Full Length Research Paper

ABSTRACT
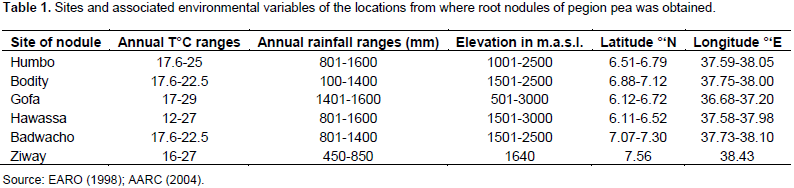
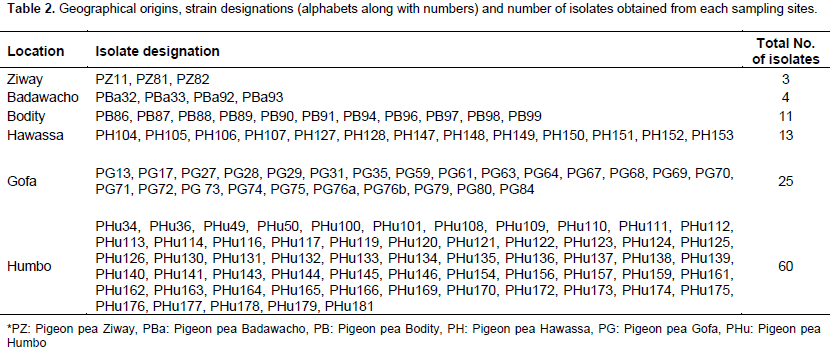
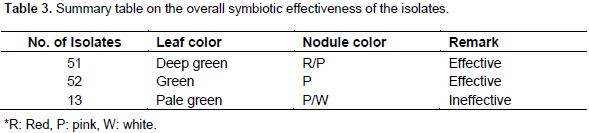
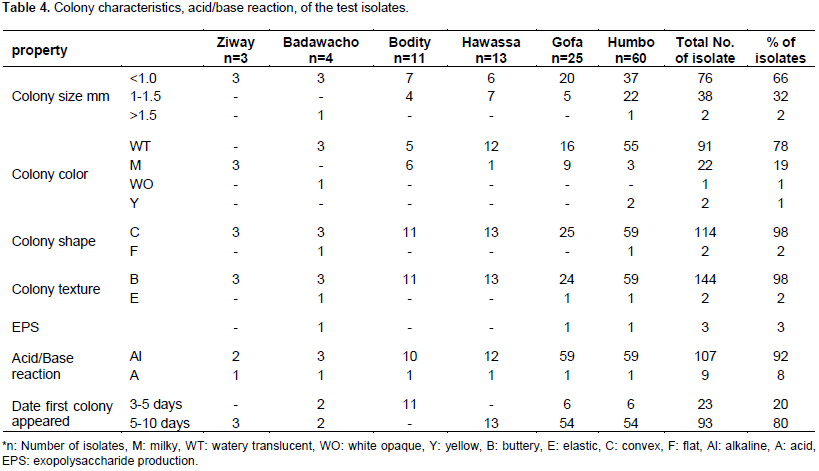
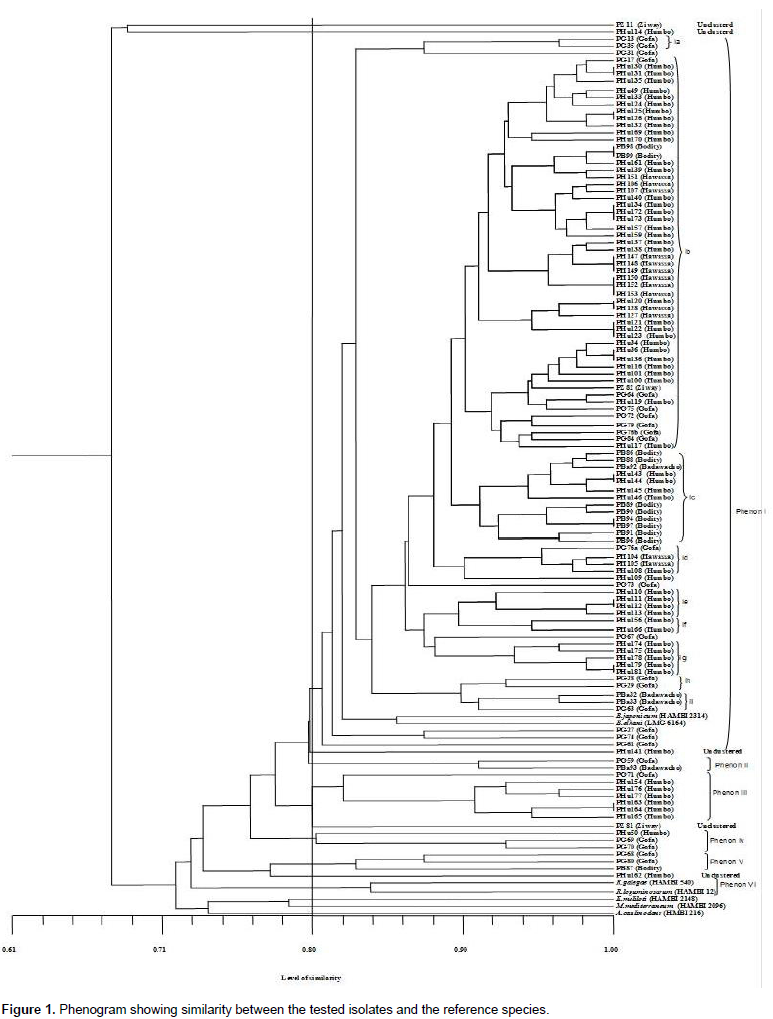
Pigeon pea (Cajanus cajan (L.) Millsp.) is an important protein source grown in several tropical and sub-tropical countries, and is considered a multi-purpose plant that is resistant to the conditions where drought and salinity is a common phenomenon. The aim of this study was to evaluate the diversity of rhizobial isolates obtained from root nodules of pigeon pea plants grown in central and southern Ethiopia. A total of 116 nitrogen-fixing rhizobial strains were isolated. The bacterial isolates were characterized by 91 phenotypic traits including cultural characteristics, intrinsic antibiotic and heavy metal resistance, salt, pH and incubation temperature tolerance, and carbon and nitrogen sources utilization ability. Preliminary symbiotic properties of the isolates were also evaluated. The isolates were compared with seven reference strains of rhizobia by application of the unweighted pair group method with arithmetic means (UPGMA) using NTSYSpc Version 2.1 software program. The dendrogram constructed from cluster analysis of 91 phenotypic traits, grouped them into six clusters and eight un-clustered positions at 80% relative similarity. Cluster I contained 83% of the test isolates that were grouped together with the reference strains Bradyrhizobium japonicum (HAMBI 2314T) and Bradyrhizobium elkanii (LMG 6164), suggesting that pigeon pea is commonly nodulated by bradyrhizobia. Results from symbiotic effectiveness test revealed that majority of the isolates were found to be effective. Generally, this investigation demonstrated that rhizobial population nodulating C. cajan on the study area were phenotypically diverse and symbiotically effective. Furthermore, the result indicates the existence of strains in the collection, which can tolerate environmental stresses, thus can be developed into inoculant for pigeon pea inoculation and production in Ethiopia and beyond.
Key words: Bradyrhizobium, Cajanus cajan, phenotypic characteristics, rhizobium.
INTRODUCTION
MATERIALS AND METHODS
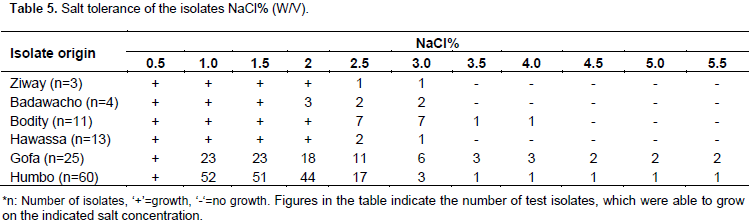
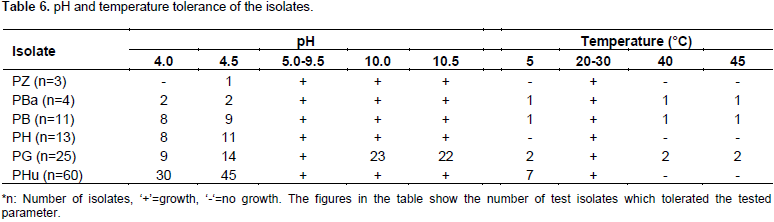
RESULTS AND DISCUSSION
CONFLICT OF INTERESTS
REFERENCES
|
