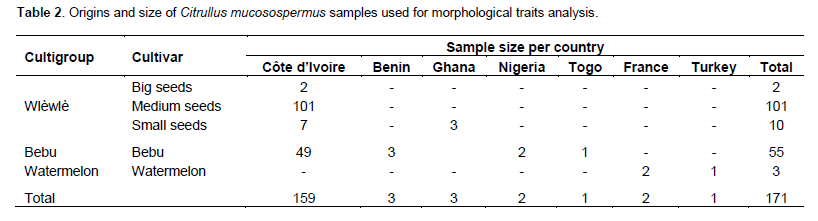
Particularly, in Côte d’Ivoire, indigenous and minor crops for local consumption have almost disappeared in favor of exotic species. Some farmers still continue to produce these crops not far from their home or on small plots cleared after harvest of the main annual crops (yams, corn, peanut, etc.) (Zoro Bi et al., 2003). Thus, some of them such as oleaginous cucurbits are still found in the market. According to Zoro Bi et al. (2003) and Djè et al. (2006), C. mucosospermus (but identified as C. lanatus by these authors in Côte d’Ivoire) is the species most widely distributed and most commercialized in Côte d’Ivoire. This plant is mainly grown in savanna areas in the North, in the pre-forest areas in Central, East, and South(Djè et al., 2006). Two distinct cultigroups have been described on the basis of the morphology of seeds. The first one (wlèwlè) is characterized by glossy seeds with a tapered proximal extremity. This morphotype is subdivided into three cultivars according to the size of the seeds(Djè et al., 2006). Fruit color of this cultigroup, is almost uniform, greenish or whitish. The second (bebu), with one cultivar has green fruit also, but with dark longitudinal bands. The seeds are heavier and have a flat ovoid shape with rugged and thick ends (Zoro Bi et al., 2003).Although the oleaginous type of Citrullus is an economical important crop in west Africa. Few studies were conducted on genetic resources of this plant (Ndabalishye, 1995; Djè et al., 2006) previously reported to be morphological differentiated especially in terms of fruit and seed characters.
The objectives of the present study were to survey and analyze variations in plant, fruit and seed morphology in C. lanatus oleaginous type collection from Côte d’Ivoire. This represents the first study on collection and characterization of C. lanatus oleaginous type germplasm in Nangui Abrogoua University.
MATERIALS AND METHODS
Plant and collection sites
In this study, 171 accessions of C. lanatus oleaginous type were used. They were provided from seven countries: Benin, Côte d’Ivoire, France, Ghana, Nigeria, Togo, and Turkey (Table 1). The samples were collected from farmers’ stock, genebank or purchased from local seed markets. The selected accessions were divided into four cultivars defined on the basis of seed size and designated small-, medium-, big-seeded, and thickened margin seeds. Thickened margin seeds size varies between 153 and 205 mm², big seeds from 125 to 151 mm², medium seeds from 86 to 110 mm², and small seeds from 41 to 52 mm²(Djè et al., 2006). Accessions (2 to 101) were sampled per cultivar, according to seeds availability (Table 2).
Study site
The trials were regularly monitored throughout the growing season in 2011 and 2012 (May to November). Farm experiment was conducted in the village of Manfla, located in the centre (6°49′34.38″N, 5°43′47.68″W) 400 km North Abidjan (Côte d’Ivoire). Annual rainfall varies from 800 to 1400 mm with a long-term mean of 1200 mm, and the annual mean temperature is 27°C. A complete description of study site was done by Kouassi and Zoro Bi (2010).
Experimental design
Planting was done according to a completely randomized block design, with seven replications. Each replicate consisted of a 24×27 m containing 70 holes at a depth of 3 cm, resulting in 7 holes per accession. The planting distance was 3 m between and within rows with 1.5 m of edges. A manual weeding was carried out during plant development to prevent weed invasion. Disease and pest control was carried out using a carbamate-based insecticide applied when necessary.
