Full Length Research Paper

ABSTRACT
Antibiotics are secondary metabolites produced by microorganisms. Actinomycetes are the top producers of antibiotics. The study was designed to isolate and screen antibiotic producing actinomycetes obtained from rhizosphere of plants and agricultural soils. A total of 30 actinomycetes were isolated. Out of the 30 actinomycetes, 18 (60%) showed antimicrobial activities against one of the tested microorganisms of which 9 isolates were selected for their wide spectrum of antibiotic activities. In this study, isolates AAUBA5 and AAUBA30 were found to inhibit all of the test organisms, except Escherichia coli ATCC 25922. The latter was found to be the most resistant and was only inhibited by 3/18 (17%) of the isolates. The isolates were identified and characterized by cultural, morphological, physiological, and biochemical characteristics and found to be classified under the genus Streptomycetes. In order to extract the active compounds from isolates, fermentation was carried out on 9 isolates and the former was extracted by using ethyl acetate. Finally, isolate AAUBA13 was selected for further study based on spectrum activity from disc diffusion assay. Antimicrobial compound obtained from AAUBA13 isolate was separated and purified with thin layer chromatography (TLC), column chromatography, and preparative chromatography. The finding of this study, in general, showed that, antimicrobial compounds obtained from AAUBA13 demonstrate broad spectrum antimicrobial activity against Staphylococcus aureus ATCC25923, E. coli ATCC 25922, Pseudomonas aeruginosa ATCC25853, S. aureus (clinical isolate), Shigella boydii (clinical isolate), Streptococcus pneumonia ATCC49619, and Candida albicans ATCC62376.
Key words: Actinomycetes, antibiotics, antimicrobial compound, disc diffusion assay, soil.
INTRODUCTION
Antibiotics are chemical substances produced by microorganisms, which in small amount selectively inhibit or kill other microorganisms (Aneja, 2005; Tortora et al., 2010). They are secondary metabolites that inhibit other competing cells to give competitive advantage to the microorganisms that produce them (Sanglier et al., 1993; Kumbhar and Watve, 2013).
Antimicrobial agents are natural products and produced by various types of bacteria and fungi. Hundreds of these natural products have been identified and developed as therapeutic agents against many infectious diseases (Berdy, 2005). Microbial natural metabolites still appear as the most promising sources of antibiotic in the future (Fernando, 2006; Wohlleben et al., 2016). Some of the important antibiotic producing microorganisms are of Streptomyces, Bacillus, Cephalosporium, and Penicillium that have been studied continuously for their ability to produce antibiotics (Brock and Madigan, 1991).
From all the known microbes, actinomycetes are the most important source of biologically active microbial products, including many medically and commercially important antibiotics (Dhanasekaran et al., 2009; Adegboye and Babalola, 2013). The broad-spectrum antibiotic, vancomycin, a potent antimicrobial agent against methicillin resistant Staphylococcus aureus and rifampcin, and effective drug against tuberculosis and leprosy is derived from several species of actinomycetes (Berdy, 2005).
The genus Streptomyces has been widely recognized as industrially important microorganism because of its ability to produce and secrete a large variety of secondary metabolites (Kornwendisch and Kutzner, 1992; Saadoun and Gharaibeh, 2003; Pandey et al., 2004). These include aminoglycosides, macrolides, β lactams, peptides, polyenes, tetracyclines, anthracycline antibiotics, and nucleosides (Vijayakumar et al., 2007; Miyadoh, 1993). It is estimated that more than 80% of the antibiotics are obtained from Streptomycetes (Vijayakumar et al., 2007). Micromonospora is the runner up with less than one-tenth Streptomyces (Saadoun and Gharaibeh, 2003; Arifuzzaman et al., 2010).
Although the introduction of antimicrobials helps to combat many diseases, large numbers of pathogenic bacteria and fungi causing different human diseases have become resistant to antibiotics in use (Livermore, 2003). S. aureus, a virulent pathogen, which is responsible for a wide range of infections, including pimples, pneumonia, osteomyelitis, endocarditis and bacteremia, has developed resistance to most classes of antibiotics (Enright, 2003). The increase in antibiotic resistance has been attributed to a combination of microbial characteristics, selective pressure of antibiotic use, and social changes that enhance the transmission of resistant organisms (Okeke et al., 2005).
With increase in misuses of antibiotics, the evolution of emergent and reemerging antibiotic resistant disease is developing at an alarming rate (Demain and Elander, 1999). In recent years, new resistant strains emerge more quickly while the rate of discovery of new antibiotics is slowing down (Oskay et al., 2004; Parungao et al., 2007). This necessitates the screening of micro-organisms for antimicrobial activity for the production of new and novel drugs (Singh et al., 2016). Hence, intensive search for new antibiotics has become imperative worldwide especially from new actinomycetes (Oskay et al., 2004; Parungao et al., 2007). In addition, it has been reported that screening programs using highly selective procedure allows the detection and isolation of effective antibiotic producing microorganisms from soil (Rondon et al., 2000; Oskay et al., 2004; Parungao et al., 2007). Soil is the largest source of microorganisms and a natural reservoir for microorganisms (Dancer, 2004; Hackl et al., 2004).
Although soils have been screened by the pharmaceutical industry for about 50 years, only a small fraction of the surface of the earth has been sampled and only a small fraction of actinomycetes taxa has been discovered (Baltz, 2007). This requires the employment of several strategies to explore new compounds from microorganisms such as actinomycetes from different ecological niches that may yield novel compounds with diverse antimicrobial properties (Pandey et al., 2004; Ningthoujam et al., 2009).
