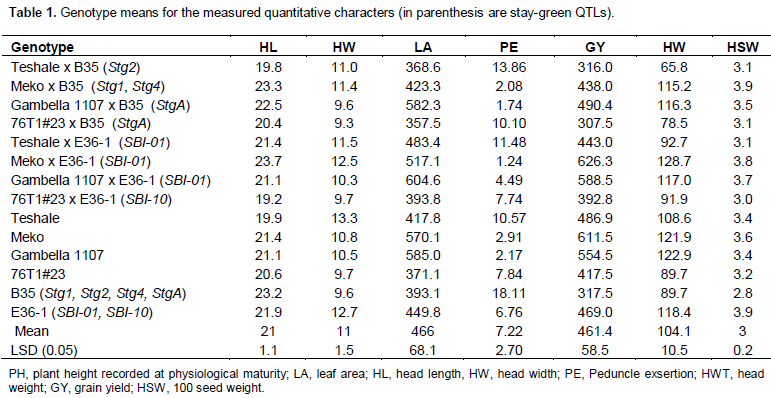
Full Length Research Paper

ABSTRACT
INTRODUCTION
Drought is the single greatest problem limiting crop productivity in the semi-arid tropics (SAT) where most of the production systems are rain-fed (Serraj et al., 2005). The trade-offs between the trends of increased population size and the associated increased demand for food on one hand and the decrease in availability of water on the other hand call for the development of more water efficient (climate smart) crops (Balota et al., 2008). Sorghum is one of the most important food, feed and bio-energy crops in the world, which tolerates drought stress better than most other cereals. It is also an important food crop in sub-Sahara Africa including Ethiopia on which millions of poor people depend. In Ethiopia, it grows in a wide range of agro-ecological zones, but it is more important in the dry lowlands where crop failure due to recurrent drought stress is common (Adugna, 2007). Cates (1922) has applied the name "crop camel" to kafir (one of the grain sorghums) because the plants can stand for considerable dry periods without apparent suffering from a deficiency of moisture. Since the crop is both drought tolerant and highly responsive to added water, it is adapted to both dryland and irrigated conditions (Eck and Musick, 1979).
Drought may occur at any stage during the growth cycle of the sorghum crop. Sorghum exhibits two distinct responses to drought stress (Rosenow et al., 1983). One occurs when plants are stressed during the head development stage prior to flowering, called preflowering, and the second occurs when plants are stressed after anthesis and during grain development, called post-flowering (Walulu et al., 1994). When it occurs as preflowering during the first growth stage (GS1), the period between seedling emergence and panicle initiation (Quinby, 1974), it affects the vegetative growth and the time of floral initiation. Drought at second growth stage (GS2), the period between panicle initiation and anthesis, affects the number of seeds that will be produced, whereas at the third growth stage (GS3), the period between floral anthesis and physiological maturity, it affects seed weight as the grain does not fill well. Therefore, grain yield reduction becomes relatively worse when drought occurs at the last two growth stages (GS2-pre flowering and GS3-post-flowering) than at GS1 since seed number and weight are the most important grain yield components in sorghum. The reason why terminal drought stress might cause yield reduction is that, the affected plants mature early as a result of the induction of premature leaf senescence (forced maturity) (Gregersen et al., 2013). Genetic variation for resistance to drought at each stage has been observed in sorghum germplasm (Walulu et al., 1994). Hence, many genotypes with a high level of resistance at one stage were found to be susceptible at the other stage (Rosenow et al., 1983). Until recently, the global sorghum improvement approach towards alleviating the effect of drought has been focused on the development of short duration varieties, those that can escape drought occurs late during the season (terminal drought). Based on this approach a number of early maturing varieties have been released and are still being produced globally including Ethiopia. However, even these early maturing types can fail to produce if drought occurs at the critical stages. For example, in pearl millet, Mahalakshmi et al. (1987) found that when drought occurred during the midseason, early genotypes, which were proved to effectively escape a terminal drought stress suffered greater grain yield reduction than the later flowering genotypes.
