Full Length Research Paper

ABSTRACT
Increased production of sugar beet under rainfed conditions on saline–sodic soils in the Iranian areas highlights the importance of salt tolerant varieties. Screening of genotypes for salinity tolerance is difficult in field due to heterogeneity of physical and chemical properties of soil. In order to evaluate the salinity tolerance of 21 sugar beet monogerm O-types lines, a pot experiment was conducted using a split plot design. The evaluation of plants was performed using 11 morphological and physiological traits at vegetative growth stage under severe salt stress (∼16 dS m-1) and control (0.3 dS m-1) for 8 weeks. Salinity stress significantly reduced weight related traits. The response of genotypes for total weights and stem weights was very similar under both conditions. But, ranking of O-type lines for root weights under normal and stress condition was different. Indeed, there was high significant genotype × treat interaction for two these traits. Cluster analysis by using STI index of all traits allowed the identification of tolerant, moderate tolerant and sensitive genotypes toward salinity. The four salt-tolerant genotypes, O-type 9669, O-type 1609, O-type 463-2, and O-type 463-5 identified in this study, could be used in the development of salt-tolerant sugar beet varieties. In the second part of this study in order to assess a simple, rapid, and nondestructive method to estimate chlorophyll content, the chlorophyll meter (SPAD 502) readings were recorded and the relation was determined. Regression analysis indicated that there was a signiï¬cant linear regression between chlorophyll content and chlorophyll meter and about 74% of changes in chlorophyll meter based on chlorophyll content were predicted.
Key words: Sugar beet (beta vulgaris L.), salt tolerant index, screening, hybrid.
INTRODUCTION
MATERIALS AND METHODS

RESULTS AND DISCUSSION
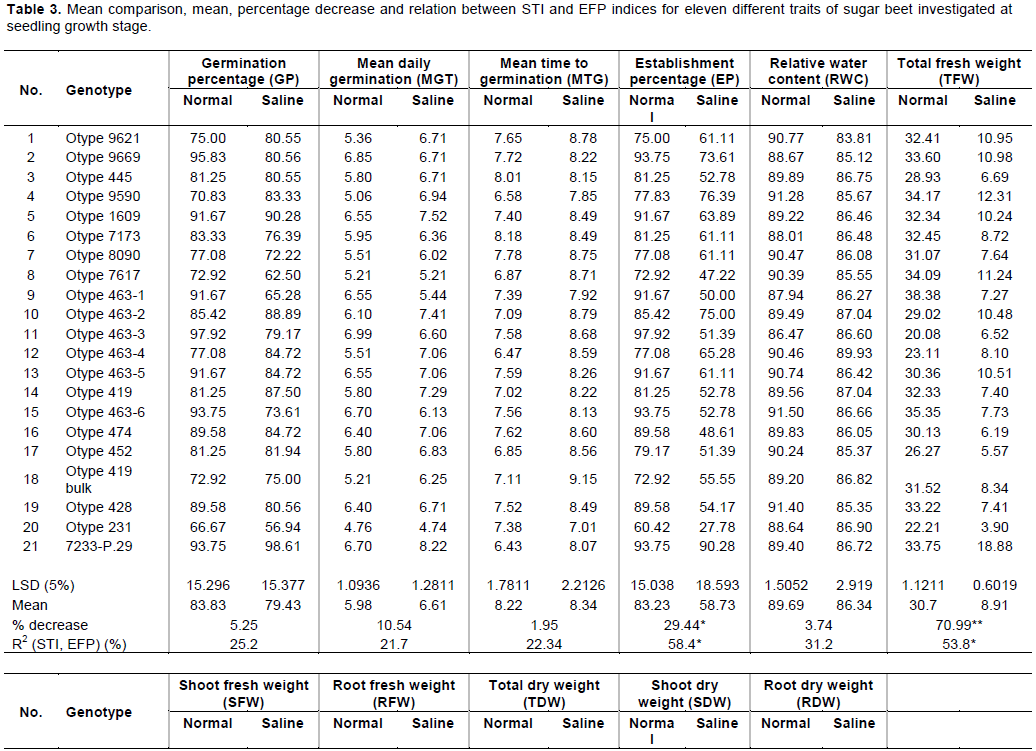
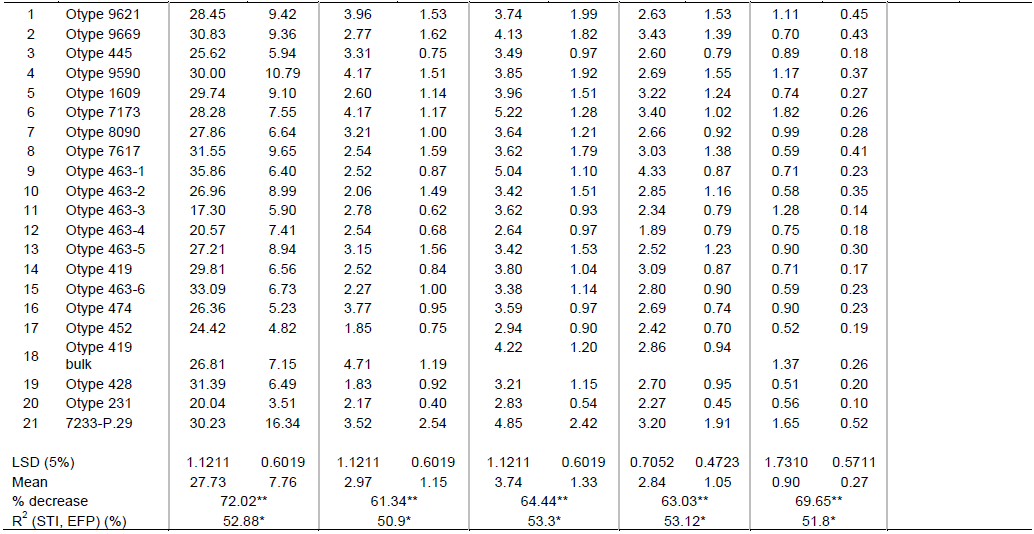
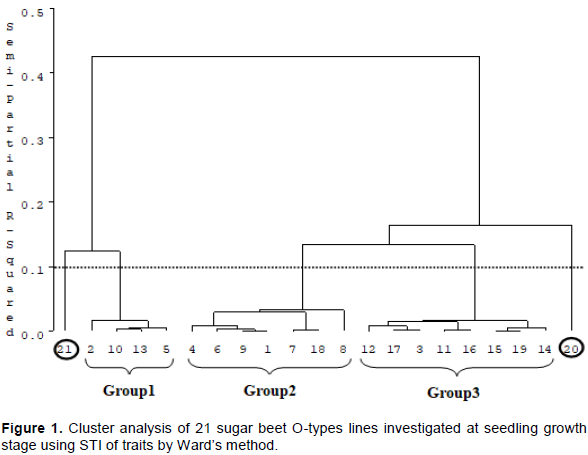
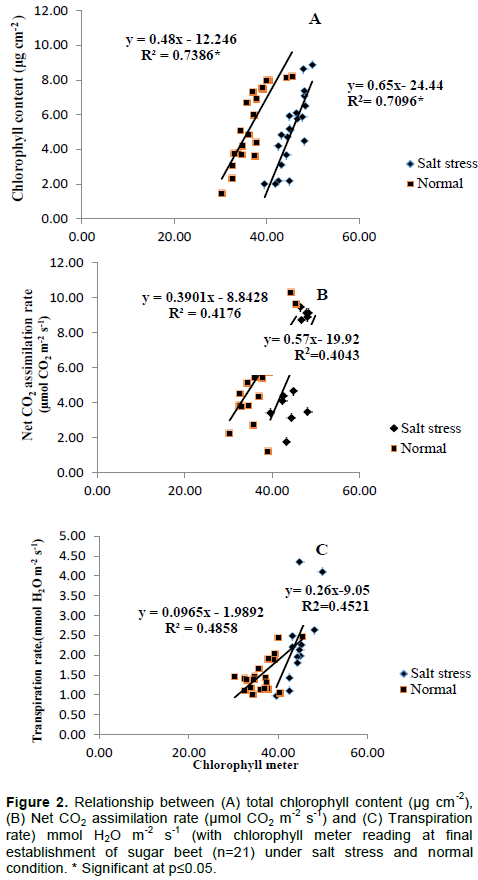
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abbasi Z, Golabadi M, Khayamim S, Pessarakli M (2018). The response of drought-tolerant sugar beet to salinity stress under field and controlled environmental conditions. Journal of Plant Nutrition 41(20):2660-2672. |
|
|
Abbasi Z, Arzani A, Majidi MM, Rajabi A, JalaliA (2019). Genetic analysis of sugar yield and physiological traits in sugar beet under salinity stress conditions. Euphytica 215(5):99-113. |
|
|