Alikhani HA, Saleh-Rastin N, Antoun H (2006). Phosphate solubilization activity of rhizobia native to Iranian soils. Plant Soil. 287:35-41. |
|
|
Amarger N, Macheret V, Laguerre G (1997). Rhizobium gallicum sp. nov. and Rhizobium giardinii sp. nov., from Phaseolus vulgaris nodules. Int. J. Syst. Evol. Microbiol. 47:996-1006. |
|
|
Anand RC, Dogra BC (1991). Physiological and biochemical characteristics of fastâ€and slowâ€growing Rhizobium sp. pigeon pea (Cajanus cajan). J. Appl. Microbiol. 70:197-202. |
|
|
Beltrame TP, Rodrigues E (2007). Guandu bean (Cajanus cajan (L.) Millsp.) on tropical forest restoration. Semina: Ciências Agrárias 28:19-28. |
|
|
Beyene D, Kassa S, Ampy F, Asseffa A, Gebremedhin T, Van Berkum P (2004). Ethiopian soils harbor natural populations of rhizobia that form symbioses with common bean (Phaseolus vulgaris L.). Arch. Microbiol. 181:129-136. |
|
|
Bordeleau LM, Prévost D (1994). Nodulation and nitrogen fixation in extreme environments. Plant Soil. 161:115-125. |
|
|
Bouhmouch I, Brhada F, Filali-Maltou A, Aurag J (2001). Selection of osmotolerant and effective strains of Rhizobiaceae for inoculation of common bean (Phaseolus vulgaris) in Moroccan saline soils. Agronomie 21:591-599. |
|
|
Chen L, Figueredo A, Villani H, Michajluk J, Hungria M (2002). Diversity and symbiotic effectiveness of rhizobia isolated from field-grown soybean nodules in Paraguay. Biol. Fertil. Soils 35:448-457. |
|
|
Coutinho HLC, Oliveira VM, Lovato A, Maia AHN, Manfio GP (1999). Evaluation of the diversity of rhizobia in Brazilian agricultural soils cultivated with soybeans. Appl. Soil. Ecol. 13:159-167. |
|
|
Degefu T, Wolde-Meskel E, Liu B, Cleenwerck I, Willems A, Frostegard A (2013). Mesorhizobium shonense sp. nov., Mesorhizobium hawassense sp. nov. and Mesorhizobium abyssinicae sp. nov., isolated from root nodules of different agroforestry legume trees. Int. J. Syst. Evol. Microbiol. 63:1746-53. |
|
|
Eaglesham ARJ (1987). The use of intrinsic antibiotic resistance for Rhizobium study. Available at: |
|
|
Elkan GH (1992). Taxonomy of the rhizobia. Can. J. Microbiol. 38:446-450. |
|
|
Fernandes JPI, Lima AAd, Passos SR, Gava CAT, Oliveira PJd, Rumjanek NG, Xavier GR (2012). Phenotypic diversity and amylolytic activity of fast growing rhizobia from pigeon pea [Cajanus cajan (L.) Millsp.]. Braz. J. Microbiol. 43:1604-1612. |
|
|
Hancock JF (2012). "Plant evolution and the origin of crop species," CABI. |