Recently, the focus of research has been changed to the development of drought tolerant varieties helped by marker assisted selection. The best example of marker assisted selection against drought in sorghum is for the stay-green trait (Rosenow et al., 1983). The term stay-green refers to delayed senescence, which is associated with post-flowering drought tolerance irrespective of the maturity of the genotype (Ejeta and Knoll, 2007). It focuses mainly on alleviating the effect of drought at grain filling as drought stress during this stage in sorghum usually results in rapid premature plant senescence (Stout and Simpson, 1978). Sorghum genotypes with functional stay-green continue to fill their grain normally under drought (Rosenow and Clark, 1981; Borrell et al., 2014). Such stay green trait, producing healthier stems on plants, may indirectly contribute to higher crop performance by providing disease resistance, standability, resistance to lodging, and higher seed quality characteristics (Cukadar-Olmedo and Miller, 1997; Cattivelli et al., 2008).
Moreover, previous reports suggest that the Stay-Green trait in sorghum and rice might contribute to increased post-anthesis biomass production under drought stress (Borrell et al., 2000; 2001). Therefore, stay-green has been suggested as an indirect selection criterion for post-flowering drought tolerance (Rosenow et al., 1983). Molecular markers linked to the already mapped sorghum stay green QTLs are available (Hash et al., 2003; Harris et al., 2007) and with which marker assisted backcrossing has been undertaken (Kassahun et al., 2010). This study was, thus intended to evaluate the response of stay-green QTL introgression sorghum lines developed using marker assisted backcrossing and their parents for induced post-anthesis drought stress.
MATERIALS AND METHODS
Sorghum varieties adapted to the dry lowlands of Ethiopia; 76T1#23, Teshale, Gambella 1107, and Meko were selected for the backcrossing program. All of them, except Gambella 1107 were selections from ICRISAT introduced regional trials. Gambella 1107 is a selection from landraces in Gambella area, Ethiopia. These varieties have maturity days ranging from 90 to 130 days. So far, the sources of the stay-green QTLs that are widely used in sorghum have been B35 and E36-1 lines, which were originally obtained from Ethiopia (Reddy et al., 2009). B35 is a BC1 derivative of landrace germplasm accession IS 12555, which is a durra sorghum (Rosenow et al., 1983). A Marker assisted backcrossing program was held in Ethiopia between 2006 and 2008 and carried out at Melkassa Agricultural Research Center in collaboration with ICRISAT, Nairobi to introgress stay-green QTLs from B35 and E36-1 to the locally adapted early maturing, but senescent sorghum varieties (76T1#23, Teshale, Gambella 1107, and Meko). Starting from the first backcrossing, all plants in a 5 m nursery plot were tagged and leaf samples were taken from each of them and sent to the Bio-Sciences for East and Central Africa (BECA) laboratory in Nairobi, Kenya for marker assisted selection of the stay green QTLs. Based on the marker information, those plants which had one or more stay-green QTLs from the donor parents were backcrossed to the respective locally adapted lowland sorghum varieties used as recurrent parents. In total, six stay-green QTLs conferring post-anthesis drought stress tolerance: StgA, Stg1, Stg2, and Stg4 from B35 and SBI-01 and SBI-10 from E36-1 were introgressed into the four elite local varieties in three molecular marker (SSRs) assisted backcrosses and the resulting populations were self-pollinated three times. Thus, all of the introgression lines used in this study had one or more of the stay-green quantitative trait loci (QTLs) (Table 1). Hence, eight backcross populations (BC3F3) here after referred to as stay-green QTL introgression lines resulted from the four lowland sorghum varieties and the two stay green source lines (B35 and E36-1) were included in the experiment. Moreover, the 6 parents (donor and recurrent) were included in the experiment for comparison (Table 1).