Balasubramanian V, Morales AC, Cruz RT, Thiyagarajan TM, Nagarajan R, Babu M, Abdulrachman S, Hai LH (2000). Adaptation of the chlorophyllmeter (SPAD) technology forreal-time N management in rice: a review. International Rice Research Notes 25:4-8. |
|
|
Bosemak NO (2006). Genetics and breeding.In Sugar beet, ed. A.P. Draycott, pp. 50-88. Oxford: Blackwell. |
|
|
Chapman SC, Barreto HJ (1997). Using a chlorophyll meter to estimate speciï¬c leaf nitrogen of tropical maize during vegetative growth. Agronomy Journal 89:557-562. |
|
|
Campbell RJ, Mobley KM, Marini RP, Pfeiffer DG (1990). Growing conditions alter the relationship between SPAD-510 values and apple leaf chlorophyll. Horticulture Science 25:330-331. |
|
|
Chikha MB, Hessini K, Ourteni RN, Ghorbel A, Zoghlami N (2016). Identification of barley landrace genotypes with contrasting salinity tolerance at vegetative growth stage. Plant Biotechnology 33(4):287-295. |
|
|
Dadkhah A, Rassam G (2017). Effect of short-term salinity on photosynthesis and ion relations in two sugar beet cultivars. Plant Physiology 7(2):1983-1989. |
|
|
De los Reyes BG, McGrathJM (2003). Cultivar-specific seedling vigor and expression of a putative oxalate oxidase germin-like protein in sugar beet (Beta vulgaris L.). Theoretical and Applied Genetics 107: 54-61. |
|
|
Durrant MJ, Gummerson RJ (1990). Factors associated with germination of sugarbeet seed in the standard test and establishment in the field. Seed Science and Technology 18(1):1-10. |
|
|
Eschie HA, Al-BarhiB, Al-GheityS, Al-KhanjariS (2002). Root and shoot growth in salinity-stressed Alfaalfa in response to nitrogen source. Journal of Plant Nutrition 25:2559-2569. |
|
|
Esfahani M, Abbasi HA, Rabiei B, Kavousi M (2008). Improvement of nitrogen management in rice paddy fields using chlorophyll meter (SPAD). Paddy and Water Environment 6(2):181-188. |
|
|
Fernandez GCJ (1991). Effective selection criteria for assessing plant stress tolerance.In: O.C.G.Kuo.(ed.). Adaptation of food crops to tempetature and water stress.Prov.Ann.Intn.Symp.Taiwan.13-18 Aug.Asian.Veget.Res.And.Develop.Center. |
|
|
Ghoulam C, FoursyA, FaresK (2002). Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environmental and Experimental Botany 47:39-50. |
|
|
Gidner S, Lennefors B-L, Nilsson N-O, Bensenfelt J, Johansson E, Gyllenspetz U, Kraft T (2005). QTL mapping of BNYVV resistance from the WB41 source in sugar beet. Genome 48:279-285. |
|
|
Hassani M, Heidari B, Mahmoudi SB, Taleghani DF, Stevanato P (2019). Identification of Owen-Type Male Sterility Maintainers Carrying Resistance AgainstRhizoctonia Crown and Root Rot (Rcrr) Disease in Sugar Beet Germplasm. Sugar Technology pp. 1-7. |
|
|
