Full Length Research Paper

ABSTRACT
Fusaria are very diverse and destructive pathogens affecting different crops. However, their identity and diversity are unresolved in countries like Ethiopia, where various crop species are grown under differing environmental conditions. The objectives of this paper were to identify Fusarium spp. associated with sorghum stalk rot in Southern Ethiopia, and elucidate the genetic diversity within and between the species. For this purpose, Fusaria associated with sorghum from two locations in Southern Ethiopia were isolated. Sequencing of the elongation factor 1-alpha gene (EF-1α) was used for species identification. In addition, AFLP analysis was employed for further diversity studies within and between the Fusarium spp. Sequence analyses revealed the presence of two Fusarium spp. The first was identified as Fusarium andiyazi, while the identity of the second remains to be solved. AFLP analysis clustered the isolates into two major groups. The Dice similarity coefficients ranged from 0.39 to 0.91 for isolates of F. andiyazi while isolates within the new Fusarium spp. had a Dice similarity coefficient varying between 0.69 and 0.96. Cluster analysis and principal coordinate analysis clearly indicated a genetic separation between the two species. Both groups were pathogenic to mature sorghum plants following a toothpick inoculation test. More researches are required to identify the new species and elucidate the pathogenicity of the isolates.
Key words: EF-1α, Fusarium andiyazi, genetic similarity, sequence analysis, Sorghum bicolor.
INTRODUCTION
MATERIALS AND METHODS
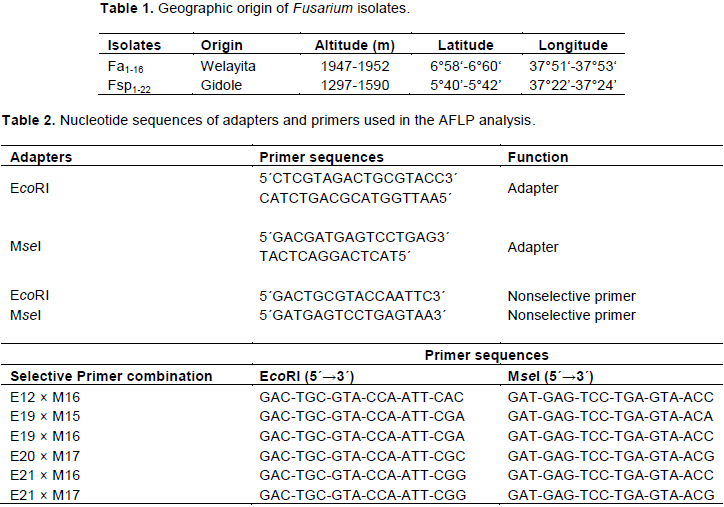
RESULTS
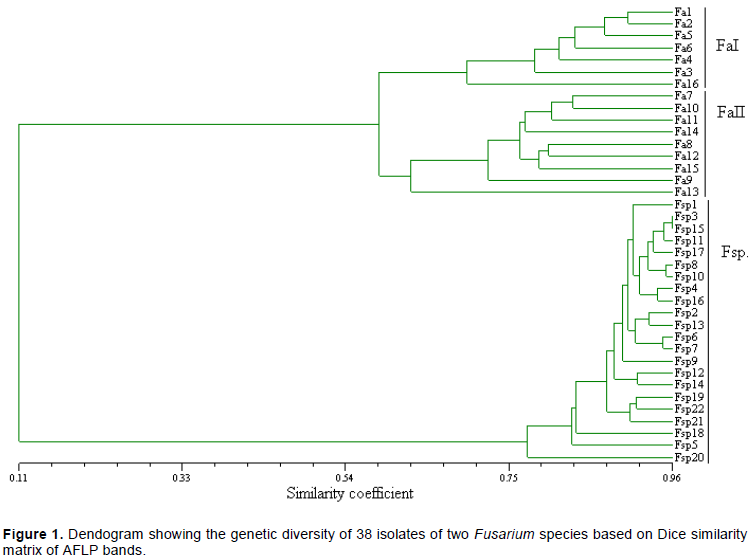
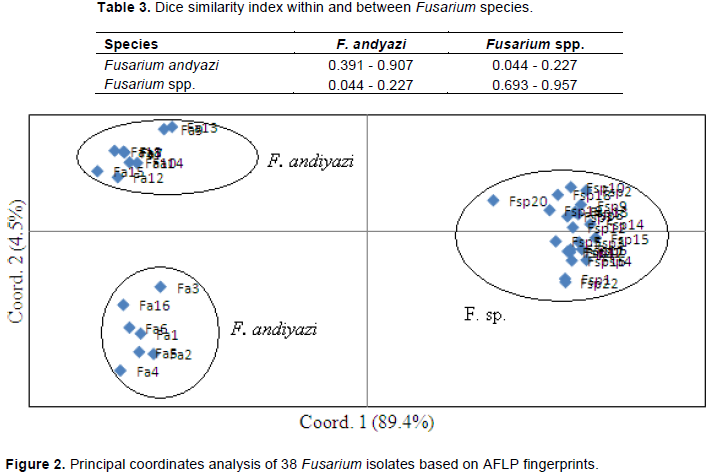
DISCUSSION
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
ACKNOWLEDGEMENT
REFERENCES
|
Antonia S (1995). The occurrence and biology of some Fusarium spp., on wheat in Slovakia. Institute of Experimental Phytopathology and Entomology. Slovak Academy of Sciences, Ivanka pri Dunaji, Slovakia. 119 p. |
|
|
Antonissen G, Martel An, Pasmans F, Ducatelle R, Verbrugghe E, Vandenbroucke V, Li S, Haesebrouck F, Immerseel FV, Croubels S (2014). The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 6:430-52. |
|
|
Ayalew A (2002). Mycoflora and mycotoxins of major cereal grains and antifungal effects of selected medicinal plants from Ethiopia. Ph.D. Dissertation]. Goettingen, Germany: Georg August University. |
|
|
Ayalew A, Fehrmann H, Lepschy J, Beck R, Abate D (2006). Natural occurrence of mycotoxins in staple cereals from Ethiopia. Mycopathologia 162:57-63. |
|
|
Backhouse D, Burgess LW, Summerell BA (2001). Biogeography of Fusarium, p. 122-137. In: Summerell BA, Leslie JF, Backhouse D, Bryden WL, Burgess LW. (Eds.), Fusarium: Paul E. Nelson Memorial Symposium. APS Press, St. Paul, Minnesota. |
|
|
Berenji J, Dahlberg J (2004). Perspectives of sorghum in Europe. Journal of Agronomy and Crop Science 190(5):332-338. |
|
|