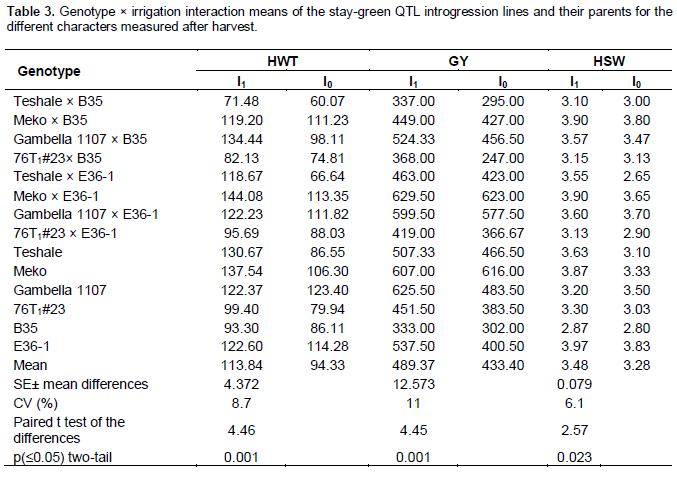
The field experiment was carried out during the off-season (from 16 December 2009 through 24 April 2010) at Melkassa Agricultural Research Center located in the Central Rift Valley of Ethiopia (39°21′E, 8°24′N, altitude=1550 m) by inducing drought at the post-anthesis stage. The soil at the experimental site is silty clay loam Andosol with a pH of ~7.8. It was a split plot design with three replications. The main plot factor was irrigation with two levels (I0 = irrigation withheld after anthesis and I1 = irrigation continued after anthesis until maturity) and the sub-plot factor was the genotypes with 14 levels (consists of 8 introgression lines, 2 donor, and 4 recurrent parents). The two groups of the irrigation (flood) treatments differed in three frequencies during the post-flowering stage. In the first case (I1), all of the 14 genotypes were irrigated every seven days throughout their growth stages. In the second (I0), however, the replicas of all of the genotypes did not receive irrigation after anthesis (the last irrigation was given just at anthesis). Hence, the total number of treatments was 2 × 14 = 28. The border of each block was sown to the variety Melkam to avoid border effect. Each plot had a single row of 4 m length and the inter-row space was 75 cm. the space between the two main plots was 1.5 m. The treatments were randomized within each main plot. The seeds were sown by hand drilling and later thinned to 15 cm between plants to give a total population density of 88 888 plants per hectare. The management practices including fertilizer and weeding except irrigation were done as per the recommendation for sorghum. Accordingly, DAP fertilizer was applied at the rate of 100 Kg/ha during planting in the seed furrow and Urea was applied at the rate of 50 Kg/ha as top dressing before booting. The field was kept free from weeds throughout the experiment. To control shoot flies and stem borers, Karate 5% EC was applied at the rate of 320 mm/ha two weeks after emergence.
As there is heavy weaver bird (Quelea quelea) pressure at Melkassa during the off-season, it was difficult to measure plot yield. Therefore, five plants were bagged with cloth bags from each row plot after anthesis and used for data recording. Data on days to flowering (DTF), plant height (PH) recorded at physiological maturity, leaf area (LA), head length (HL), head width (HW), Peduncle exsertion (PE), head weight (HWT), grain yield (GY) (Kg per plot, in this case mean of 5 plants), and 100 seed weight (HSW) at 12% moisture content, were measured from the five plants based on the new key access and utilization of sorghum descriptors (Bioversity International, 2010). In addition, the visual leaf senescence data scoring was modified from the descriptors for sorghum (IBPGR/ICRISAT, 1993) as 0 = ~100% green leaf area; 1=75% green leaf area; 2= 50% green leaf area; 3= 25% green leaf area; 4 = 100% leaves and stalk dead. This data were recorded at physiological maturity. Leaf area was calculated as Leaf length × Leaf width × 0.71 following Krishnamurthy et al. (1974). The data were subjected to analysis of variance to see individual treatment and interaction effects for all the measured quantitative characters using SPSS Release 17 Software. Moreover, paired t-test was performed in all means of the measured characters to evaluate the trend of the differences between irrigated and non-irrigated treat-ments using the protected LSD procedure at P = 0.05 (Carmer and Swanson, 1973).
RESULTS
The result of this study showed a high significant difference in the genotypes for all the traits measured; indicating that the tested genotypes were diverse. Analysis of variance (ANOVA) showed highly significant difference (p<0.01) for genotypes factor for all the traits measured, which indicated that the tested genotypes were diverse. Highly significant differences were also observed among the genotypes for leaf area, head length, head weight, grain yield, and hundred seed weight in response to the induced post-anthesis drought stress, which was further confirmed by paired t-test (Table 1). In these characters, the stressed plots showed reduced mean values. Although, significant differences were observed between mean grain yield, panicle exsertion and head weight of irrigated and non-irrigated sets of genotypes with higher values being in the former (Table 2), genotype-by-irrigation interaction was not significant for these traits except head weight (Tables 3 and 4).