Hoagland DR, Arnon DI (1959). The water culture method for growing plants without soil. California Agricultural Experiment Station 307:32p. |
|
|
Jamil M, Rehman S, Rha ES (2007). Salinity effect on plant growth, PSII photochemistry and chlorophyll content in sugar beet (Beta vulgaris L.) and cabbage (Brassica oleraceacapitata L.).Pakistan Journal of Botany 39(3):753-760. |
|
|
Jamil A, Riaz S, Ashraf M, Foolad MR (2011). Gene expression profiling of plants under salt stress. Critical Reviews in Plant Sciences 30(5):435-458. |
|
|
Jangpromma N, Songsri P, Thammasirirak S, Jaisil P (2010). Rapid assessment of chlorophyll content in sugarcane using a SPAD chlorophyll meter across different water stress conditions. Asian Journal of Plant Sciences 9(6):368. |
|
|
Khayamim S, Tavkol Afshari R, Sadeghian SY, Poustini K, Roozbeh F, Abbasi Z (2014). Seed germination, plant establishment, and yield of sugar beet genotypes under salinity stress. Journal of Agriculture of Science 16:779-790. |
|
|
Kim J, Liu Y, Zhang X, Zhao B, Childs KL (2016). Analysis of salt-induced physiological and proline changes in 46 switchgrass (Panicumvirgatum) lines indicates multiple response modes. Plant Physiology and Biochemistry105:203-212. |
|
|
Lein JC, Sagstetter CM, Schulte D, Thurau T, Varrelmann M, Saal B, Koch G, Borchardt DC, Jung C (2008). Mapping of rhizoctonia root rot resistance genes in sugar beet using pathogen response-related sequences as markers. Plant Breeding 127:602-611. |
|
|
Li W, WangR, WangW, LiuH, LiuJ, ZhangS, Ai Y(2007). Effect of NaCl stress on sugar beet growth. Sugar Crops China 2:17-19. |
|
|
McGrath JM, Derrico CA, Morales M, Copeland LO, Christenson DR (2000). Germination of sugar beet (Beta vulgaris L.) seed submerged in hydrogen peroxide and water as a means to discriminate cultivar and seedlot vigor. Seed Science Technology 28(3):607-620. |
|
|
McGrath JM, ElawadyA, El-Khishin D, NaegeleRP, CarrKM, De los Reyes B( 2008). Sugar beet germination:Phenotypic selection and molecular profiling to identify genes involved in abiotic stress response. Acta Horticulture 782(35). |
|
|
Mini ML, Sathya M, Arulvadivookarasi K, Jayachandran KS ,Anusuyadevi M (2015). Selection of salt tolerant cowpea genotypes based on salt tolerant indices of morpho-biochemical traits. Current Trends in Biotechnology and Pharmacy 9(4):306-316. |
|
|
Minolta (1989). Chlorophyll Meter SPAD-502.Instruction Manual. Minolta Co., Ltd., Radiometric Instruments Operations, Osaka, Japan. |
|
|
Moritani M, Taguchi K, Kitazaki K, Matsuhira H, Katsuyama T, Mikami T, Kubo T (2013). Identiï¬cation of the predominant nonrestoring allele for Owen-type cytoplasmic male sterility in sugar beet (Beta vulgaris L.): development of molecular markers for the maintainer genotype. Molecular Breeding 32(1):91-100. |