Traits measurement
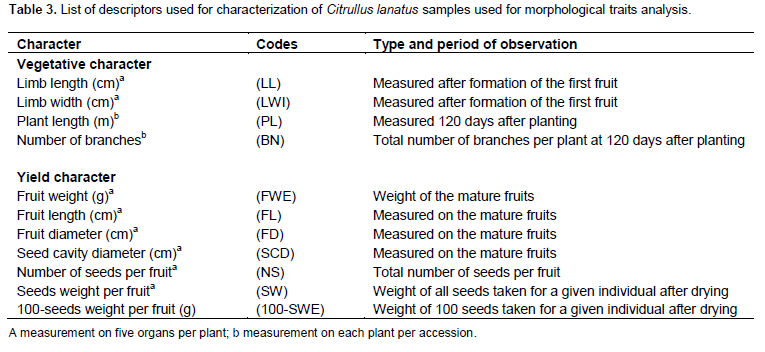
Accessions were examined using 11 quantitative traits subdivided into four vegetative and seven yield characters selected from standard descriptors for cucurbits(Maggs-Kölling et al., 2000; Marr et al., 2007; Morimoto et al., 2005; Koffi et al., 2008). Data were measured in five plants and five fruits per accession.
Vegetative traits measured were limb length (LL), limb width (LWI), plant length (PL), and number of branches from the central taproot (BN) for each plant. Yield characters measured were weight (FWE), length (FL), width (FWI), seeds cavity diameter (SCD), seeds number (SN), and seeds weight (SW). Seed traits analyzed included 100-seeds weight (100-SWE). Measurements on 100- seeds weight were scored using five individuals randomly selected on each plant (Table 3).
Data analysis
Statistical analyses were carried out with STATISTICA software package version 7.1(StatSoft, 2005). Principal Component Analysis (PCA) was applied to identify most discriminant parameters followed by hierarchical cluster analysis using unweighted pair-group method using arithmetic average (UPGMA) method. Multivariate analysis of variance (MANOVA) and a factorial discriminant analysis (FDA) were performed to check difference between the variable means for each group obtained with the clustering analysis. A confusion matrix was constructed, to check the reliability of groups as defined by the hierarchical clustering.
RESULTS
Descriptors variation
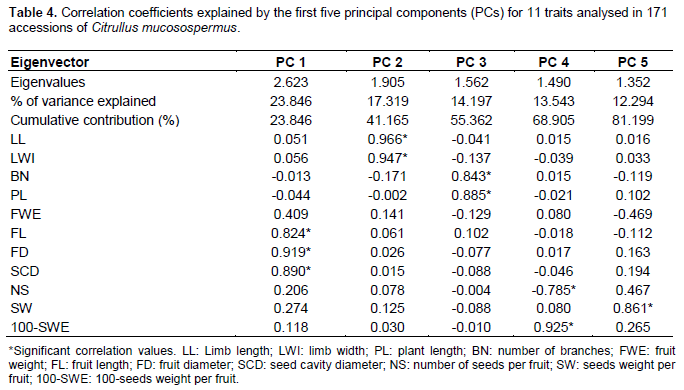
The relative discriminating capacity of the PCA is shown by their Eigenvalues. The result of the PCA (Table 4) showed that five Principal Component axes (PC) had Eigenvalues greater than 1 all together accounted for over 80% of the total variability. The first five principal components accounted for 81.19% of the total variability. The first principal component (PC1) accounted for 23.85%, while the second principal component (PC2) accounted for 17.32%. The PC1 is loaded with fruit length (0.824), fruit diameter (0.919), and seed cavity diameter (0.890). These results indicated that this component was determined by individuals with big fruit. The PC2 was loaded with limb length (0.966), and the limb width (0.947). This component was determined by individuals with broad leaves. PC3, PC4, and PC5 accounted for 14.19, 13.54, and 12.29%. They were correlated to number of branches, and plant length for PC3, number of seeds per fruit and 100-seeds weight per fruit (100-SWE) for PC4 and seeds weight per fruit for PC5. These results indicated that PC3 was determined by individuals of large size plant. PC4 and PC5 were determined by most productive individuals.
Accessions structuration
The unweighted pair group method arithmetic (UPGMA) using a Euclidian distances matrix, subdivided the accessions analyzed into four major groups. These groups differ in the distance by 30 (Figure 1). All groups are mixture of accession of various origin, cultivar or cultigroup.