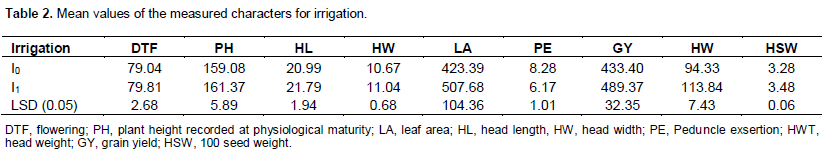
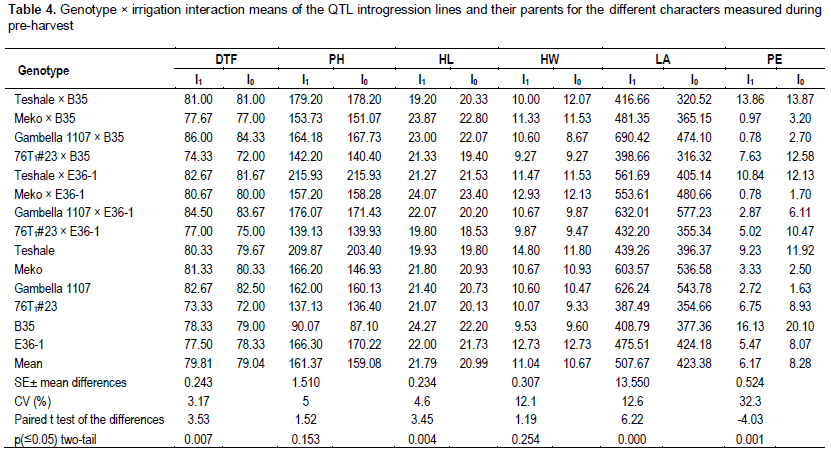
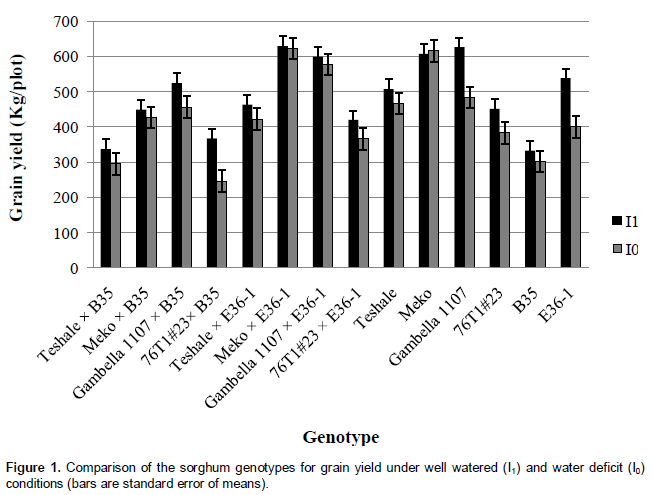
Grain yield had significant positive correlation (p<0.05) with head weight (r=0.66) and hundred seed weight (r=0.52) under moisture stressed condition. The reduction in leaf area was significant and ranged from 8.3% (B35) to 45.6% (Gambella 1107 × B35) in response to the changed irrigation levels. Genotype-by-irrigation interaction was significant for head weight indicating differential response of the genotypes in response to irrigation levels for this trait. There was a wide range of head weight reduction between 0 and 78%. Head weight had positive correlation (r=0.66) with grain yield under non-irrigated condition and even the correlation was stronger (r= 0.87) under irrigated condition. Even though there is significant difference among the mean grain yield and panicle exsertion of irrigated and non-irrigated sets of genotypes, the absence of genotype-by-irrigation interaction in these traits may indicate that moisture stress at the post-anthesis stage affects these traits regardless of genotype. In a well-watered condition, Gambella 1107 gave the highest and B35 the lowest yield of all the parental lines. Meko × E36-1 gave the highest yield of all the introgression lines under both water regimes. Meko and its introgression line, Meko × E36-1 gave comparative grain yield under both well watered and water deficit conditions. On the contrary, B35 and its introgression lines with 76T1#23 and Teshale gave relatively lower grain yield (Figure 1). Irrigation factor was significant (p<0.05) for hundred seed weight. When the data were pooled for all the genotypes, the range of reduction of hundred seed weight in response to the induced post-flowering drought was 0.5% (76T1#23 × B35) to 34.0% (Teshale × E36-1). All of the introgression lines, except Teshale × E36-1 and crosses of Gambella 1107, had lower sensitivity to the simulated drought stress response to seed weight than their recurrent parents (Table 3). Hundred seed weight had strong positive correlation (r=0.52) with leaf area. It had also very high positive correlation (r=0.79) with head weight under moisture stress and even higher positive correlation (r=0.84) under irrigated condition. Consequently, withholding irrigation during post-anthesis stage significantly reduced weight of the seeds by affecting grain filling (Table 2).
