Full Length Research Paper

ABSTRACT
Sweet potato [Ipomoea batatas) (L.) Lam] is grown by small holder farmers across a wide range of environments in Malawi. A multi-location trial of eight genotypes for three seasons at six research stations was undertaken using additive main effects and multiplicative interaction (AMMI) model analysis to determine the genotypes’ stability and influence of genotype × environment interactions (GEI) on storage root yield. ANOVA showed high significant differences in storage root yield of the genotypes among seasons and locations (p≤0.01). Genotype, environment and genotype × environmental interaction significantly influenced storage root yield variation of the cultivars (p≤0.01). The variance in yield was mainly attributable to environment variability (62.86%) than genotypes variation (14.25%) and G × E interactions (15.06%). Semusa was superior for storage root yield (27.77t/ha) and Lu96/334 was the most inferior (11.19 t/ha). AMMI stability analysis revealed that LU96/303 (24.72 t/ha) was the most stable genotype across sites. Biplot analysis showed that Chitedze and Baka were sites conducive for high yields hence can be used for preliminary yield evaluation to capture maximum genotypes’ yield potential, while Lunyangwa was the lowest yields site; therefore useful for assessing the potential of worst performance of genotypes under unfavourable environmental conditions.
Key words: G × E interactions, multi-locational trial, stability, sweet potato, genotypes, root yield.
INTRODUCTION
MATERIALS AND METHODS

RESULTS AND DISCUSSION
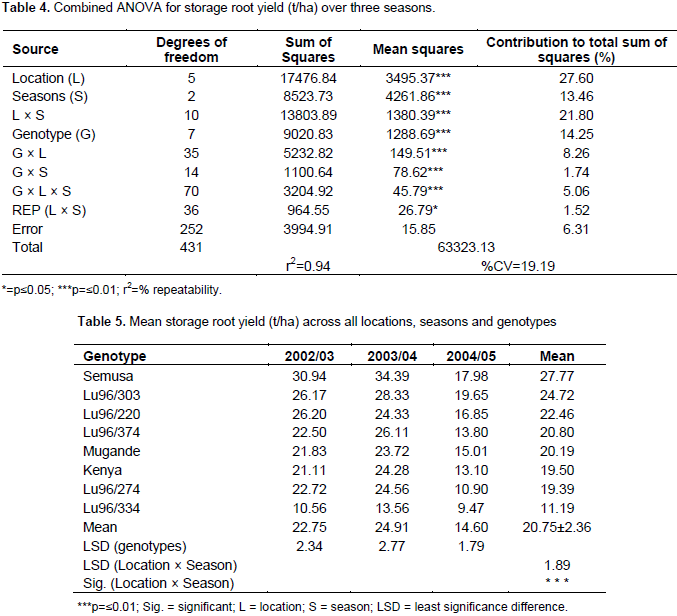
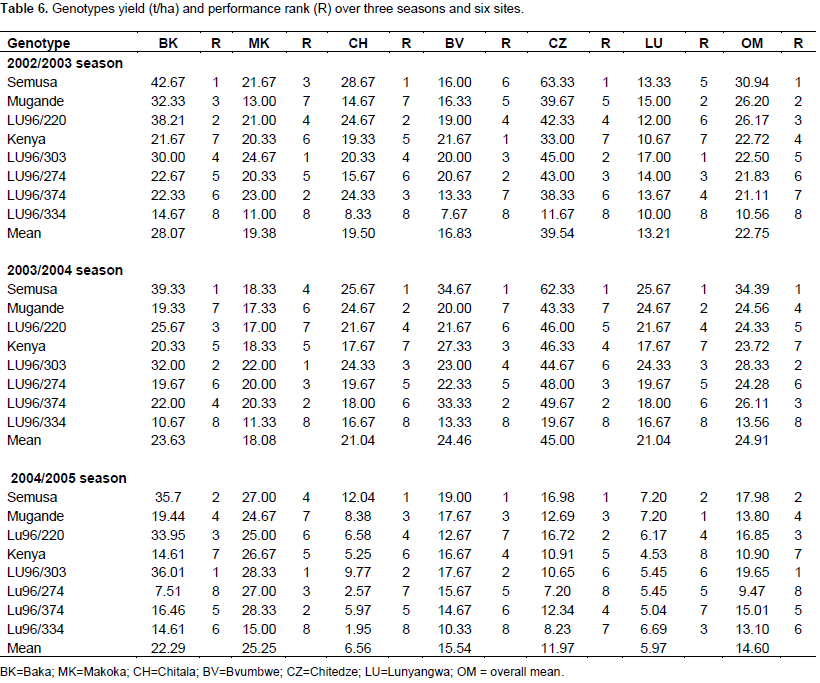
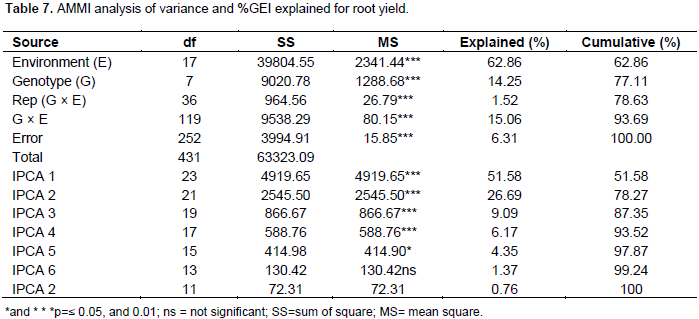
