Full Length Research Paper

ABSTRACT
Wide spread and indiscriminate use of antibiotics is accompanied by the emergence of microorganism that are resistant to these agents. Therefore, new approaches are required to address the problem of antimicrobial resistance. Epithelial linings of living organisms are the source of various antimicrobial peptides. Keeping this in view, the present experiment was designed to characterize one of the important natural antimicrobial peptide gene, that is, cathelicidin gene from the buffalo (Bubalus bubalis) uterine epithelium and explored its potency for use as template for synthesis of novel antimicrobial agents. Total RNA was isolated from epithelial layer of buffalo uterus and reverse transcribed using designed primers. The amplified PCR product was purified and cloned. Positive clone was sequenced and result was analysed using laser gene software (DNA Star, USA). The cDNA of uterus cathelicidin had 516 bases with complete ORF from 6-452 bp. The predicted pre-propeptide comprised of 148 amino acids. Mature active peptide had 18 amino acids and had five each of arginine and tryptophan and four proline residues. From this study, it can be concluded that the buffalo (Bubalus bubalis) uterus expressed a potent antimicrobial peptide and amino acid sequence of mature peptide can be used as template for synthesis of novel antimicrobial agents.
Key words: Antimicrobial peptide, buffalo, cationic peptide and cathelicidin.
INTRODUCTION
The availability of complete genome sequences and development of information technology have provided a greater opportunity for peptide based drug designing. The field of structure based drug designing is a rapidly growing area and the exposition of genomic, proteomic and structural information has provided new targets and opportunities for drug lead discovery. In the meat industry, the use of antibiotics as growth enhancers is a common practice and extensive use of antibiotic in meat industry causes an alarming increase of antibiotic resistance microbes across the world (Gorbach, 2001). Antibiotic resistance has been posing increasingly serious concern to the public, health specialist and animal food producers. Therefore, there is a need of alternative group of drugs which are active in vivo and are able to act fast and has broad-spectrum activity, do not induce bacterial resistance and have limited or no side effects. Antimicrobial peptides are prevalent throughout the nature as part of the intrinsic defenses of most organisms. These peptides represent ancient host defense molecules and act as key elements in non-specific immunity (Ganz and Lehrer, 1989). Their wide spread distribution throughout the animal and plant kingdoms, suggest that antimicrobial peptides have served a fundamental role in the successful evolution of complex multicellular organisms (Bals, 2000). New strategies are required for synthesis of novel antimicrobial agents to deal with the threat of bacterial resistance (Ravi et al., 2011).
Antimicrobial peptides hold promise as broad-spectrum alternatives to conventional antibiotics (Gee et al., 2013). Mamalian defensin and cathelicidin are the two broad classes of natural antimicrobial peptides constitute a large family of endogenous peptide antibiotics with broad spectrum activity against various bacteria, fungi and viruses. Expression of human cathelicidin namely hCAP-18 and LL-37 is reported respectively in the reproductive tract (Malm et al., 2000) and skin epithelial cell (Markus et al., 2012).
Several β-defensin namely, human β-defensin-4 from testis (Garica et al., 2001), cryptidin from mouse sertoli cells (Grandjean et al., 1997); Bin1b from rat epididymis (Li et al., 2001) has been isolated. Antimicrobial peptide gene from buffalo tongue has been sequenced and characterized (Kalita and Kumar, 2009). Synthesis of different length of natural analogue of buffalo lingual antimicrobial peptide and functional study revealed its potency against both gram positive and negative bacteria (Kalita et al., 2009). However till date, antimicrobial peptide gene in the uterine tract has not been characterized. Keeping this in view, the present study was carried out to characterize the cathelicidin from buffalo (Bubalus bubalis) uterine epithelia to elucidate its potency to use as blue print for novel antimicrobial agents.