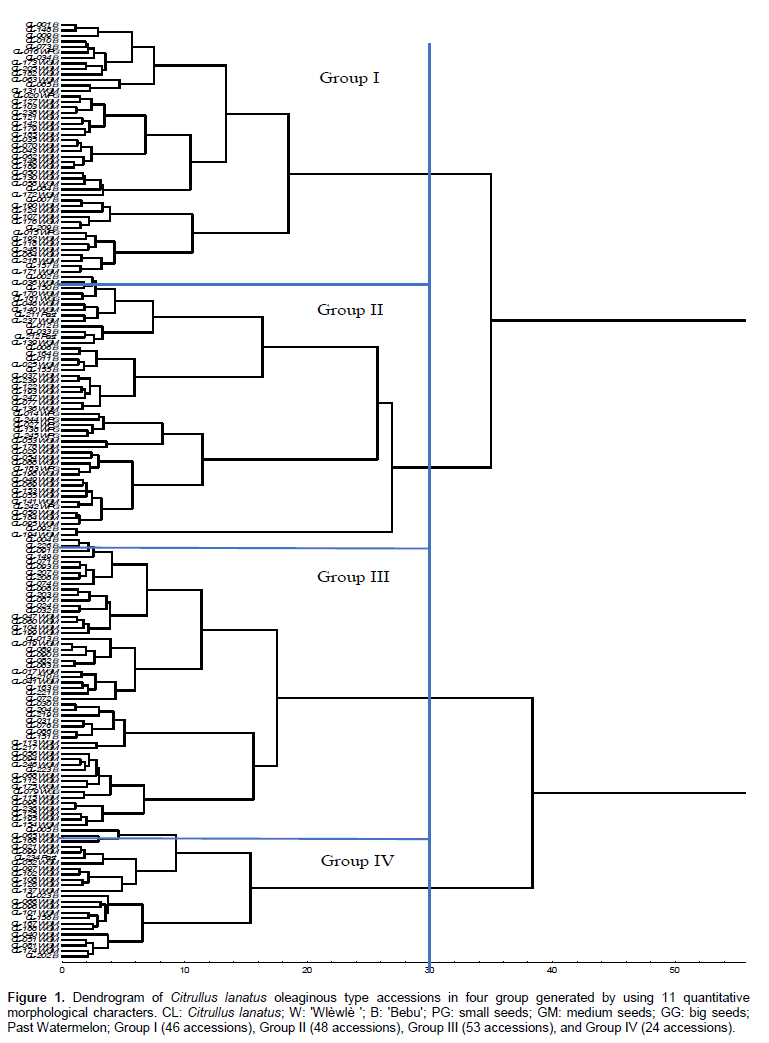
The highest number of accessions was located in cluster III with 53 accessions, followed by clusters II and I with 48 and 46 accessions, respectively. Cluster IV had 24 accessions. Comparison of groups using the MANOVA showed significant differences between the four groups (F= 19.91; P<0.001). This is due to all the 11 characters measured. The ANOVA showed significant difference among groups (Table 5). All parameters revealed a partial distinction of groups.