|
|
Munns R, James RA (2003). Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 53:201-218. |
|
|
Ober ES, Rajabi A (2010).Abiotic stress in sugar beet. Sugar Tech 12(3-4):294-298. |
|
|
Owen FV (1945). Cytoplasmically inherited male-sterility in sugar beets. Journal of Agricultural Research 71:423-440. |
|
|
Peng S, Laza RC, Garcia FC, Cassman KG (1995b). Chlorophyll meter estimates leaf area-based N concentration of rice. Commun Soil Science Plant Analysis 26:927-935. |
|
|
Peng S, Garcia FC, Laza RC, Cassman KG (1993). Adjustment for speciï¬c leaf weight improves chlorophyll meter's estimation of rice leaf nitrogen concentration. Agronomy Journal 85:987-990. |
|
|
Peng C, Geng G, Yu L, Yang Y, Pi Z, Sun F, Sun X, Zhao H (2014). Effect of different Na+ concentrations on growth and physiological traits of sugar beet. Journal of Plant Nutition and Fertilization 20:459-465. |
|
|
Rozema J, Flowers TJ (2008). Crops for a salinized world. Science 322:1478-1480. |
|
|
Sakina A, AhmedI, ShahzadA, IqbalM, Asif M (2016). Genetic variation for salinity tolerance in Pakistani rice (Oryza sativa L.) germplasm. Journal of Agronomy and Crop Science 202(1):25-36. |
|
|
Smeal D, Zhang H (1994). Chlorophyll meter evaluation for nitrogen management in corn. Commun Soil Plant Analysis 25:1495-1503. |
|
|
Steel RGD, Torrie JH (1984). Principles and procedures of statistics: A biometrical approach. New York: McGraw Hill. |
|
|
Tahjib-UI-Arif M, Sohag AAM, Afrin S, Bashar KK, Afrin T, Mahamud AGM, Brestic M (2019). Differential response of sugar beet to long-term mild to severe salinity in a soil-pot culture. Agriculture 9(10):223-242. |
|
|
Turner FT, Jund MF (1991). Chlorophyll meter to predict nitrogen topdress requirement for semidwarf rice. Agronomy Journal 83:926-928. |
|
|
Uddling J, Gelang-AlfredssonJ, Piikki K, PleijelH(2007). Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynthesis Research 91:37-46. |
|
|
Wang Y, Stevanato P, Yu L, Zhao H, Sun X, Sun F, Li J, Geng G (2017). The physiological and metabolic changes in sugar beet seedlings under different levels of salt stress. Journal of Plant Research 130:1079-1093. |
|
|
Win KT, Aung ZO, Hirasawa T, Ookawa T, Yutaka H (2011). Genetic analysis of Myanmar Vigna species in responses to salt stress at the seedling stage. African Journal of Biotechnology 10 (9):1615-1624. |
|
|
Zhang L, Ma H, Chen T, Pen J, Yu S, Zhao X (2014). Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS ONE 9:e112807. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