|
|
Hernandez BS, Focht DD (1984). Invalidity of the concept of slow growth and alkali production in cowpea rhizobia. Appl. Environ. Microbiol. 48: 206-210. |
|
|
Hungria M, Vargas MAT (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 65:151-164. |
|
|
Jordan DC (1982). Transfer of Rhizobium japonicum Buchanan 1980 to Bradyrhizobium gen. nov., a Genus of Slow-Growing, Root Nodule Bacteria from Leguminous Plants. Int. J. Syst. Bacteriol. 32:136-139. |
|
|
Jordan DC (1984). Family III. Rhizobiaceae. Bergey's Manual of Systematic Bacteriology 1:234-242. |
|
|
Júnior PIF, De Oliveira PJ, Rumjanek NG, Xavier GR (2011). Poly-β-hydroxybutyrate and exopolysaccharide biosynthesis by bacterial isolates from pigeon pea [Cajanus cajan (L.) Millsp] root nodules. Appl. Biochem. Biotechnol. 163:473-484. |
|
|
Kalita M, Małek W (2004). Phenotypic and genomic characteristics of rhizobia isolated from Genista tinctoria root nodules. Syst. Appl. Microbiol. 27:707-715. |
|
|
Kennedy LD, Greenwood RM (1982). 6-Phosphogluconate and glucose-6-phosphate dehydrogenase activities, growth rate and acid production as taxonomic criteria for Rhizobium. NZL. J. Sci. |
|
|
Kishinevsky BD, Nandasena KG, Yates RJ, Nemas C, Howieson JG (2003). Phenotypic and genetic diversity among rhizobia isolated from three Hedysarum species: H. spinosissimum, H. coronarium and H. flexuosum. Plant Soil. 251:143-153. |
|
|
Lie TA, Göktan D, Engin M, Pijnenborg J, Anlarsal E (1987). Co-evolution of the legume-Rhizobium association. In "Plant and Soil Interfaces and Interactions", pp. 171-181. Springer. |
|
|
Lindström K, Lehtomäki S (1988). Metabolic properties, maximum growth temperature and phage sensitivity of Rhizobium sp.(Galega) compared with other fast-growing rhizobia. FEMS Microbiol. Lett. 50:277-287. |
|
|
Moreira FMS, Gillis M, Pot B, Kersters K, Franco AA (1993). Characterization of rhizobia isolated from different divergence groups of tropical Leguminosae by comparative polyacrylamide gel electrophoresis of their total proteins. Syst. Appl. Microbiol. 16:135-146. |
|
|
Odee DW, Sutherland JM, Makatiani ET, McInroy SG, Sprent JI (1997). Phenotypic characteristics and composition of rhizobia associated with woody legumes growing in diverse Kenyan conditions. Plant Soil. 188:65-75. |
|
|
Peoples MB, Herridge DF, Ladha JK (1995). Biological nitrogen fixation: an efficient source of nitrogen for sustainable agricultural production? Plant Soil. 174:3-28. |