MATERIALS AND METHODS
Sample collection, RNA extraction and RT-PCR
A total of five numbers of uteri was collected from apparently healthy buffalo based on the observation and pre rectal clinical examination before slaughter from local abattoir in ice after washing by chilled phosphate buffer saline (pH 7.4). RNA was isolated using TRI Reagent TM (Sigma-Aldrich, USA) following manufacturer protocol. The purity and integrity of RNA was checked spectrophotometrecally (A260/A280) and 1% agarose gel electrophoresis, respectively. Reverse transcription of uterus RNA was done with specific primer designed from published conserve sequences (Accession No.NM_174001, Accession No. NM_174150, Accession No. NM_174831, Accession No.174827) and oligonucleotide sequences for forward 5/GGACCATGCAGACCCAGA 3/ and reverse primers were 5/TGTCCAGAAGCCCGAATC3/). One-Step RT-PCR kit (Qiagen Inc., Chatsworth, CA) was used to carry out the reverse transcription as well as amplification in a single step. The reaction mixture was prepared by adding 5x RT-PCR buffer 10 µl, 10 mM dNTP mixture 2 µl, primer both forward and reverse (20 pmoles/µl) 2 µl each, RT- PCR master enzyme mix 2 µl, template RNA (50 ng/µl) 5 µl and nuclease free water 27 µl. The different cycle condition in PCR was: reverse transcription for 30 min at 50°C, initial PCR activation for 15 min at 95°C, denaturation for 1 min at 94°C, annealing for 1 min at 52°C and extension for 45 s at 72°C (35 cycles) and final extension was done at 72°C for 5 min. 1.0% agarose gel electrophoresis was performed for confirmation of amplified PCR product and specific product was purified by ‘DNA Elution Kit’ (Sigma - Aldrich, USA.
Cloning, sequencing and in silico analysis
The purified PCR product was ligated to pGEM-T Easy (Promega, Madison, USA) cloning vector and transformed into freshly prepared E. coli DH5a competent cells (Chung et al., 1989). Plating was done on LB agar containing ampicillin (50mg/ml), IPTG and X-gal (25 mg/ml). The plates were incubated for overnight at 37°C and white recombinant colonies were picked up after completion of incubation period. Plasmids were isolated (Sambrook and Russel, 2001) from the overnight grown culture and insert release was confirmed by EcoR1 digestion. The positive recombinant plasmids of five clones were sequenced and analyzed using Lasergene Software (DNA Star, USA) with other published sequences. The amino acid sequence was predicted and the amino acid distribution in different domain was analyzed using clustal W analysis programme (DNA Star, USA).