Genotype-by-irrigation interaction was also significant for hundred seed weight indicating differential response of the genotypes to the varying irrigation levels for this trait. However, hundred seed weight in the introgression lines alone had no correlation with leaf area (r=0.427, p=0.164) under stressed condition. It had also very high positive correlation with head weight under both moisture stress (r=0.887, p=0.003) and well watered (r=0.876, p=0.004) conditions. Irrigation factor was also significant (p<0.05) for peduncle exsertion. Twelve of the 14 introgression lines showed an increase in peduncle exsertion in response to moisture stress. The highest increase was observed in 76T1#23 × E36-1 followed by 76T1#23 × B35, both introgression lines of 76T1#23 with the two stay green source lines.
Leaf senescence is the most pertinent trait so far as this study was concerned. Five of the 14 genotypes, viz., Meko and Gambella 1107 and their introgression lines Meko × E36-1, Gambella 1107 × E36-1, Gambella 1107 × B35 showed early leaf senescence in one or more of the stressed plots. However, high level of average leaf senescence was visually observed in Gambella 1107 × B35, Meko, and Gambella 1107. The stay-green source line, B35 and its introgression lines 76T1#23 × B35 and Meko × B35 (Figure 2) maintained their green leaves until maturity.

DISCUSSION
Leaf characters
Even though no leaf size increase is generally expected beyond flowering, the reduction in leaf area in this experiment was probably due to contraction of the full sized leaves in response to the induced moisture deficit. However, leaf area was highly and positively correlated with grain yield (r=0.885, p=0.004) under moisture deficit. It has been reported that decreasing leaf area reduces crop water use and results in reduced grain yield (Borrell et al., 2014; Vadez et al., 2014). Early studies by Swanson (1941) and Blum et al. (1989) showed that early maturing varieties having a small leaf area are the most efficient in the production of grain per unit of leaf area whereas later varieties have a higher biomass, higher daily transpiration and a longer transpiration period and are likely to suffering dry seasons. In the present study, most of the introgression lines showed higher sensitivity to moisture stress with respect to the change in leaf area. Perhaps that was one mechanism of drought tolerance beyond the maintenance of green leaf area that the stg QTLs are contributing. In their recent study aiming at investigating the positive effect of Stg QTLs on grain yield under drought, Borrell et al. (2014) concluded that leaf area and transpiration per unit leaf area could be some of the mechanisms that Stg loci have impacts to regulate transpiration on the demand for water. Although, the reduction in the mean values in the remaining characters is a disadvantage, the reduction in leaf area could be considered as an important adaptive physiological mechanism expressed in response to the changed osmotic potential contributing to drought tolerance. In agreement with this finding, Swanson (1941) reported that leaf area was the greatest when there was an abundant moisture supply and the influence of drought on leaf development was shown by the results when the rainfall was deficient throughout the growing season. The same study showed that leaf area per acre in droughty season was 73 to 81% less than when there was abundant moisture and very heavy leaf development.
Inheritance study of Walulu et al. (1994) suggested that the stay green traits in one of the known source lines, B35 is influenced by a major gene but, later, it has been mapped as a quantitative trait and four QTLs controlling the stay-green (Stg1 through Stg4) were identified (Xu et al., 2000). Moreover, the effect of environment on expression of the stay-green trait in sunflower was suggested by Cukadar-Olmedo and Miller (1997). Therefore, these show that multi-location testing of the introgression lines would have helped the evaluation complete. The stay-green (non-senescence) trait in sorghum is reported to be often associated with good plant health and increased plant resistance to insects and diseases (Cukadar-Olmedo and Miller, 1997). Moreover, as it results in greater functional leaf area during grain filling and reduces the need for translocation of stored assimilates from the stem during grain filling, non-senescent sorghum accumulates more soluble sugars in the stem than does senescent sorghum, both during and after grain filling (McBee et al., 1983), which in turn improves the digestible energy content of the Stover (van Oosterom et al., 1996). In addition, stay-green QTLs may improve Stover digestibility by 3 to 5% units without negatively affecting grain and Stover yields (Reddy et al., 2012). This is also a very important trait in places like in Ethiopia where sorghum Stover is equally valued for cattle feed.