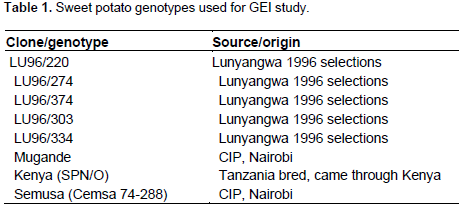
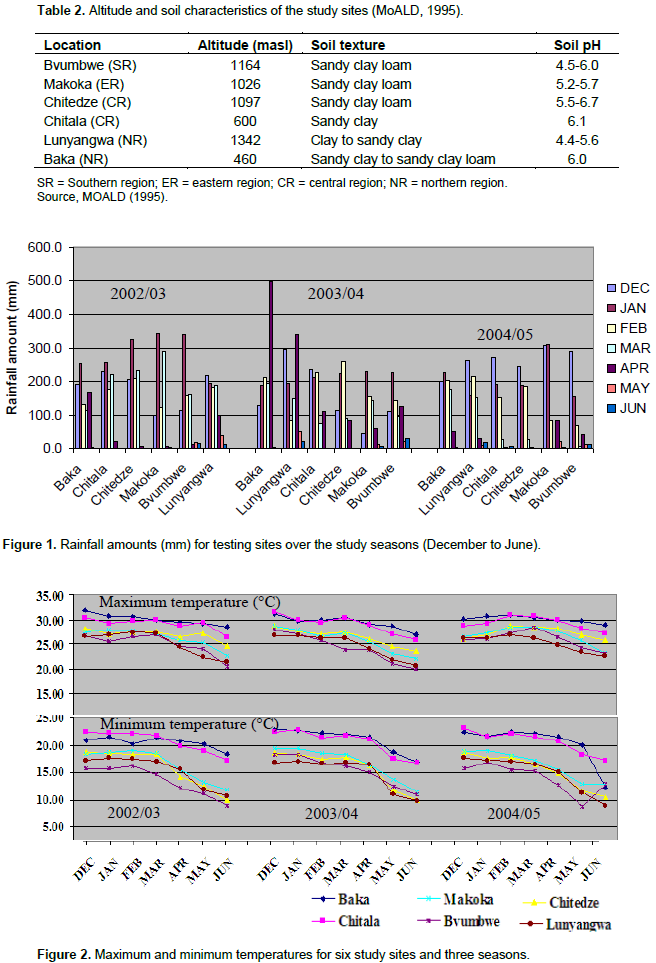
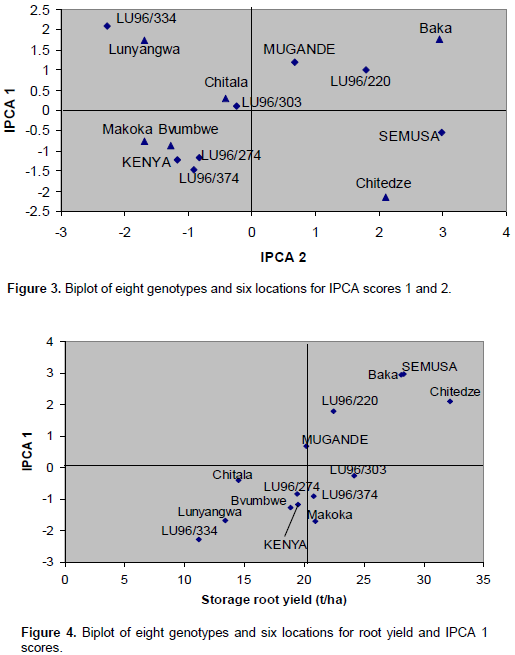
The magnitude of the GEI (15.06%) sum of squares was above that of genotypes (14.25%), indicating that there were differences in genotypic response across environments (Table 7). Results of the AMMI analysis (Table 7) also showed that the first principal component axis (PCA 1) captured 51.58% of the interaction sum of squares. Similarly, the second principal component axis (PCA 2) explained a further 26.69% of the GEI sum of squares. The mean squares for the PCA 1 and PCA 2 were significant at P≤0.01 and cumulatively contributed to 78.27% of the total GEI. A F-test at P≤0.01 suggested that the two principal component axes of interaction were significant for the model with 44 degrees of freedom. Zobel et al. (1988) and Kaya et al. (2002) showed in their prediction assessment that AMMI with only two IPCA axes was the best model. Further interaction principal component axes capture mostly noise and therefore did not help to predict the interactions (Daba et al., 2015). Thus, the interaction of the eight genotypes with 18 environments in this study was best predicted by the first two principal components and the rest were less informative. Thiyagu et al. (2013) reported in their stability study that sweet potato accumulated contribution of IPCA1 and IPCA2 of 85.26%. Comparably, in studies on linseed yield by Adugna and Labuschagne (2002), the two IPCAs accounted for 69.5% of the total interaction and the remaining 30.5% was considered residual or noise not interpretable and was therefore discarded. In order to identify genotypes adapted to specific test environments