RESULTS
The yield of isolated total RNA from epithelial layer of uterus was 24.6 µg per mg of tissue and A260/A280 was 11.87. All the three bands (28S, 18S and 5S) of RNA was revealed in 1% agarose gel electrophoresis. RT-PCR product of total RNA gave a specific product of approximately 516 bp of cathelicidin gene. The purified product was cloned in pGEM-T Easy (Promega, Madison, USA) cloning vector (Figure 1).
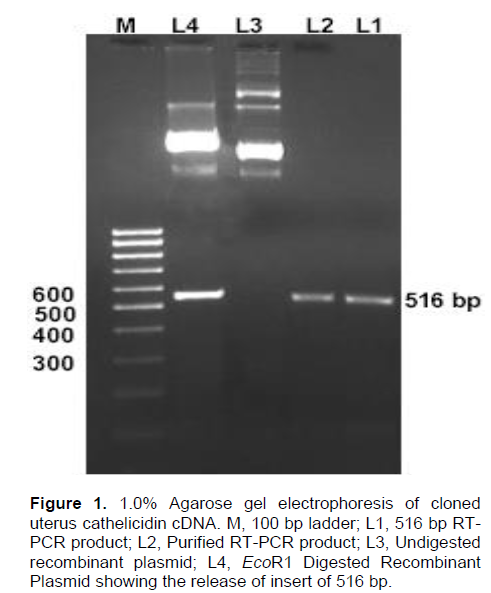
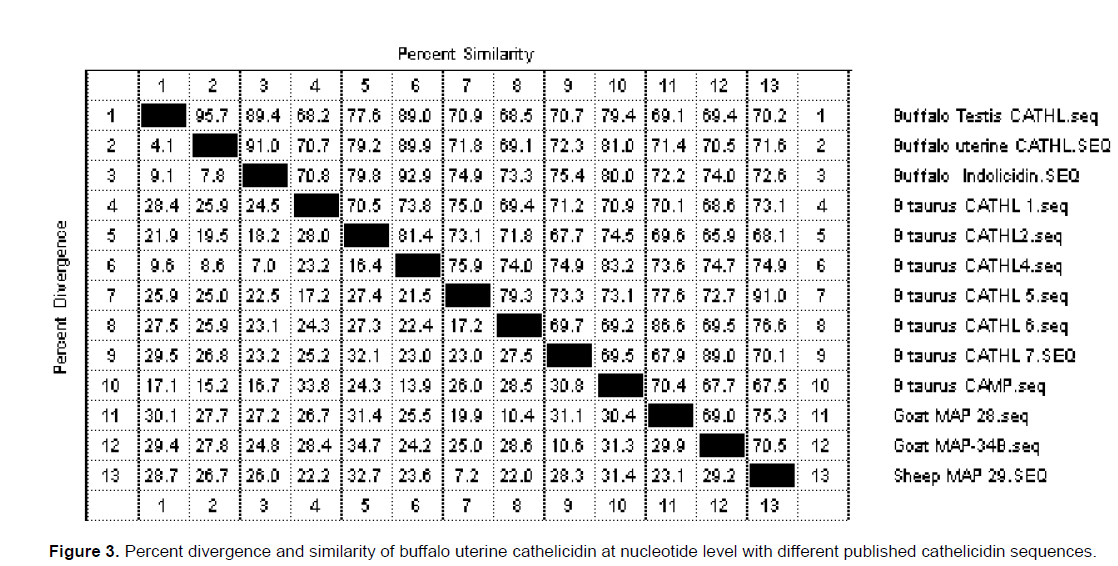
The sequence result of uterus cathelicidin has been submitted to NCBI gene data bank and Accession No. EF 050433 has been offered for the sequence). Buffalo uterus cathelicidin cDNA have full length ORF region from 6-452 and code prepropeptide of 148 amino acids (Figure 2). The ORF region of buffalo uterus and testis (Accession No. DQ832665) cathelicidin differs by 18 nucleotides at 8, 20, 25, 26, 28, 84, 108, 147, 179, 183, 186, 203, 276, 277, 284, 377, 416 and 436. The buffalo myeloid indolicidin (AJ812216) was varied by 32 nucleotides with buffalo uterus cathelicidin sequence. At nucleotide level, uterine cathelicidin showed similarity of 95.7% and 91.0% with testis cathelicidin and indolicidin, respectively (Figure 3). Similarly, buffalo uterine cathelicidin shared 70.7, 79.2, 89.9, 71.8, 69.1, 72.3 and 81.0% of identity with Bos taurus CATHL1, 2, 4, 5, 6, 7 and

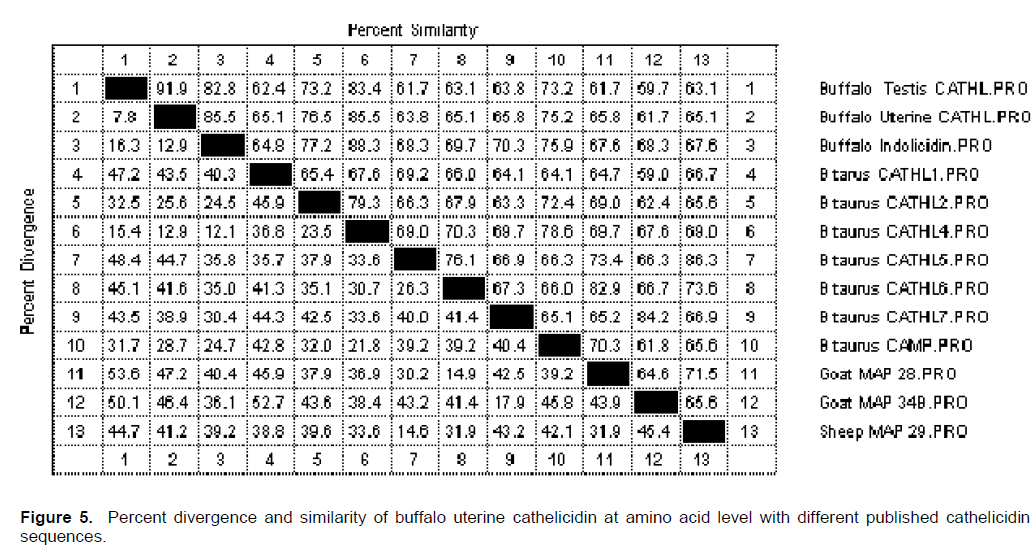
The predicted prepropeptide of buffalo uterus cathelicidin precursor peptide contain 20 strongly basic, 19 strongly acidic, 48 hydrophobic and 42 polar amino acids. The amino acid alignment with different buffalo cathelicidin congeners revealed 120 conserved amino acids (Figure 4). Four additional amino acids (132 to 135) were observed in both uterine cathelicidin as compared to buffalo indolicidin. The deduced amino acid sequences of cathelicidin showed 31 conserved amino acids with different species of animals. The percentage similarity and divergence of uterine cathelicidin with publish sequences at amino acid level is depicted in the Figure 5. The uterine cathelicidin had 91.9% amino acid similar with testis and 85.5% with buffalo indolicidin. Similarly, the amino acid sequence of uterine cathelicidin shared a similarity of 65.1, 76.5, 85.5, 63.8, 65.1, 65.8 and 75.2% with Bos taurus CATHL 1, 2, 4, 5, 6, 7 and CAMP, respectively.