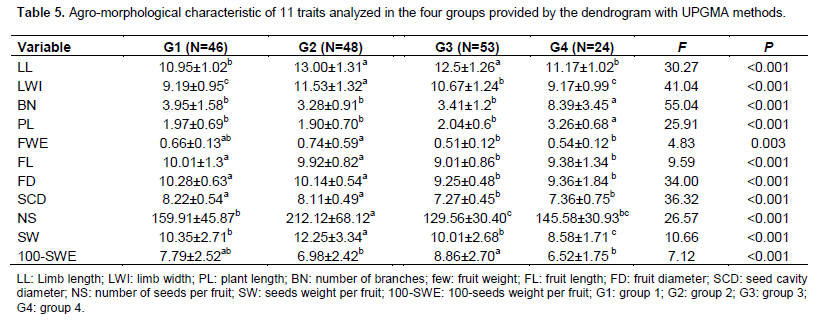
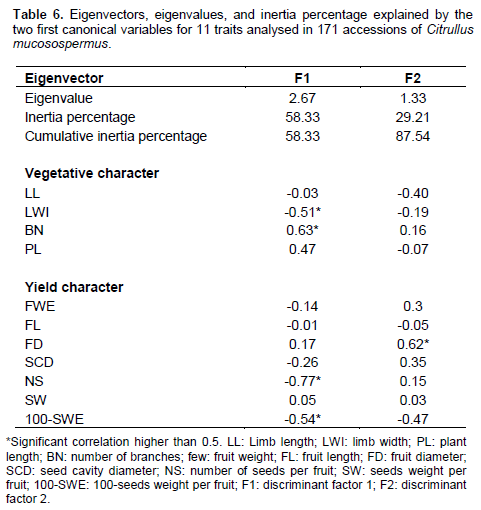
Discriminant functions resulting from variables to classify the units formed on the four groups are shown in Table 6. The two first canonical variables which had an Eigenvalue higher than 1 were retained to describe the total variability of the accessions. The two factors selected together, explained 87.54% of the total variation. The first canonical variable, FDA1, described 58.33% of the total variation. This component was positively correlated with the number of branches and was negatively correlated with limb width, number of seed per fruit, and 100-seeds weight. The second canonical variable, FDA2, explained 29.21% of the total variation and was mostly and positively constructed from fruit diameter. Figure 2 shows the position of the accessions in relation with the first two discriminant factors. Based on these analyses, accessions of group 2 were noted to have the biggest leaves. The smallest leaves were observed in accession of group 1. The most number of the longest plants were observed in group 4. Groups 1 and 2 have the larger and heavier fruit. The best seed yield is obtained with accessions of group 2, but 100 seed-weight was higher in groups 1 and 3.
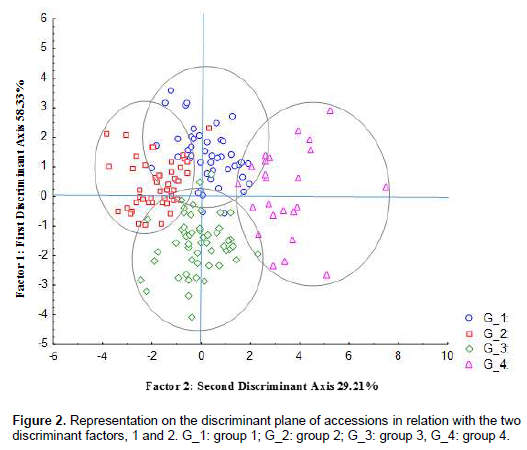
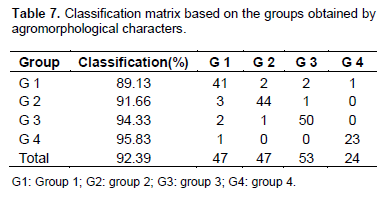
According to confusion matrix, the composition of groups as defined by the hierarchical clustering is different. Group 1 contains 41 accessions instead of 46. Accessions CL-146 and CL-172 have been reclassified in group 2. Similarly, accessions CL-142 and CL-085 have been reclassified in group 3 and accession CL-131 has been reclassified in group 4. Group 2 consists of 44 instead of 48 accessions. An enrollment of 50 accessions belongs to group 3 instead of 53. Group 4 includes 23 accessions instead of 24 (Table 7).
DISCUSSION
To maintain, evaluate and utilize germplasm efficiently, it is important to investigate the extent of genetic diversity available. Morphological characterization is an important step in the description and classification of crop germ-plasm, because a breeding program mainly depends upon the magnitude of morpho-phenological variability (Smith et al., 1991).
In this study, 171 accessions of Citrullus of Nangui University collection were characterized based on morphological traits. In the analysis made to estimate the relative contributions of the different traits studied towards the overall phenotypic variation among the 171 accessions, a total of five principal components (PCs), explained 81.2% of variability. Fruit-related parameters were the most discriminant. In fact, fruit length, fruit diameter, and seed cavity diameter were parameters which contributed more to the first principal component (24%), implying that 24% of the variability was explained by these parameters. These results suggest that fruit size is a useful criterion for separating Citrullus accessions. In the cucurbit family, the significant contribution of the fruit traits to morphological variability has been reported for watermelon (Gusmini and Wehner, 2003; Maggs-Kölling et al., 2000). According to Achigan-Dako (2015) diagnostic morphological markers among species include fruit such as fruit size.