and their yield performance and stability, biplots were used (Yan, 2002). The biplot (Figure 3) was generated using genotypic and environmental scores of the first two PCA components as suggested by Vargas and Crossa (2000). Genotypes and location environments that appear almost on horizontal line (y axis) have a similar negative or positive interaction pattern for the IPCA 2 and equally those that fall along the x axis or close to it have similar interaction pattern along IPCA 1. Therefore, genotype LU96/334 is best for Lunyangwa while Kenya, LU96/274 and LU96/374 are best for Bvumbwe and Makoka and LU96/303 for Chitala. LU96/303 is located near the plot origins of both IPCA1 and 2 and therefore less responsive than the vertex genotypes, hence more stable across the testing environments since its IPCA scores are closer to zero (Figure 3) (Mwololo et al., 2009; Osiru et al., 2009; Thiyagu et al., 2013). Such a stable performance is a desirable attribute of cultivars, particularly in Malawi where environmental variations especially the rainfall pattern are very high and unpredictable as shown in Figure 2. Semusa was unstable (Figure 4) across the six sites when variation was explained by IPCA 1 but stabilised (Figure 3) when IPCA 2 was included. In contrast, LU96/334 is a non-adaptable genotype as indicated by its large IPCA 1 and IPCA 2 scores. Mugande and LU96/220 (high yielding) are moderately stable and are not closely associated with any site (Figure 3). Other genotypes of interest are LU96/274 and LU96/374 which are stable when explained by IPCA 2. Makoka and Bvumbwe are similar environments and Kenya, LU96/274 and LU96/374 exhibit similarity in interaction patterns (Figure 3).