DISCUSSION
The nucleotide sequence of uterine cathelicidin showed a highly conserve 5' nucleotides region and a limited similarity in 3' region with other published sequences, which is the common structural features of the cathelicidin gene (Ganz et al., 1989; Storici and Zanetti, 1993). The deduced prepropeptides of uterus cathelicidin cDNA had conserved N-terminal and diverse heterogeneous C-terminal was in agreement with other cathelicidin congeners (Popsueva et al., 1996; Wu et al., 1999). Alanine at 29 was conserved in most of the congeners and it might be the probable site of elastase mediated proteolytic cleavage to separate the signal sequence from the prosequence. The signal sequence of uterine cathelicidin comprised of 1-29 hydrophobic stretch of amino acid residues and corroborated with other congeners (Storici et al., 1992; Das et al., 2006). Valine at 130 was common to all most all prepropeptide including uterine cathelicidin indicating the common processing sites to yield the mature carboxyl terminal peptides (Storici and Zanetti, 1993; Skerlavaj et al., 1996). The prosequence of uterine cathelicidin comprised of 101 amino acid residues from 30 to 130 residues, which are highly identical to the cathelin motif, an inhibitor of thiol proteases (Ritonja et al., 1989). The cathelin like prosequence is the hallmark for all cathelicidin as it prevents the tissue injury and inflammation by neutralizing the high cationic charge of mature peptides by the presence of clusters of negatively charged amino acids (Storici et al., 1992; Selested et al., 1992; Ganz, 1989). The cathelin domains also have anti-protease activity and thus confer stability during storage (Ganz, 1989).
Uterine cathelicidin had 18 amino acid residues in the mature peptide from 131 to 148. The glycine was the last residues in most of the congeners along with the uterine cathelicidin and it might probably act as amide donor in post translation amidation of the C-terminus (Skerlavaj et al., 1996; Bradbury, 1991). The C-terminal amidation is essential for its optimum antimicrobial activity which increases the lipolysaccharide binding ability and enhances the outer membrane permeabilization (Sitaram and Nagaraj, 1999; Timothy and Roberte, 1997, Peters et al., 2010). The C-terminal amidation of peptides provides an additional hydrogen bond for α-helix stabilization (Dennison et al., 2005). The amino acid sequence of uterine cathelicidin had 4 extra stretch of amino acids namely arginine, phenylalanine, proline and tryptophan from 133 to 136, respectively. Thus, the mature peptide of uterine cathelicidin had 5 arginine, 5 tryptophan and 4 proline. The positively charged arginine is essential to initiate interaction with negatively charged outer bacterial surface (Timothy and Roberte, 1997; Boman, 1991). The tryptophan influences the localization of these peptides in to membrane interfaces (Schiffer et al., 1992).