Brandfass C, Karlovsky P (2008). Upscaled CTAB-Based DNA Extraction and Real-Time PCR Assays for Fusarium culmorum and F. graminearum DNA in Plant Material with Reduced Sampling Error. International Journal of Molecular Sciences 9(11):2306-2321. |
|
|
Browning M, Rowley LV, Zang P, Chandlee JM, Jackson N (1999). Morphological, pathogenic and genetic comparisons of Colletotrichum graminicola isolates from Poaceae. Plant Disease 83(3):286-92. |
|
|
Chala A, Tronsmo AM, Brurberg MB (2011). Genetic differentiation and gene flow in Colletotrichum sublineolum in Ethiopia, the centre of origin and diversity of sorghum, as revealed by AFLP analysis. Plant Pathology 60(3):474-482. |
|
|
Chala A, Tronsmo AM (2012). Evaluation of Ethiopian sorghum accessions for resistance against Colletotrichum sublineolum. European Journal of Plant Pathology 132(2):179-189. |
|
|
Chala A, Taye W, Ayalew A, Krska R, Sulyok M, Logrieco A (2014). Multimycotoxin analysis of sorghum (Sorghum bicolor L. Moench) and finger millet (Eleusine coracana L. Garten) from Ethiopia. Food Control 45:29-35. |
|
|
Claflin LE (2000). Fusarium root and stalk rot. In: Frederiksen RA, Odvody GN (eds.). Compendium of Sorghum Diseases. The American Phytopathology Society. St. Paula MN USA. pp 29-30. |
|
|
Crouch JA, Clarke BB and Hillman BI (2006). Unraveling evolutionary relationships among divergent lineages of Colletotrichum causing anthracnose disease in turfgrass and corn. Phytopathology 96(1):46-60. |
|
|
Central Statistical Agency (CSA) (2018). Agricultural Sampling Survey. Report on area and production of crops. http://www.csa.gov.et. Accessed February 2019. |
|
|
Cumagun CJR, Ramos JS, Dimaano AO, Munaut F, Van Hove F (2009). Genetic characterization of Fusarium verticilloides from corn in the Philippines. Journal of General Plant Pathology 75:405-412. |
|
|
Dice LR (1945). Measures of the amount of ecologic association between species. Ecology 26(3):297-302. |
|
|
D'Mello JPF, Placinta CM, Macdonald AMC (1999). Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Animal Feed Science and Technology 80(3-4):183-205. |
|
|
De Wolf ED, Madden LV, Lipps PE (2003). Risk assessment models for wheat Fusarium head blight epidemics based on within-season weather data. Phytopathology 93(4):428-435. |
|
|
Duan C, Qin Z, Yang Z, Li W, Sun S, Zhu Z, Wang X (2016). Identification of pathogenic Fusarium spp. causing maize ear rot and potential mycotoxin production in China. Toxins 8(6):186. |
|
|
Eshte Y, Mitiku M, Shiferaw W (2015). Assessment of important plant disease of major crops (Sorghum, Maize, Common Bean, Coffee, Mung bean, Cowpea) in south Omo and Segen Peoples Zone of Ethiopia. Current Agriculture Research Journal 3(1):75-79. |
|
|
Food and Agriculture Organization of the United Nations (FAO) (2017). FAOSTAT-Crop production data. http://faostat.fao.org. Accessed February 2019. |
|
|
Frederiksen RA, Odvody GN (2000). Compendium of Sorghum Diseases. 2nd ed. The American Phytopathological Society, St. Paul, MN. |
|
|
Laurence MH, Walsh JL, Shuttleworth LA, Robinson DM, Johansen RM, Petrovic T, Vu TTH, Burgess LW, Summerell BA, Liew ECY (2016). Six novel species of Fusarium from natural ecosystems in Australia. Fungal Diversity77(1):349-66. |