Yield and yield attributes
Although grain yield is a function of head weight and hundred grain weight, the significance of genotype × irrigation interaction in these characters in this experiment was not adequate to bring about a parallel significance in this major trait of importance. Eck and Musick (1979) found that a 27-day stress period (with average afternoon leaf water potential, - 22.7 bars) beginning at early grain filling reduced yields by 12% only. Similarly, in the present experiment whereby the non-irrigated treatments were exposed to a month of drought stress, the range of yield reduction in all the genotypes was from 1.0% (Meko × E36-1) to 49.0% (76T1#23 × B35) with an average of 12.9%. The variety Gambella 1107 was the most sensitive local variety that showed a yield reduction of 29.4%. On the other hand, the variety Meko was the least sensitive, which performed similarly in both water regimes. Unexpectedly, the stay green source line, E36-1 showed 34.2% yield reduction under drought stress. However, B35 had very low yield reduction indicating its low sensitivity (stability) to the induced drought, which in turn confirms its tolerance to drought stress. The introgression line Gambella 1107 × E36-1 showed better grain yield than its recurrent parent, Gambella 1107 under moisture stress. Meko × E36-1 was the highest yielding of all the genotypes included in the experiment, but had no advantage over its recurrent parent, Meko (Figure 1). Earlier reports showed that yield increases in stay-green types have been directly associated with maintenance of photosynthetic capability during the grain filling period (McBee, 1984; Wolfe et al., 1988) and longevity of a leaf is intimately related to its nitrogen status or water availability (Thomas and Rogers, 1990; Borrell et al., 2001). In the present preliminary experiment, although 76T1#23 × B35 maintained >50% of its green leaves until maturity, its yield reduction by 49% was probably because it possessed a cosmetic type (types C and D) of stay-green (Borrell et al., 2014).
Reduction in yield components as a result of drought stress was previously reported. For instance, similar to the present study (Table 4), Hooker (1985) found a decrease in head weight and weight of the 100 seeds associated with reduction in soil moisture by affecting grain filling. In their comparative study of senescent and non-senescent sorghum genotypes, Duncan et al. (1981) also found that the non-senescent (stay-green) genotypes had higher test weight. However, genotypic differences do exist for the period they take to fill their grain due to their efficiency in using the available moisture during post anthesis deficit. For instance, in the present study the variety Meko and its introgression line with E36-1 showed more or less similar grain yield in both moisture regimes. Kassahun et al. (2010) reported the low yield potential of B35 due to its small panicle size, non-tillering and small number of seeds per panicle. The lower grain yield in B35 and its introgression lines than their respective elite recurrent parents was in agreement with the findings of Kassahun et al. (2010) and may be an indication of yield drag caused by one or more of the stay-green QTLs of B35. Gambella 1107 × B35 had the same stay-green QTL (StgA) as that of 76T1#23 × B35, but it senesced earlier, which is an indication of the possibility of existence of QTL × Genotype or QTL × Genotype × Environment interaction for phenotypic expression of the trait, which calls for multilocation testing.
Previous reports showed that increased peduncle exsertion in rice was found to be associated with drought resistance and is controlled genetically (O’Toole and Cruz, 1983). This was in agreement with the present study and can be considered as an indirect selection criterion for drought tolerance in sorghum.