Proline is an important amino acid that enhances the microbicidal activity by forming flexible helical kink and more ordered structure, which increases membrane permeability (Park et al., 2002). Besides, prolines also provide some protection against nonspecific proteolytic degradation of proteases secreted by microorganisms (Vanhoof et al., 1995). Peptide rich in arginine, tryptophan and proline are shown to have strong microbicidal activity against gram positive and gram negative bacteria (Selested et al., 1992), fungi (Subbalakshmi and Sitaram, 1998) and virus (Robinson (jr) et al., 1998). Expression of antimicrobial peptide with high number of arginine along with tryptophan and proline by buffalo uterine epithelium help this species to be free from several infectious reproductive disorders as compared to cattle and other species. The mature peptide domain had more variation compared to other region and it is attributed as, genes involved in immunity and host defense is rapidly diverged and had undergo positive selection to fight competently in antigerm war (Emes et al., 2003; Crovella et al., 2005).
CONCLUSION
From the present study it can be concluded that, mature amino acids sequence of uterine cathelicidin is comprise of 5 arginine, 5 tryptophan and 4 proline residues and the domain can serve as blue print for synthesis of a novel antimicrobial agent.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
REFERENCES
|
Bals R (2000). Epithelial antibacterial peptides in host defense against infection. Resp. Res. 1:141-150. |
|
|
Boman HG (1991). Antibacterial peptides: Key components needed in immunity. Cell, 65: 205-207. |
|
|
Bradbury AF, Smyth DG (1991). Peptide amidation. Trend. Biochem. Sci. 16:112-115. |
|
|
Chung CT, Niamela SL, Miller RH (1989). One-step preparation of competent Eschericia coli: transformation and storage of bacterial cells in the same solution. Proced. Natur. Acad. Sci. 86:2172-2175. |
|
|
Crovella S, Antcheva N, Zelezetsky I, Boniotto M, Pacor S, Vittoria M, Tossi A (2005). Primate β-defensins structure-function and evolution. Curr. Pro. Pept. Sci. 6:7-21. |
|
|
Das H, Sharma B, Kumar A (2006). Cloning and characterization of novel cathelicidin cDNA sequence of Bubalus bubalis homologus to Bos taurus cathelicidins- 4. DNA Seq. 17:407-414. |
|
|
Dennison SR, Wallace J, Harris F, Phoenix D (2005). Amphiphilic α-Helical Antimicrobial Peptides and Their Structure/ Function Relationships. Pro. Pept let. 12:31-39. |
|
|
Emes RD, Goodstadt L, Winter E, Ponting CP (2003). Comparison of genomes of human and mouse lays the foundation of genome zoology. Hum. Mol. Gen. 12:701-709. |
|
|
Ganz T, Lehrer R (1989). Antibiotics peptides from higher eukaryotes: biology and applications. Mol. Med. Today. 5:292-297. |
|
|
Ganz T, Rayner JR, Valore EV, Tumolo A, Talmadge K, Fuller F (1989). The structure of the rabbit macrophage defensin genes and their organ-specific expression. J. Immunol. 143:1358-1365. |
|
|
Garica JR, Krause A, Scchulz S (2001). Fimpong-Boateng A. Human beta defensin 4: a novel inducible peptide with a specific salt-sensitive spectrum of antimicrobial activity. Feder. Americ. Soc. Exp. Biol. Lett. 15:1819-1821. |
|
|
Gee ML, James, AG, Hossain MA, McArthur S, Palombo EA, Wade JD, Clayton AH (2013). Imaging the action of antimicrobial peptides on living bacterial cells. Sci. Reports. 3:1557 |
|
|
Gorbach SL (2001). Antimicrobial use in animal feed: ime to stop. N. Engl. J. Med. 345:1202-1203. |
|
|
Grandjean V, Vincent S, Martin L, Rassoulzadegan M, Cuzin F (1997). Antimicrobial protection of the mouse testis : Synthesis of defensin of the cryptdin family. Biol. Reprod. 57:1115-1122. |
|
|