The least discriminant parameters were numbers of seeds and seeds weight. This finding is due to the fact that medium-sized seeds represented 60% of sample analyzed. Contrary to these results, Achigan-Dako et al. (2015) demonstrated that other major differentiation traits is related to seed.
Although our accessions originated from seven different countries, they did not cluster according to geographical distribution. This may be suggestive of the single ancestry of the analyzed plants (Dane and Liu, 2007). This may be because genetically, the cultivated watermelon is closest to plants from West Africa that represents the gene pool from which watermelon was domesticated (Chomicki and Renner, 2014). The sister species of watermelon is C. mucosospermus, the egusi melon, ranging from Nigeria to Senegal (Renner et al., 2014). Plants from Benin closely resemble the collection type of the name C. mucosospermus (which is from Ghana). In order to explain the low genetic diversity of C. mucosospermus (but identified as C. lanatus), Dane and Liu (2007) demonstrated that the watermelon varieties have diverged into small populations. Moreover, the domestication and the resulting selection may have reduced the genetic diversity from the original watermelon crops. Levi et al. (2001) also revealed that cultivated watermelon exhibited narrow genetic base as a results of many years cultivation and selection for specific qualities.
Morphological and genetic characterization of Hungarian and Turkish accessions of watermelon (including C. lanatus and C. colocynthis) revealed that germplasm resources present a wide range of diversity (Sari et al., 2005; Solmaz and Sari, 2009; Solmaz et al., 2010). However, accessions of the two countries show many similarities and therefore cannot be separated clearly (Szamosi et al., 2009). In the present study, comparison of the four groups using a MANOVA combined with factorial discriminant analysis, allowed the classification of accessions according to agro-morphological characters. Accessions of group 2 had the biggest leaves. The smallest leaves were observed in accession of group 1. The largest number of ramification and longest plants were observed in group 4.
Groups 1 and 2 have the largest and heaviest fruits. The best seed yield is obtained with accessions of group 2, but 100 seed-weight was higher in groups 1 and 3 than in the two other groups. Morphological variation analysis in landraces of C. mucosospermus (but identified as C. lanatus by authors in Côte d’Ivoire) also allowed to classify accessions based on fruit and seed traits (Adjoumani et al., 2012).
Although, the Hierarchical Cluster Analysis gave a clear separation of accessions in distinct groups, the confusion matrix displayed a reclassification of some accessions. Accessions CL-146 and CL-172 has been reclassified in group 2. Accessions CL-142 and CL-085 has been reclassified in group 3 and accession CL-131 has been reclassified in group 4.
Traditionally, watermelon breeders focus on fruit quality traits. These traits include sugar content, flesh color, fruit size, and rind patterns (Hashizume et al., 2003). Investigations of the inheritance of fruit morphology and quality traits date back as far at the 1930’s (Porter, 1933, 1937). Since then, many efforts have been made to better understand traits associated with watermelon fruit quality and morphology. Many genes have been described that control the internal fruit quality and morphology in watermelon (Guner and Wehner, 2004). An internal fruit characteristic that has received attention is the Brix value.
A linkage map was developed and a QTL that accounts for 19% of the variation in Brix was mapped on linkage group (LG) 8 (Prothro, 2010). Fruit shape and weight are important external characteristics that breeders must consider when developing watermelon cultivars.
Watermelon shape can be classified as being either round, oval, blocky or elongate (Wehner et al., 2001). The only gene described that controls fruit shape is the elongate fruit gene (O). Fruit weight has recently become an important consideration for breeders due to increased consumer preference for smaller sized watermelons (Gusmini and Wehner, 2007). Watermelons have traditionally been classified into five categories based on fruit weight. These are icebox (less than 5.5 kg), small (5.5 to 8.0 kg), medium (8.1 to 11.0 kg), large (11.1 to 14.5 kg), giant (greater than 14.5 kg), and recently mini fruit size (1.5 to 4.0 kg) was added (Gusmini and Wehner, 2007; Maynard, 2001). No genes have been identified that are responsible for fruit weight inwatermelon (Gusmini and Wehner, 2007; Maynard, 2001). Fruit quality and morphological traits, such as the ones described earlier, gain the most attention from watermelon breeders. There are many other important traits in watermelon that receive little to no attention. The egusi seed trait is one of these traits. The egusi seed trait has been shown to be inherited as a single gene (Gusmini et al., 2004).