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Adugna W, Labuschagne MT (2002). Genotype-environment and phenotypic stability analysis of linseed in Ethiopia. Plant Breed. 121:66-71. |
|
|
Agrobase TM (1999). Agronomic Software. 71 Waterloo St Winnipeg, R3N0S4, Canada. |
|
|
Allard RW (1960). Principles of plant breeding. John Wiley and Sons, NY, pp. 88-92. |
|
|
Altay F (2012). Yield stability of some Turkish winter wheat (Triticum aestivum L.) genotypes in the western transitional zone of Turkey. Turk. J. Field Crops 17:129-134. |
|
|
Andrade MI, Naico A, Ricardo J, Eyzaguire R, Makunde, GS, Ortiz R, Gruneberg W (2016). Genotype X environment interaction and selection for drought adaptation in Sweet potato (Ipomoea batatas [L] Lam) in Mozambique. Euphytica 209:261-280. |
|
|
Becker HC, Léon J (1988). Stability analysis in plant breeding. Plant Breed. 101:1-23. |
|
|
Bilbro JD, Ray LL (1976). Environmental stability and adaptation of several cotton cultivars. Crop Sci. 16:821-824. |
|
|
Boney W, Yalu A, Ramakrishna A, Deros M (2014). Genotypic Variability Estimates of Agronomic Traits for Selection in A Sweet potato [Ipomoea batatas (L.) Lam.] Poly crosses Population in Papua New Guinea. J. Plant Breed. Genet. 2(3):131-136. |
|
|
Chalwe A, Chiona M, Sichilima S, Njovu J, Chama C, Ndhlovu D (2017). Genotype stability index for root yield and tolerance to Sweet potato weevil (Cylas puncticolis): A tool for identifying climate smart varieties. Open Agric. 2:166-174. |
|
|
Chipungu FP, Benesi IRM, Moyo CC, Mkumbira J, Sauti RFN, Soko MM, Sandifolo V (1999). Field evaluation of introduced Sweet potato clones in Malawi. In: food security and crop diversification in SADC Countries: the role of cassava and sweet potato. Proceedings of the scientific workshop of the Southern African Root Crops Research Network (SARRNET). Pamodzi Hotel, Lusaka, Zambia. Akorod MO, Teri JM (Eds.). pp. 151-156. |
|
|
Comstock RE, Moll RH (1963). Genotype-environment interactions. In: Statistical genetics and plant breeding. National Academy of Sciences-National Research Council Publication 982. Hanson WD, Robinson HF (Ed.). NAS-NRC, Washington, DC. pp. 164-196. |
|
|
Daba C, Ayana A, Zeleke H, Wkjira A (2015). Genotype x Environment interactions for seed yield in Sesame in western Ethiopia. East Afr. J. Sci. 9(2):85-96. |
|
|
Dia M, Wehner TC, Hassell R, Price DS, Boyhan GE, Olson S, King S, Davis AR, Tolla GE (2016). Genotype x environment interaction and stability analysis for watermelon fruit yield in the United States. Crop Sci. 56:1645-1661. |
|
|
Eberhart SA, Russel WA (1966). Stability parameters for comparing varieties. Crop Sci. 6:36-40. |
|
|
Farshadfar E (2008). Incorporation of AMMI stability value and grain yield in a single non-parametric index (GSI) in bread wheat. Pak. J. Biol. Sci. 11:1791-1796. |
|
|
Fikere M, Fikiru E, Tadesse T, Legesse T (2009). Parametric stability analysis in field peas (Pisum sativum L.) under South Eastern Ethiopian condition. World J. Agric. Sci. 5:146-151. |
|
|
Gedif M, Yigzaw D (2014). Genotype by Environment Interaction Analysis for Tuber Yield of Potato (Solanum tuberosum L.) using a GGE Biplot Method in Amhara Region, Ethiopia. Agric. Sci. 5:239-249. |
|
|
Gruneberg WJ, Manrique K, Zhang D, Hermann M (2005). Genotype-by-environment interactions for a diverse set of Sweet potato clones evaluated across varying eco-geographic conditions in Peru. Crop Sci. 45:2160-2171. |
|
|
Kang MS, Magari R (1996). New developments in selecting for phenotypic stability in crop breeding. In: Genotype by environment interaction. Kang MS, Zobel Jr HG (Eds.). CRC Press, Boca Raton, New York. pp. 51-174. |
|
|
Kang MS (2002). Genotype-environment interaction: progress and prospects. In: Quantitative genetics and plant breeding. Kang MS (Ed.). CABI Pulishing, Wallingford, New York. pp. 221-243. |
|
|
Kaya YK, Palta E, Taner S (2002). Additive Main Effects and Multiplicative Interactions Analysis of Yield Performances in Bread Wheat Genotypes across Environments. Turk. J. Agric. 26:275-279. |
|
|