Li P, Chang Chan H, He Bin, Cheung SOS, Chung YW, Shang Q, Zhang Y, Zhang Y (2001). An antimicrobial peptide gene found in the male reproductive system of rats. Sci. 291:1783-1785. |
|
|
Malm J, Sorenson O, Persson T, Nilsson F, Johansson B, Bjartell A, Lilja H, Borregard M, Engstein AQ (2000). The human cationic Antimicrobial protein (h CAP-18) is expressed in the epithelium of human epididymis, is present in the seminal plasma at high concentration and is attached to spermatozoa. Infec. Immun. 68:4297-4302. |
|
|
Markus Reinholz MD, Thomas Ruzicka MD, Jürgen Schauber MD (2012). Cathelicidin LL-37: An Antimicrobial Peptide with a Role in Inflammatory Skin Disease. Ann. Drmato. 24:126-135. |
|
|
Park S, Kim H, Kim C, Yun H, Choi E, Lee B (2002). Role of proline, cysteine and disulphide bridge in the structure and activity of the antimicrobial peptide gaegurin5. Biochem. J. 368:171-182. |
|
|
Peters BM, Shirtliff, EM, Jabra-Rizk MA (2010). Antimicrobial Peptides: Primeval molecules or future Drugs? Pathogens 6:1-4. |
|
|
Popsueva A, Zinovjeva M, Visser J, Zijilmans J, Fibbe W, Belyavsky A (1996). A novel murine cathelin-like protein expressed in bone marrow. Feder. Am. Soci. Exp. Biol. Lett. 391:5-8. |
|
|
Ravi C, Jeyashree AR, Devi K (2011). Antimicrobial Peptides from Insects: An Overview. Res. Biotech. 2:1-7. |
|
|
Ritonja A, Kopiter M, Jerala R, Turk V (1989). Primary structure of a new cysteine proteinase inhibitor from pig leucocytes. Feder. Am. Soci. Exp. Biol. Lett. 255:211-214. |
|
|
Robinson WE (Jr), McDougall B, Tran D, Selested ME (1998). Anti-HIV-1 activity of indolicidin, an antimicrobial peptide from neutrophils. J. Leuk. Biol. 63:94-100. |
|
|
Sambrook J, Russel T (2001). Preparation of plasmid DNA by alkaline lysis with SDS :Minipreparation.Molecular Cloning:A Laboratory Manual. 3rd Edition. (Cold Spring Harbor, NY). pp. 1.32-1.34. |
|
|
Schiffer MCH, Chang CH, Stevens J (1992). The functions of tryptophan residues in membrane proteins. Pro. Eng. 5:213-214. |
|
|
Selested ME, Novotny MJ, Morris WL, Tang YQ, Smith W, Cullor JS (1992). Indolicidin, a novel bactericidal tridecapeptide amide from neutrophils. J. Biol. Chem. 267:4292-4295. |
|
|
Sitaram N, Nagaraj R (1999). Interaction of antimicrobial peptides with biological and model membranes: structural and charge requirements for activity. Biochem. Biophys. Acta. 1462:29-54. |
|
|
Skerlavaj B, Gennaro R, Bagella L, Merluzzi L, Risso A, Zanetti M (1996). Biological characterization of two novel cathelicidin-derived peptides and identification of structural requirements for their antimicrobial and cell lytic activities. J. Biol. Chem. 271:28375-28381. |
|
|
Storici P, Schneider C, Zanetti M (1992). cDNA sequence analysis of an antibiotic dodecapeptide from neutrophils. Feder. Ameri. Soci. Exp. Biol. Lett. 314:187-190. |
|
|
Storici P, Zanetti M (1993). A cDNA derived from pig bone marrow cells predicts a sequence identical to the intestinal antibacterial peptide, pR-39. Biochem Biophy. Res. Commun. Sci. 196:1058-1065. |
|
|
Subbalakshmi C, Sitaram N (1998). Mechanism of antimicrobial action of Indolicidin FEMS Microbiol. Lett. 160: 91-96. |
|
|
Timothy J, Robert EWH (1997). Improved Activity of a Synthetic Indolicidin Analog. Antimicrob. Agent. Chemo. 41: 771-775. |
|
|
Vanhoof G, Goossens F, Meester ID, Hendriks D, Scharpe S (1995). Proline motifs in peptides and their biological processing. Feder. Am. Soci. Exp. Biol. J. 9:736-744. |
|
|
Wu M, Maier E, Benz R, Hancock RE (1999). Mechanism of different classes of cationic antimicrobial peptides with planer bilayers and with the cytoplasmic membrane of Escherichia coli. Biochem. J. 38:7235-7248. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