CONCLUSION
The scarcity of water associated with the impending climate change demands introduction and development of climate smart crops. Sorghum is one of these crops due to its adaptation to survive in severely stressed environments. Among the cope up strategies that sorghum as a climate smart crop possesses are drought and heat tolerance. Stay-green is a post-anthesis drought tolerance mechanism. In this experiment, stay-green QTLs were introgressed from known sources to the farmer preferred Ethiopian local sorghum varieties. Eight of the resulting introgression lines those possessed at least one Stg QTL were organized in an experiment to observe their performance under well watered and water deficit conditions. The result of this single season and single location experiment has indicated that most of the introgression lines maintained their green leaf area until maturity under conditions of post anthesis moisture deficit. However, most of them did not show better grain yield than their recurrent parents probably because the introgressed stay-green was cosmetic type. The reduction in leaf area was significant in the Stg QTL introgression lines under moisture stress perhaps because the QTLs play significant role in leaf area dynamics. Multienviroment testing of the performance of the introgression lines may give better understanding of the effect of the introgressed QTLs on the various characters.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interest.
ACKNOWLEDGEMENT
The introgression part of the study was supported by the project from Syngenta Foundation for Sustainable Agriculture through ICRISAT. We thank the research and technical staff of the national sorghum research program at Melkassa Agricultural Research Center for handling the backcross populations.
REFERENCES
| Adugna A (2007). The role of introduced sorghum and millets in Ethiopian agriculture. J. SAT Agric. Res. 3:1. | ||||
|
Balota M, Payne WA, Rooney W Rosenow D (2008). Gas exchange and transpiration ratio in sorghum. Crop Sci. 48:2361- 2371. Crossref |
||||
| Bioversity International (2010). Key access and utilization descriptors for sorghum genetic resources. Rome, Italy. | ||||
|
Blum A, Mayer J, Golan G (1989). Agronomic and Physiological Assessments of Genotypic Variation for Drought Resistance in Sorghum. Aust. J. Agric. Res. 40:49-61. Crossref |
||||
|
Borrell A, Hammer G, van Oosterom E (2001). Stay-green. A consequence of the balance between supply and demand for nitrogen during grain filling? Ann. Appl. Biol. 138: 91-95. Crossref |
||||
|
Borrell AK, Hammer GL, Henzell RG (2000). Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci. 40:1037-1048. Crossref |
||||
|
Borrell AK, Mullet JE, George-Jaeggli B, van Oosterom EJ, Hammer GL, Klein PE, Jordan DR (2014). Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth, and water uptake. J. Exp. Bot. |
||||
|
Carmer SG, Swanson MR (1973). An evaluation of ten pairwise multiple comparison procedures by Monte Carlo methods. J. Amer. Stat. Assoc. 68:66-74. Crossref |
||||
| Cates JS (1922). Kafir, the crop camel. Count. Gent. 87: 5. | ||||
|
Cattivelli L, Rizza F, Badeck F-W, Mazzucotelli E, Mastrangelo AM, Francia E, Mare`C, Tondelli A, Stanca AM (2008). Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 105:1-14 Crossref |
||||
|
Cukadar-Olmedo B, Miller JF (1997). Inheritance of the Stay Green Trait in Sunflower. Crop Sci. 37: 150-153. Crossref |
||||
|
Duncan RR, Bockholt AJ, Miller FR (1981). Descriptive comparison of senescent and nonsenescent sorghum genotypes. Agron. J. 73:849-853. Crossref |
||||
|
Eck HV, Musick JT (1979). Plant water stress effects on irrigated grain sorghum I. effects on yield. Crop Sci. 19:589-592. Crossref |
||||
| Ejeta G, Knoll JE (2007). Marker-assisted selection in sorghum. In: Varshney RK, Tuberosa R (Eds) Genomics Assisted Crop Improvement-Vol.2. Genomics Applications in Crops. Springer, 187-205. | ||||
|
Hooker ML (1985). Grain Sorghum Yield and Yield Component Response to Timing and Number of Irrigations. Agron. J. 77:810-812. Crossref |
