Full Length Research Paper

ABSTRACT
INTRODUCTION
ADVANCEMENT OF DNA MARKER TECHNOLOGIES
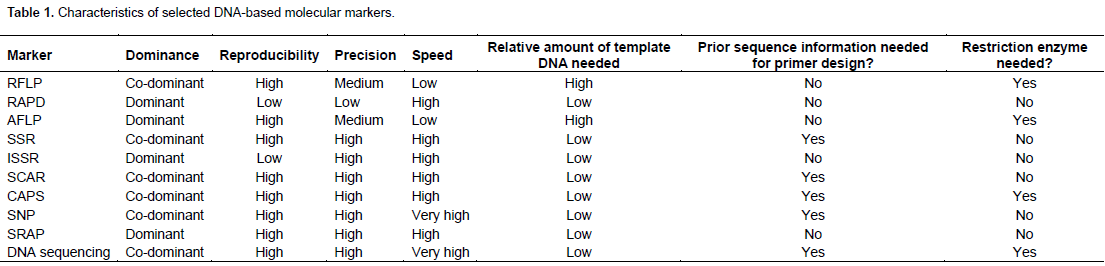
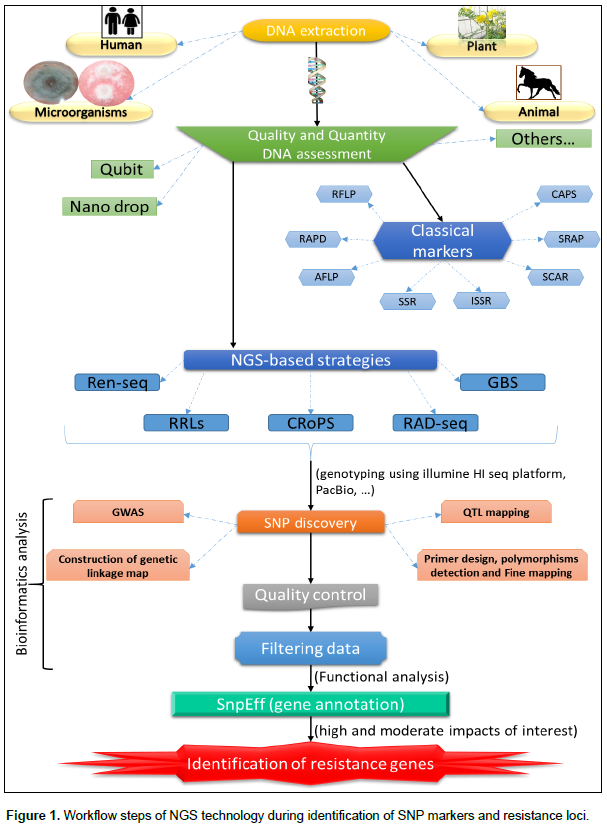
IDENTIFICATION OF KEY INTERACTIVE GENES BETWEEN PLANTS AND OOMYCETE PATHOGENS
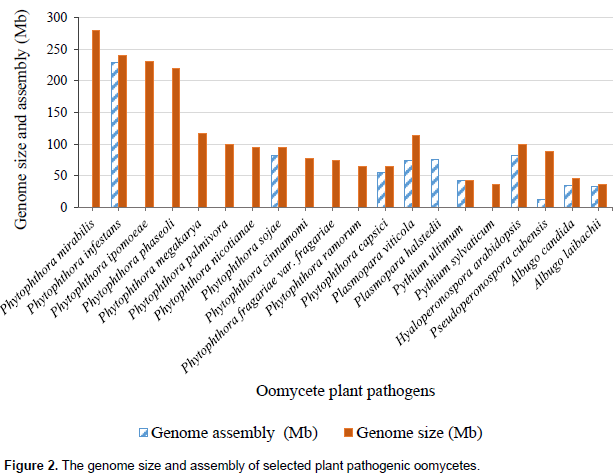

FUTURE DIRECTION OF BREEDING STRATEGIES FOR PLANT DISEASE RESISTANCE
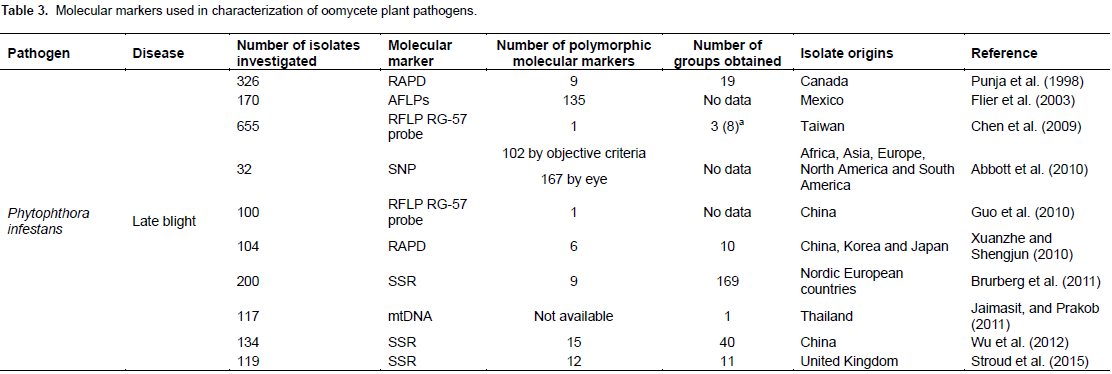
CONFLICT OF INTERESTS
REFERENCES
|
Abad ZG, Abad JA (2003). Advances in the integration of morphological and molecular characterization in the genus Phytophthora: The case of P. niederhauseria sp. nov. Phytopathology 93:S1. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Abbott C, Gilmore S, Lewis C, Chapados J, Peters R, Platt H, Coffey MD, Lévesque CA (2010). Development of a SNP genetic marker system based on variation in microsatellite flanking regions of Phytophthora infestans. Can. J. Plant Pathol. 32(4):440-457. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Altshuler D, Pollara VJ, Cowles CR, Van Etten WJ, Baldwin J, Linton L, Lander ES (2000). An SNP map of the human genome generated by reduced representation shotgun sequencing. Nature 407:513-516. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Arafa RA, Rakha MT, Soliman NEK, Moussa OM, Kamel SM, Shirasawa K (2017). Rapid identification of candidate genes for resistance to tomato late blight disease using next-generation sequencing technologies. Plos One 12:e0189951. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Arafa RA, Soliman NEK, Moussa OM, Kamel SM, Shirasawa K (2018). Characterization of Egyptian Phytophthora infestans population using simple sequence repeat markers. J. Gen. Plant Pathol. 84:104-107. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Aragaki M, Uchida JY (2001). Morphological distinctions between Phytophthora capsici and P. tropicalis sp. nov," Mycologia 93:137-145. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Baird NA, Etter PD, Atwood TS, Currey MC, Shiver AL, Lewis ZA, Selker EU, Cresko WA, Johnson EA (2008). Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS One 3:e3376. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Baxter SW, Davey JW, Johnston JS, Shelton AM, Heckel DG, Jiggins CD, Blaxter ML (2011). Linkage mapping and comparative genomics using next-generation RAD sequencing of a non-model organism. PLoS One 6:e19315. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Bellin D, Peressotti E, Merdinoglu D, Wiedemann-Merdinoglu S, Adam-Blondon AF, Cipriani G, Morgante M, Testolin R, Gaspero GD (2009). Resistance to Plasmopara viticola in grapevine 'Bianca' is controlled by a major dominant gene causing localized necrosis at the infection site. Theor. Appl. Genet. 120:163-176. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Brurberg MB, Elameen A, Le VH, Nærstad R, Hermansen A, Lehtinen A, Hannukkala A, Nielsen B, Hansen J, Andersson B, Yuen J (2011). Genetic analysis of Phytophthora infestans populations in the Nordic European countries reveals high genetic variability. Fungal Boil. 115(4): 335-342. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Burnham K, Dorrance A, VanToai T, St. Martin S (2003). Quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop Sci. 43:1609-1617. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Carlson MO, Gazave E, Gore MA and Smart CD (2017). Temporal genetic dynamics of an experimental, biparental field population of Phytophthora capsici. Front. Genet. 8:26. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chen AL, Liu CY, Chen CH, Wang JF, Liao YC, Chang CH, Tsai MH, Hwu KK, Chen KY (2014). Reassessment of QTLs for late blight resistance in the tomato accession L3708 using a restriction site associated DNA (RAD) linkage map and highly aggressive isolates of Phytophthora infestans. PLoS One 9(5):e96417. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chen CH, Wang TC, Black L, Sheu ZM, Perez F, Deahl K (2009). Phenotypic and genotypic changes in the Phytophthora infestans population in Taiwan-1991 to 2006. J. Phytopathol. 157(4):248-255. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Chunwongse J, Chunwongse C, Black L, Hanson P (2002). Molecular mapping of the Ph-3 gene for late blight resistance in tomato. J. Hortic. Sci. Biotechnol. 77(3):281-286. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Collard B, Jahufer M, Brouwer J, Pang E (2005). An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: the basic concepts. Euphytica 142:169-196. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Davey JW, Hohenlohe PA, Etter PD, Boone JQ, Catchen JM, Blaxter ML (2011). Genome-wide genetic marker discovery and genotyping using next-generation sequencing, Nat. Rev. Genet. 12:499-510. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
del Castillo-Múnera J, Cárdenas M, Pinzón A, Casta-eda A, Bernal AJ, Restrepo S (2013). Developing a taxonomic identification system of Phytophthora species based on microsatellites. Rev. Iberoam. Micol. 30(2):88-95. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Delmotte F, Machefer V, Giresse X, Richard-Cervera S, Latorse M, Beffa R (2011). Characterization of single-nucleotide-polymorphism markers for Plasmopara viticola, the causal agent of grapevine downy mildew. Appl. Environ. Microbiol. 77(21):7861-7863. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Devran Z, Kahveci E, O¨zkaynak E, Studholme DJ, To¨r M (2015). Development of molecular markers tightly linked to Pvr4 gene in pepper using next-generation sequencing. Mol. Breed. 35:101. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Drenth A, Whisson S, Maclean D, Irwin J, Obst N, Ryley M (1996). The evolution of races of Phytophthora sojae in Australia. Phytopathology 86(2):163-169. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Elshire RJ, Glaubitz JC, Sun Q, Poland JA, Kawamoto K, Buckler ES, Mitchell SE (2011). A robust simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS One 6:e19379. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Flier WG, Grünwald NJ, Kroon LP, Sturbaum AK, van den Bosch TB, Garay-Serrano E, Lozoya-Salda-a H, Fry WE, Turkensteen LJ (2003). The population structure of Phytophthora infestans from the Toluca Valley of central Mexico suggests genetic differentiation between populations from cultivated potato and wild Solanum spp. Phytopathology 93(4):382-390. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Forster H, Oudemans P, Coffey MD (1990). Mitochondrial and nuclear DNA diversity within six species of Phytophthora. Exp. Mycol. 14(1):18-31. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fu YB, Cheng B, Peterson GW (2014). Genetic diversity analysis of yellow mustard (Sinapis alba L.) germplasm based on genotyping by sequencing. Genet. Resour. Crop Evol. 61:579-594. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Fulcher M, Carlson M, Smart C (2014). Geneva blight genetics: Genotyping-by-sequencing a bi-parental population of Phytophthora capsici," in Proceedings of the 2014 Summer Scholars Program, Geneva, NY. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gao R, Cheng Y, Wang Y, Guo L, Zhang G (2015). Genome sequence of Phytophthora fragariae var. fragariae, a quarantine plant-pathogenic fungus. Genome Announc. 3(2):e00034-15. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
GelvonauskienÄ— D, Rugienius R, Šikšnianas T, StanienÄ— G, Sasnauskas A, Stanys V (2007). Screening of apple and strawberry plants carrying fungal disease resistance oligogenes using molecular markers. Zemdirbyste Agric. 94:139-145. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gessler C, Pertot I, Perazzolli M (2011). Plasmopara viticola: a review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 50:3-44. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gobbin D, Pertot I, Gessler C (2003). Identification of microsatellite markers for Plasmopara viticola and establishment of high throughput method for SSR analysis. Eur. J. Plant Pathol. 109:153-64. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gobbin D, Rumbou A, Linde CC, Gessler C (2006). Population genetic structure of Plasmopara viticola after 125 years of colonization in European vineyards. Mol. Plant Pathol. 7(6): 519-531. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gobena D, Roig J, Galmarini C, Hulvey J, Lamour K (2012). Genetic diversity of Phytophthora capsici isolates from pepper and pumpkin in Argentina. Mycologia 104(1):102-107. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gore MA, Chia JM, Elshire RJ, Sun Q, Ersoz ES, Hurwitz BL, Peiffer JA, McMullen MD, Grills GS, Ross-Ibarra J, Ware DH, Buckler ES (2009). A first-generation haplotype map of maize. Science 326:1115-1117. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Grunwald NJ, McDonald BM, Milgroom MG (2016). Population genomics of fungal and oomycete pathogens. Annu. Rev. Phytopathol. 54:323-346. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Guo L, Zhu XQ, Hu CH, Ristaino J B (2010). Genetic structure of Phytophthora infestans populations in China indicates multiple migration events. Phytopathology 100(10):997-1006. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Guo ZZ (2011). Screening of molecular markers for downy mildew resistance introgression line of Cucumis Hystrix-C. Sativus and analysis of programmed cell death. China: Nanjing Agricultural University, Master's thesis |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Gururani MA, Venkatesh J, Upadhyaya CP, Nookaraju A, Pandey SK, Park SW (2012). Plant disease resistance genes: current status and future directions. Physiol. Mol. Plant Pathol.78:51-65. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hansen ZR, Everts KL, Fry WE, Gevens AJ, GruÈnwald NJ, Gugino BK, Johnson DA, Johnson SB, Judelson HS, Knaus BJ, McGrath MT, Myers KL, Ristaino JB, Roberts PD, Secor GA, Smart CD (2016). Genetic variation within clonal lineages of Phytophthora infestans revealed through genotyping-by-sequencing, and Implications for late blight epidemiology. PLoS One 11:e0165690. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Haymes K, Henken B, Davis T, Van de Weg W (1997). Identification of RAPD markers linked to a Phytophthora fragariae resistance gene (Rpf1) in the cultivated strawberry. Theor. Appl. Genet. 94:1097-1101. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Haymes K, Van de Weg W, Arens P, Maas J, Vosman B, Den Nijs A (2000). Development of SCAR markers linked to a Phytophthora fragariae resistance gene and their assessment in European and North American strawberry genotypes. J. Am. Soc. Hortic. Sci. 125:330-339. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
He J, Zhao X, Laroche A, Lu ZX, Liu H, Li Z (2014). Genotyping-by- sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Front. Plant Sci. 5:484. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hickman C (1941). The red core root disease of the strawberry caused by Phytophthora fragariae n. sp. J. Pomol. Hortic. Sci. 18(2):89-118. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hogenhout SA, Van der Hoorn RA, Terauchi R, Kamoun S (2009). Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 22:115-122. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hulvey J, Gobena D, Finley L, Lamour K (2010). Co-occurrence and genotypic distribution of Phytophthora species recovered from watersheds and plant nurseries of eastern Tennessee. Mycologia 102(5):1127-1133. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Hyten DL, Cannon SB, Song Q, Weeks N, Fickus EW, Shoemaker RC (2010). High-throughput SNP discovery through deep resequencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genomics 11:38. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Islam SZ, Babadoost M, Lambert KN, Ndeme A, Fouly HM (2005). Characterization of Phytophthora capsici isolates from processing pumpkin in Illinois. Plant Dis. 89(2):191-197. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jaimasit P, Prakob W (2011). Characterization of Phytophthora infestans population in potato crops from Chiang mai and Tak provinces. J. Agric. Technol. 7(2): 431-439. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jarne P, Lagoda PJ (1996). Microsatellites, from molecules to populations and back. Trends Ecol. Evol. 11:424-9. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Judelson HS (2002). Sequence variation and genomic amplification of a family of Gypsy-like elements in the oomycete genus Phytophthora. Mol. Biol. Evol. 19:1313-1322. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Jupe F, Witek K, Verweij W, Sliwka J, Pritchard L, Etherington GJ, Maclean D, Cock PJ, Leggett RM, Bryan GJ, Cardle L, Hein I, Jones JD (2013). Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 76:530-44. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kasuga T, Salimath SS, Shi J, Gijzen M, Buzzell RI, Bhattacharyya MK (1997). High resolution genetic and physical mapping of molecular markers linked to the Phytophthora resistance gene Rps1-k in soybean. Mol. Plant Microbe Interact. 10:1035-1044. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kato M (2010). Recent research on Phytophthora root and stem rot of soybean in Japan. Plant Prot. 64:497-500. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Kole C, Ashrafi H, Lin G, Foolad M (2006). Identification and molecular mapping of a new R gene, Ph-4, conferring resistance to late blight in tomato," Solanaceae Conference, University of Wisconsin, Madison. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lamour K, Kamoun S (2009). Oomycete genetics and genomics: diversity, interactions and research tools. John Wiley & Sons. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lebeda A, Urban J (2007). Temporal changes in pathogenicity and fungicide resistance in Pseudoperonospora cubensis populations. Acta. Hortic. 731: 327-336. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lebeda A, Widrlechner MP, Urban J (2006). Individual and population aspects of interactions between cucurbits and Pseudoperonospora cubensis: pathotypes and races. Proceedings of Cucurbitaceae, Asheville, North Carolina, USA. pp. 453-467. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lee JH, Park MH, Lee S (2016). Identification of Pseudoperonospora cubensis using real-time PCR and high resolution melting (HRM) analysis. J. Gen. Plant Pathol. 82(2):110-115. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lee S, Mian MR, McHale LK, Wang H, Wijeratne AJ, Sneller CH, Dorrance AE (2013). Novel quantitative trait loci for partial resistance to Phytophthora sojae in soybean PI 398841. Theor. Appl. Genet. 126:1121-1132. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Li P, Cao S, Dai Y, Li X, Xu D, Guo M, Pan YM, Gao ZM (2012). Genetic diversity of Phytophthora capsici (Pythiaceae) isolates in Anhui Province of China based on ISSR-PCR markers. Genet. Mol. Res. 11:4285-4296. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
MacGregor T, Bhattacharyya M, Tyler B, Bhat R, Schmitthenner AF, Gijzen M (2002). Genetic and physical mapping of Avr1a in Phytophthora sojae. Genetics 160(3):949-959. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Malvick D, Grunden E (2004). Traits of soybean-infecting Phytophthora populations from Illinois agricultural fields. Plant Dis. 88:1139-1145. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Martin FN, Abad ZG, Balci Y, Ivors K (2012). Identification and detection of Phytophthora: reviewing our progress, identifying our needs. Plant Dis. 96(8):1080-1103. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Mathey MM (2013). Phenotyping diverse strawberry (Fragaria spp.) germplasm for aid in marker-assisted breeding, and marker-trait association for red stele (Phytophthora fragariae) resistance marker Rpf1. USA. Oregon State University, Master's thesis P 144. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Meng X, Shoemaker R, Yang X (1999). Analysis of pathogenicity and genetic variation among Phytophthora sojae isolates using RAPD. Mycol. Res. 103(02):173-178.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