A single QTL was identified for rind thickness (Prothro, 2010). The objective when breeding for this trait should be to have a rind thickness that is a small percentage of the fruit diameter. Three QTL were identified for fruit length and two QTL were identified for fruit width, in the same region on LG5. Another QTL for fruit length was identified on LG3. But it is not known if they are associated with the elongate fruit gene. Kumar et al. (2009) produced a population segregating for fruit shape and found that his data did not fit the single gene theory for fruit shape inheritance. Watermelons varieties that produce fruit that fit into the weight category most preferred by consumers must be available for growers(Gusmini and Wehner, 2007). The QTL identified for fruit length and fruit width should be useful for fruit weight selection in a breeding program. Two QTL were identified for this trait on LG2. Another QTL was identified for seed oil percentage. This QTL falls between the locations of the two QTL identified for the egusi seed trait on LG2 (Prothro et al., 2012).
CONCLUSION
A great deal of diversity morphological traits has been observed in C. mucosospermus oleaginous type accessions from the Nangui Abrogoua University collection. From the present investigation, it was concluded that the indigenous oleaginous cucurbit C. mucosospermus displayed a wide range of diversity for most of the morphological traits studied. Traits related to fruit size and weight were the most accession distinctive. Consequently, fruit morphological traits could be used for accession identification during collecting missions. Accessions had been divided into four groups in which group 2 had the highest seed yield. Therefore, accessions of this group must be used to enhance yield. These ï¬ndings are important to plan future germplasm sampling and evaluation studies.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Achigan-Dako EG, Avohou ES, Linsoussi C, Ahanchede A, Vodouhe RS, Blattner FR (2015). Phenetic characterization of Citrullus spp. (Cucurbitaceae) and differentiation of egusi-type (C. mucosospermus). Genet. Resour. Crop. Evol. 62(8):1159-1179. |
|
|
Achigan Dako EG, Fagbemissi R, Avohou HT, Vodouhe RS, Coulibaly O, Ahanchede A (2008). Importance and practices of egusi crops (Citrullus lanatus (Thunb.) Matsum. & Nakai, Cucumeropsis mannii Naudin and Lagenaria siceraria (Molina) Standl. cv.'Aklamkpa') in sociolinguistic areas in Benin. Biotechnol. Agron. Soc. Environ. 12(4):393-403. |
|
|
Adjoumani K, Kouonon LC, Akaffou DS, Dje Y (2012). Diversité variétale chez l'espèce Citrullus lanatus (Matsumura et Nakai) et opportunités d'amélioration génétique des cultivars. Eur. J. Sci. Res. 67(4):564-579. |
|
|
Bailey LH (1930). Three discussions in Cucurbitaceae. Gentes Herb. 2:175-186. |
|
|
Burkill HM (1985). The useful plants of West Tropical Africa. Richmond, Surrey, UK: Royal Botanic Gardens, Kew. |
|
|
Che K-p, Liang C-y, Wang Y-g, Jin D-m, Wang B (2003). Genetic assessment of watermelon germplasm using the AFLP technique. HortScience 38(1):81-84. |
|
|
Chomicki G, Renner SS (2014). Watermelon origin solved with molecular phylogenetics including Linnaean material: another example of museomics. New Phytol. 205: 526-532. |
|
|
Dane F, Liu J (2007). Diversity and origin of cultivated and citron type watermelon (Citrullus lanatus). Genet. Resour. Crop. Evol. 54:1255-1265. |
|
|