|
|
Laraba I, Boureghda H, Abdallah A, Bouaicha O, Obanor F, Moretti A, Geiser DM, Kim HS, McCormick SP, Proctor RH, Kelly AC, Ward TJ, O'Donnell K (2017). Population genetic structure and mycotoxin potential of the wheat crown rot and head blight pathogen Fusarium culmorum in Algeria. Fungal Genetics and Biology 103:34-41. |
|
|
Leavitt SD, Esslinger TL, Spribille T, Divakar PK, Thorsten LH (2013). Multilocus phylogeny of the lichen-forming fungal genus Melanohalea (Parmeliaceae, Ascomycota): insights on diversity, distributions, and a comparison of species tree and concatenated topologies. Molecular Phylogenetics and Evolution 66(1):138-152. |
|
|
Leslie JF, Zeller KA, Lamprecht SC, Rheeder JP, Marasas WFO (2005). Toxicity, pathogenicity, and genetic differentiation of five species of Fusarium from sorghum and millet. Phytopathology 95(3):275-283. |
|
|
Leslie JF (2000). Fusarium species associated with sorghum. In: Frederiksen RA, Odvody GN. (eds.). Compendium of Sorghum Diseases. The American Phytopathology Society. St. Paula. MN. USA P 30. |
|
|
Leslie JF (2002). Sorghum and pearl millet diseases in the horn of Africa. In: Leslie JF (ed.). Sorghum and millet diseases. Blackwell publishing. Iowa State Press pp. 383-387. |
|
|
Leslie JF, Summerell BA (2006). The Fusarium laboratory manual. Blackwell, IA, P 388. |
|
|
Maphosa MN, Steenkamp ET, Wingfield BD (2016). Genome-based selection and characterization of Fusarium circinatum-specific sequences. G3 (Bethesda) 6(3):631-639. |
|
|
Marasas WFO, Rheeder JP, Lamprecht SC, Zeller KA, Leslie JF (2001). Fusarium andiyazi sp. nov., a new species from sorghum. Mycologia 93(6):1203-1210. |
|
|
Marley P, Marasas WFO, Hester V (2004). Occurrence of Fusarium andiyazi associated with sorghum in Nigeria. Archives of Phytopathology and Plant Protection 37(3):177-181. |
|
|
McDonald MC, Razavi M, Friesen TL, Brunner PC, McDonald BA (2012). Phylogenetic and population genetic analyses of Phaeosphaeria nodorum and its close relatives indicate cryptic species and an origin in the fertile crescent. Fungal Genetics and Biology 49(11):882-895. |
|
|
Minnaar-Ontong A, Herselman L, Kriel WM, Leslie JF (2017). Morphological characterization and trichothecene genotype analysis of a Fusarium Head Blight population in South Africa. European Journal of Plant Pathology 148(2):261-269. |
|
|
Moschini RC, Carranza MR, Carmona MA (2004). Meteorological-based predictions of wheat head blight epidemics in the Southern Argentinian Pampas region. Cereal Research Communications 32(1):45-52. |
|
|
Moussa TAA, Al-Zahrani HS, Kadasa NMS, Ahmed SA, de Hoog GS, AlHatmi AMS (2017). Two new species of the Fusarium fujikuroi species complex isolated from the natural environment Antonie van Leeuwenhoek 110(6):819-832. |
|
|
O'Donnell K, Ward TJ, Robert VARG, Crous PW, Geiser DM, Seogchan K (2015). DNA sequence-based identification of Fusarium: current status and future directions. Phytoparasitica 43(5):583-595. |
|