Khamphas S, Lertrat K, Lomthaisong K, Simla S, Suriharn B (2015). Effect of location, genotype and their interactions for anthocyanins and antioxidant activities of purple waxy corn cobs. Turk. J. Field Crops 20(1):15-23. |
|
|
Kivuva BM, Githiri SM, Yencho GC, Sibiya J (2014). Genotype and environment interaction for storage root yield in Sweet potato under managed drought stress conditions. J. Agric. Sci. 6:41-56. |
|
|
Makunde GS, Andrade MI, Ricardo J, Alvaro A, Menomussanga J, Gruneberg W (2017). Adaptation to mid- season drought in a Sweet potato (Ipomoea batatas [L] Lam) germplasm collection grown in Mozambique. Open Agric. 2:133-138. |
|
|
Ministry of Agriculture and Livestock Development (MoALD) (1995). The agricultural and livestock development strategy and action plan. Lilongwe, Malawi. |
|
|
Mitrović B, Stanisavljevi D, Treski S, Stojaković M, Ivanović M, Bekavac G, Rajković M (2012). Evaluation of Experimental Maize Hybrids Tested in Multi-Location Trials Using AMMI and GGE Biplot Analyses. Turk. J. Field Crops 17:35-40. |
|
|
Mulema JK, Adipala E, Olanya OM, Wagoire W (2008). Yield stability analysis of late blight resistant potato selections. Exp. Agric. 44:145-155. |
|
|
Mwololo JK, Muturi PW, Mburu MWK, Njeru RW, Kiarie N, Munyua JK, Ateka EM, Muinga RW, Kapinga RE, Lemaga B (2009). Additive main effects and multiplicative interaction analysis of genotype x environmental interaction among Sweet potato genotypes. J. Anim. Plant Sci. 2(3):148-155. |
|
|
Ntawuruhunga P, Rubaihayo P, Whyte JBA, Dixon AGO, Osiru DSO (2001). Additive main effects and multiplicative interaction analysis for storage root yield of cassava genotypes evaluated in Uganda. Afr. Crop Sci. J. 9:591-958. |
|
|
Osiru MO, Olanya OM, Adipala E, Kapinga R, Lemaga B (2009). Yield stability analysis of Ipomoea batatas L. cultivars in diverse environments. Aust. J. Crop Sci. 3(4):213-220. |
|
|
Piepho HP (1996). Analysis of genotype by environment interaction and phenotypic stability. In: Genotype by environment interaction. Kang MS, Zobel Jr HG (Eds.). CRC Press, Boca Raton, New York, pp. 151-174. |
|
|
Rea R, De Sousa Vieira O (2002). Genotype by environment interactions in sugar cane yield trials in the Central-Western Region of Venezuela. Interciencia 27 (11):620-624. |
|
|
Rukundo P, Shimelis H, Laing M, Gahakwa D (2013). Storage root formation, dry matter synthesis, accumulation and genetics in Sweet potato. Aust. J. Crop Sci. 7:2054-2061. |
|
|
Shafii B, Mahler KA, Price WJ, Auld DL (1992).Genotype-environment interaction effects on winter rapeseed yield and oil content. Crop Sci. 2:922-927. |
|
|
Sial MA, Arain MA, Javed MA, Rajput MA (2001). Genotype-environment interaction for grain yield in bread wheat. Proc. Pak. Acad. Sci. 38:41-46. |
|
|
Singh D, Guaf J, Okpul T, Wiles G, Hunter D (2006). Taro(Colocasia esculenta) variety release recommendations for Papua New Guinea based on multi-location trials. New Zealand J. Crop Hort. Sci. 34:163-171. |
|
|
Tekalign T (2007). Genotype 9 environment interaction for root yield of elite Sweet potato (Ipomoea batatas (L.) Lam.) genotypes. S. Afr. J. Plants Soil 24:144-146. |
|
|
Thiyagu D, Rafii MY, Mahmud TMM, Latif MA, Malek MA, Sentoor G (2013). Genotype by environment assessment in Sweet potato as leafy vegetable using AMMI model. Pak. J. Bot. 45(3):843-852. |
|
|
Vargas MV, Crossa J (2000). The AMMI analysis and graphing the biplot. CYMMIT, Mexico. pp. 1-23. |
|
|
Yahaya SU, Saad AM, Mohammed SG, Afuape SO (2015). Evaluating the performance of improved Sweet potato (Ipomoea batatas L. Lam) advanced lines in Kano, Sudan savanna of Nigeria. Int. J. Agron. Agric. Res. 7(4):52-60. |
|
|
Yan W, Rajcan I (2002). Biplot evaluation of test sites and trait relations of soybean in Ontario. Crop Sci. 42:11-20. |
|
|
Yan W (2002). Singular-value partitioning in biplot analysis of multi-environment trial data. Agron. J. 94:990-996. |
|
|
Zhang Y, He Z, Zhang A, Van Ginkel M, Ye G (2006). Pattern analysis on grain yield of Chinese and CIMMYT spring wheat cultivars grown in China and CIMMYT. Euphytica 147:409-420. |
|
|
Zobel RW, Wright MJ, Gauch HG (1988). Statistical analysis of yield trial. Agron. J. 80:388-393 |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