Djè Y, Kouonon CL, Zoro Bi IA, Gnamien YG, Baudoin JP (2006). Etude des caractéristiques botaniques, agronomiques et de la biologie florale du melon africain (Cucumis melo L. var. agrestis Naudin, Cucurbitaceae). Biotechnol. Agron. Soc. Environ. 10(2):109-119. |
|
|
Fursa TB (1972). On the taxonomy of the genus Citrullus Schad. Bot. Zh. 57:31-34. |
|
|
Fursa TB (1981). Intraspecific classification of watermelon under cultivation. Kulturpflanze 29: 297-300. |
|
|
Fursa TB (1983). Novyi vid arbuza Citrullus mucosospermus (Fursa) Fursa (A new species of watermelon Citrullus mucosospermus (Fursa) Fursa.) Trudy Prikl. Bot. Genet. Selek. 81:108-112. |
|
|
Guner N, Wehner TC (2004). The genes of watermelon. HortScience 39:1175-1182. |
|
|
Gusmini G, Wehner TC (2003). Polygenic inheritance of some vine traits in two segregating watermelon families. Cucurbit Genet. Coop. Rep. 26:32-35. |
|
|
Gusmini G, Wehner TC (2007). Heritability and genetic variance estimates for fruit weight in watermelon. HortScience 42:1332-1336. |
|
|
Gusmini G, Wehner TC, Jarret RL (2004). Inheritance of Egusi Seed Type in Watermelon. Journal of Heredity 95(3):268-270. |
|
|
Hammer K, Gladis T (2014). Notes on infraspecific nomenclature and classifications of cultivated plants in Compositae, Cruciferae, Cucurbitaceae, Gramineae (with a remark on Triticum dicoccon Schrank) and Leguminosae. Genet. Resour. Crop. Evol. 61:1555-1567. |
|
|
Hashizume T, Shimamoto I, Hirai M (2003). Construction of a linkage map and QTL analysis of horticultural traits for watermelon [Citrullus lanatus (THUNB.) MATSUM & NAKAI] using RAPD, RFLP and ISSR markers. Theor. Appl. Genet. 106:779-785. |
|
|
Hepper FN (1990). Pharaoh's flowers: the botanical treasures of Tutankhamun. Chicago, IL, USA & London, UK: HMSO. |
|
|
Hutchinson J, Dalziel JM (1954). Flora of West Tropical Africa: the British West African Territories, Liberia, the French and Portugese |
|
|
Koffi KK, Gbotto AA, Malice M, Djè Y, Bertin P, Baudoin JP, Zoro Bi IA (2008). Morphological and allozyme variation in a collection of Cucumeropsis mannii Naudin (Cucurbitaceae) from Côte d'Ivoire. Biochem. Syst. Ecol. 36: 777-789. |
|
|
Kouassi NJ, Zoro Bi IA (2010). Effect of sowing density and seedbed type on yield and yield components in Bambara Groundnut (Vigna subterranea) in woodland savannas of Côte d'Ivoire. Exp. Agric. 46:99-110. |
|
|
Kumar RS, Parthiban KT, Rao GM (2009). Molecular characterization of Jatropha genetic resources through inter-simple sequence repeat (ISSR) markers. Mol. Biol. Rep. 36:1951-1956. |
|
|
Levi A, Thomas CE, Wehner TC, Zhang X (2001). Low genetic diversity indicates the need to broaden the genetic base of cultivated watermelon. HortScience 36(6):1096-1101. |
|
|
Loukou AL, Gnakri D, Djè Y, Kippré AV, Malice M, Baudoin JP, Zoro Bi IA (2007). Macronutrient composition of three cucurbit species cultivated for seed consumption in Côte d'Ivoire. Afr. J. Biotechnol. 6(5):529-533. |
|
|
Maggs-Kölling GL, Madsen S, Christiansen JL (2000). A phenetic analysis of morphological variation in Citrullus lanatus in Namibia. Genet. Resour. Crop. Evol. 47:385-393. |
|
|
Marr KL, Xia Y-M, Bhattarai NK (2007). Allozymic, morphological, phenological, linguistic, plant use, and nutritional data of Benincasa hispida (Cucurbitaceae). Econ. Bot. 61:44-59. |
|
|
Maynard DN (2001). An introduction to the watermelon. Alexandria, VA, USA: ASHS Press. |
|
|
Meeuse AD (1962). The Cucurbitaceae of Southern Africa. Bothalia 8: 1-111. |
|
|