||||
| IBPGR/ ICRISAT (1993). Descriptors for sorghum [Sorghum bicolor (L.) Moench]. International Board of Plant Genetic Resources, Rome, Italy. ICRISAT, Patancheru, India. | ||||
|
Kassahun B, Bidinger FR, Hash CT, Kuruvinashetti MS (2010). Stay-green expression in early generation sorghum [Sorghum bicolor (L.) Moench] QTL introgression lines. Euphytica 172:351-362. Crossref |
||||
|
Krishnamurthy K, Jagannath MK, Rajashekara BG, Raghunatha G (1974). Estimation of Leaf Area in Grain Sorghum from Single Leaf Measurements. Agron. J. 544-545. Crossref |
||||
|
Mahalakshmi V, Bidinger FR, Raju DS (1987). Effect of timing of water stress on pearl millet (Pennisetum americanum). Field Crops Res.15:327-339. Crossref |
||||
| McBee GG (1984). Relation of senescence, nonsenescence, and kernel maturity to carbohydrate metabolism in sorghum. In: Mughogho LK (Ed.) Sorghum root and stalk diseases, a critical review. Proceedings of the consultative group discussion of research needs and strategies for control of sorghum root and stalk diseases. Bellagio, pp. 119-129. | ||||
|
McBee GG, Waskom RM, Miller FR, Creelman RA (1983). Effect of senescence and nonsenescence on carbohydrates in sorghum during late kernel maturity states. Crop Sci. 23: 372-376. Crossref |
||||
|
O'Toole JC, Cruz RT (1983). Genotypic variation in epicuticular wax of rice. Crop Sci. 23: 392-394. Crossref |
||||
| Quinby JR (1974). Sorghum improvement and the genetic of growth. Texas A&M University Press, 108. | ||||
| Reddy BVS, Kumar AA, Sharma HC, Rao SP, Blümmel M, Reddy RC, Sharma R, Deshpande S, DattaMazumdar S, Dinakaran E (2012). Sorghum improvement (1980-2010): Status and way forward. J. SAT Agric. Res. 10. | ||||
|
Reddy BVS, Ramesh S, Reddy PS, Kumar AA (2009). Genetic enhancement for drought tolerance in sorghum. In: Janick J (Ed.) Plant Breeding Reviews. John Willey & Sons, Inc. pp. 189-222. Crossref |
||||
| Rosenow DT, Clark LE (1981). Drought tolerance in sorghum. In: Loden HD, Wilkinson D (Eds.) Proceedings of the 36th annual corn and sorghum industry research conference, 9-11 Dec. 1981, Chicago, (Eds.).American Seed Trade Association, Washington DC, pp. 18-21. | ||||
|
Rosenow DT, Quisenberry JE, Wendt CW (1983). Drought tolerant sorghum and cotton germplasm. Agric.Water Manag. 7:207-22. Crossref |
||||
| Serraj R, Hash TC, Buhariwalla HK, Bidinger FR, Folkertsma RT, Chandra S, Gaur PM, Kashiwagi J, Nigam SN, Rupakula A, Crouch JH (2005). Marker-assisted breeding for crop drought tolerance at ICRISAT: Achievements and prospects. In: Tuberosa R, Phillips RL, Gale M (Eds.), Proceedings of the international congress "In the wake of the double helix: From the green revolution to the gene revolution", 27-31 May 2003, Bologna, Italy, pp. 217-238. | ||||
|
Stout DG, Simpson GM (1978). Drought resistance of Sorghum bicolor. I. Drought avoidance mechanisms related to leaf and water status. Canad. J. Plant Sci. 58:213-224. Crossref |
||||
|
Swanson AF (1941). Relation of Leaf Area to Grain Yield in Sorghum. Agron. J. 908-914. Crossref |
||||
| Thomas H, Rogers LJ (1990). Turning over an old leaf. Univ. Wales Rev. Sci. Tech. 6:29-38. | ||||
|
Vadez V, Kholova J, Medina S, Kakkera A, Anderberg H (2014). Transpiration efficiency: new insights into an old story. J. Exp. Bot. Crossref |
||||
|
van Oosterom EJ, Jayachandranan R, Bidinger FR (1996). Diallel Analysis of the Stay-Green Trait and Its Components in Sorghum. Crop Sci. 36: 549-555. Crossref |
||||
|
Walulu RS, Rosenow DT, Wcstcr DB, Nguyen HT (1994). Inheritance of the stay green trait in Sorghum. Crop Sci. 34:970-972. Crossref |
||||
|
Wolfe DW, Henderson DW, Hsiao TC, Alvino A (1988). Interactive water and nitrogen effects on senescence of maize. II. Photosynthetic decline and longevity of individual leaves. Agron. J. 80:865-870. Crossref |
||||
|
Xu W, Subudhi PK, Crasta OR, Rosenow DT, Mullet JE, Nguyen HT (2000). Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome 43:461-469. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