|
|
Ramsubhag A, Umaharan P, Donawa A (2002). Partial 16S rRNA gene sequence diversity and numerical taxonomy of slow growing pigeon pea (Cajanus cajan L. Millsp) nodulating rhizobia. FEMS Microbiol. lett. 216: 139-144. |
|
|
Singleton PW, AbdelMagid HM, Tavares JW (1985). Effect of phosphorus on the effectiveness of strains of Rhizobium japonicum. Soil Sci. Soc. Am. J. 49:613-616. |
|
|
Somasegaran P, Hoben H (1994). Methods in Legume-rhizobium technology; NIFTAL project. Springer-Verlag, Heidelberg, Germany. |
|
|
Surange S, Wollum Ii AG, Kumar N, Nautiyal CS (1997). Characterization of Rhizobium from root nodules of leguminous trees growing in alkaline soils. Can. J. Microbiol. 43:891-894. |
|
|
Swelim DM., Hashem FM, Kuykendall LD, Hegazi NI, Abdel-Wahab SM (1997). Host specificity and phenotypic diversity of Rhizobium strains nodulating Leucaena, Acacia, and Sesbania in Egypt. Biol. Fertil. Soils. 25:224-232. |
|
|
Tena W, Wolde-Meskel E, Degefu T, Walley F (2017a). Genetic and phenotypic diversity of rhizobia nodulating Chickpea (Cicer arietinum L.) in soils from southern and central Ethiopia. Can. J. Microbiol. 63:690-707. |
|
|
Tena W, Wolde-Meskel E, Degefu T, Walley F (2017b). Lentil (Lens culinaris Medik.) nodulates with genotypically and phenotypically diverse rhizobia in Ethiopian soils. Syst. Appl. Microbiol. 40:22-33. |
|
|
Vincent JM (1970). A manual for the practical study of the root-nodule bacteria. A manual for the practical study of the root-nodule bacteria. |
|
|
Wolde-Meskel E, Terefework Z, Frostegard A, Lindstrom K (2005). Genetic diversity and phylogeny of rhizobia isolated from agroforestry legume species in southern Ethiopia. Int. J. Syst. Evol. Microbiol. 55:1439-52. |
|
|
Wolde-meskel E, Terefework Z, Lindström K, Frostegård Å (2004). Metabolic and genomic diversity of rhizobia isolated from field standing native and exotic woody legumes in southern Ethiopia. Syst. Appl. Microbiol. 27:603-611. |
|
|
Workalemahu A, Assefa F (2007). Phenotypic characteristics of common bean (Phaseolus vulgaris)-nodulating rhizobia from some parts of southern Ethiopia. Ethiop. J. Biol. Sci. 6:97-114. |
|
|
Zhang X, Harper R, Karsisto M, Lindström K (1991). Diversity of Rhizobium bacteria isolated from the root nodules of leguminous trees. Int. J. Syst. Evol. Microbiol. 41:104-113. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