|
Parry DW, Jenkinson P, Mc Leod L (1995). Fusarium ear blight (scab) in small grain cereals – a review. Plant Pathology 44(2):207-238. |
|
|
Peakall R, Smouse PE (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes 6(1):288-295. |
|
|
Pestka JJ, Smolinski AT (2005). Deoxynivalenol: Toxicology and potential effects on humans. Journal of Toxicology and Environmental Health, Part B 8(1):39-69. |
|
|
Ramdial H, Latchoo RK, Hosein FN, Rampersad SN (2017). Phylogeny and haplotype analysis of fungi within the Fusarium incarnatum-equiseti species complex. Phytopathology 107(1):109-20. |
|
|
Rivera-Vargas LI, Lugo-Noel Y, McGovern RJ, Seijo T, Davis MJ (2006). Occurrence and distribution of Colletotrichum spp. on mango (Mangifera indica L.) in Puerto Rico and Florida, USA. Plant Pathology Journal 5(2):191-98. |
|
|
Saremi H, Burgess LW, Backhouse D (1999). Temperature effects on the relative abundance of Fusarium species in a model plant-soil ecosystem. Soil Biology and Biochemistry 31(7):941-47. |
|
|
Summerell BA, Leslie JF, Liew ECY, Laurence M, Bullock S, Petrovic T, Bentley AR, Howard CG, Peterson SA, Walsh JL, Burgess LW (2011). Fusarium species associated with plants in Australia. Fungal Diversity 46(1):1-27. |
|
|
Taye W, Ayalew A, Chala A, Dejene M (2016). Aflatoxin B1 and total fumonisin contamination and their producing fungi in fresh and stored sorghum grain in East Hararghe, Ethiopia. Food Additives and Contaminants: Part B Surveillance 9(4):237-245. |
|
|
Taye W, Ayalew A, Dejene M, Chala A (2018). Fungal invasion and mycotoxin contamination of stored sorghum grain as influenced by threshing methods. International Journal of Pest Management 64(1):66-76. |
|
|
Thakur RP, Mathur K (2000). Anthracnose: Compendium of sorghum diseases. In Frederiksen RA, Odvody GN (Eds.). The American Phytopathological Society, St. Paul MN. pp. 10-12. |
|
|
Valente MT, Desideriob F, Infantinoa A, Vale G, Abbruscatod P, Aragona M (2017). Genetic variability of Fusarium fujikuroi populations associated with bakanae of rice in Italy. Plant Pathology 66(3):469-79. |
|
|
Van der Lee T, Zhang H, Van Diepeningen A, Waalwijk C (2015). Biogeography of Fusarium graminearum species complex and chemotypes: a review. Food additives and contaminants. Part A. Chemistry, analysis, control, exposure & risk assessment 32(4):453-460. |
|
|
Vigier B, Reid LM, Seifert KA, Stewart DW (1997). Distribution and prediction of Fusarium species associated with maize ear rot in Ontario. Canadian Journal of Plant Pathology 19(1):60-65. |
|
|
Vos P, Hogers R, Bleeker M, Rijans M, Van de Lee T, Hornes M, Frijters A, Pot J, Kuiper M, Zebeau M (1995). AFLP: A new technique for DNA fingerprinting. Nucleic Acids Research 23(21):4407-4414. |
|
|
Wollenweber HW, Reinking OA (1935). Die Fusarien, ihre Beschreibung, Schadwirkung und Bekämpfung. Paul Parey, Berlin, Germany. |
|
|
Wu F, Groopman JD, Pestka JJ (2014). Public health impacts of foodborne mycotoxins. Annual Review of Food Science and Technology 5:351-372. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