Morimoto Y, Maundu P, Fujimaki H, Morishima H (2005). Diversity of landraces of the white-flowered gourd (Lagenaria siceraria) and its wild relatives in Kenya: fruit and seed morphology. Genet. Resour. Crop. Evol. 52:737-747. |
|
|
Ndabalishye I (1995). Agriculture vivrière Ouestafricaine à travers le cas de la Côte d'Ivoire: monographie. IDESSA, Bouaké, Côte d'Ivoire. |
|
|
Oyulu C (1977). A quantitative and qualitative study of seed types in egusi (Citrullus colocynthis L). Trop. Sci. 19:55-62. |
|
|
Porter DR (1933). Watermelon breeding. Hilgardia 7: 533-552. |
|
|
Porter DR (1937). Inheritance of certain fruit and seed characters in watermelons. Hilgardia, pp. 489-509. |
|
|
Prothro J, Sandlin K, Abdel-Haleem H, Bachlava E, White V, Knapp S, McGregor C (2012). Main and epistatic quantitative trait loci associated with seed size in watermelon. J. Am. Soc. Hortic. Sci. 137(6): 452-457. |
|
|
Prothro JM (2010). Genetic mapping of phenotypic and quantitatibe trait loci underlyinf horticulturaly important traits in watermelon, The University of Georgia, Athens, Georgia. |
|
|
Renner SS, Chomicki G, Greuter W (2014). (2313) Proposal to conserve the name Momordica lanata (Citrullus lanatus) (watermelon, Cucurbitaceae), with a conserved type, against Citrullus battich. Taxon 63(4):941-942. |
|
|
Sari N, Solmaz I, Yetisir H, Unlu H (2005). Watermelon genetic resources in Turkey and their characteristics. III International symposium on cucurbits 731:433-438. |
|
|
Schaefer H, Renner SS (2011). Phylogenetic relationships in the order Cucurbitales and a new classiï¬cation of the gourd family (Cucurbitaceae). Taxon 60:122-138. |
|
|
Schulz E (1987). Die Holozäne Vegetation der Zentralen Sahara (N-Mali, N-Niger, SW-Libyen). In Palaeoecology of Africa and the surrounding Islands, 18:143-161. |
|
|
Schulz E (1991). Holocene environments in the central Sahara. Hydrobiologia 214:359-365. |
|
|
Smith SE, Doss AA, Warburton M (1991). Morphological and agronomic variation in North African and Arabian alfalfas. Crop Sci. 31:1159-1163. |
|
|
Solmaz I, Sari N (2009). Characterization of watermelon (Citrullus lanatus) accessions collected from Turkey for morphological traits. Genet. Resour. Crop. Evol. 56:173-188. |
|
|
Solmaz I, Sari N, Aka-Kacar Y, Yalcin-Mendi YN (2010). The genetic characterization of Turkish watermelon (Citrullus lanatus) accessions using RAPD markers. Genet. Resour. Crop. Evol. 57:763-771. |
|
|
StatSoft (2005).STATISTICA, logiciel d'analyse de données. www.statsoft.fr. |
|
|
Szamosi C, Solmaz I, Sari N, Bársony C (2009). Morphological characterization of Hungarian and Turkish watermelon (Citrullus lanatus (Thunb.) Matsum. et Nakai) genetic resources. Genet. Resour. Crop. Evol. 56:1091-1105. |
|
|
van der Vossen HAM, Denton OA, El Tahir IM (2004).Citrullus lanatus (Thunb.) Matsum. & Nakai. In PROTA (Plant Resources of Tropical Africa / Ressources végétales de l'Afrique tropicale) (Eds Grubben GJH and Denton OA). Wageningen, Netherlands. |
|
|
Wehner TC, Shetty, N.V., Elmstron GW (2001). Breeding and seed production. In Watermelons: Characteristics, production, and marketing, 27-73 (Ed Maynard DN). Alexandria, VA: ASHS Press. |
|
|
Whitaker TW, Bemis WB (1976). Cucurbits. In Evolution of crop plants, pp. 64-69. |
|
|
Zoro Bi IA, Koffi KK, Djè Y (2003). Caractérisation botanique et agronomique de trois espèces de cucurbites consommées en sauce en Afrique de l'Ouest: Citrullus sp.,Cucumeropsis mannii Naudin et Lagenaria siceraria (Molina) Standl. Biotechnol. Agron. Soc. Environ. 7(3-4):189-199. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