In Ethiopia, a few investigators showed the existence of antibiotic producing microorganisms from different ecosystems. Biniam (2008) isolated antimicrobial producing actinomycetes from southern part of Ethiopian Rift Valley alkaline lakes. The potential of mushroom compost as a good source of antibiotic producing thermophilic actinomycete was also reported by Moges (2009). However, there is a further need to explore indigenous actinomycetes for their potential to produce effective antibiotics from different sources in the country to combat infectious diseases. Hence, this study aimed to isolate, screen, and characterize effective antibiotics producing actinomycetes and evaluate their potential against some test microorganisms.
MATERIALS AND METHODS
Soil sample collection
A total of 15 soil samples were collected from the rhizosphere of plants and agricultural soils from field sites of Deberizit (5 samples) and Holeta (5 samples) and garden soil from the College of Natural Sciences (5 samples) in December 2010. The soils were excavated from depth of 5 to 15 cm by using sterile spatula and collected in clean, dry and sterile polyethylene bags. All samples were labeled, transported to Addis Ababa University, Applied Microbiology Laboratory and stored in the refrigerator at 4°C for further investigations.
Media for the cultivation of actinomycetes
For isolation of actinomycetes, the following two media were used: starch casein agar medium (SCA, g/L) containing soluble starch 10; casein, 0.3; KNO3 2, NaCl 2, K2HP04 2, MgSO4.7H2O 0.05, CaCO3 0.02, FeSO4.7H2O 0.01; agar, 15; and pH was adjusted to 7.0±2 before sterilization (Arifuzzaman et al., 2010) and actinomycetes isolation agar (AIA) medium of g/L containing heart infusion broth, 25.0; casein hydrolysate, 4.0; yeast extract, 5.0; dextrose, 5.0; cysteine HCl, 1.0; soluble starch, 1.0; potassium phosphate, 15.0; ammonium phosphate, 1.0; magnesium sulphate, 0.2; calcium chloride, 0.02; Agar, 20; and pH was adjusted to 7 before sterilization (Awad et al., 2009). The two media were autoclaved at 121°C for 15 min; cooled up to 50°C and supplemented with 50 μg/ml of amphotericin B to minimize fungal growth; 20 to 25 ml of media was poured on sterile Petri dishes allowed to cool at room temperature.
Isolation, maintenance and designation of isolates
Actinomycetes were isolated by serial dilution plate technique (Arifuzzaman et al., 2010). About 1 g of each soil sample was suspended in 10 ml of sterile normal saline (0.85%) and shaken on orbital shaker for about 30 min at 121 rpm; the suspension was left for 30 min in the Laminar Air Flow hood. Consequently, 1 ml of suspension was taken and diluted 7-fold in 9 ml of sterile normal saline (0.85%). The suspensions were agitated with vortex and 0.1 ml of suspension was taken from 10-3, 10-5, and 10-7 dilutions and spread on each of the Starch Casein Agar medium and Actinomycetes Isolation Agar medium. The plates were incubated at 30°C for 7 to 10 days (Dhanasekaran et al., 2009). All isolates were identified as actinomycetes based on colony morphology and color of mycelium (Williams and Cross, 1971). The isolates were further sub-cultured to ensure their purity and maintained on starch casein agar medium. The inoculated agar media or slant was incubated at 30°C for 7 days and then stored at 4°C for further use. Code was assigned (AAUBA1 to AAUBA31) for every entity to confirm the source of the organism and the color of aerial mycelium.
Screening of antimicrobial activity of actinomycetes isolates against test microorganisms
Test microorganisms
The test organisms used for antimicrobial screening were: S. aureus ATCC25923, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 25853, S. aureus (clinical isolate), Shigella boydii (clinical isolate), Streptococcus pneumonia ATCC 49619, Sallmonella typhi ATCC 6539, Candida albicans ATCC 62376 and Cryptococcus neoformance (clinical isolate). The test bacteria and fungi were obtained from Ethiopian Health and Nutrition Research Institute (EHNRI) and Biomedical Laboratory, Faculty of Life Sciences, AAU.
Turbidity standard for inoculum preparation
Standardization of the inoculum density of isolates for susceptibility test was done by the method described in Lalitha (2004). In order to determine the active phase of test organisms, each isolate was grown in 100 ml of nutrient broth for bacteria and Sabourouds broth for fungi in 250 ml Erlenmeyer flask on a rotary shaker at 120 r/min and 37°C. Samples were taken every 2 h to measure optical density using spectrophotometer (JENWAY, London) at 660 nm. The optical density values were extra plotted against time to determine the different phases of the growth curve.
Samples from the exponential phase were taken to adjust the inoculum density with 0.5 McFarland Turbidity Standard prepared by adding a 0.5 ml of BaCl2 solution into 99.5 ml of solution H2SO4 (Lalitha, 2004). The density of the turbidity standard was determined using spectrophotometer (JENWAY, London) at 660 nm.
Primary screening
A total of 30 isolates were primarily screened for antimicrobial activity against five test microorganisms according to Pandey et al. (2004). Seven day grown isolates were streaked as a straight line across diameter on Nutrient Agar plates (Oxiode) and incubated at 30°C for 6 days. After 6 days, the test microorganisms, namely, S. aureus ATCC 25923, E. coli ATCC 25922, P. aeruginosa ATCC 25853, S. aureus clinical isolate and S. boydii clinical isolate from overnight culture broth were streaked at right angle, but not touching the streaked isolate and incubated at 37°C for 24 h. Clearing zone formation between the antibiotic producing isolate and the test organisms was considered positive for antibiotic production. The isolates were then selected based on a wide spectrum activity against tested microorganisms for further studies.
Secondary screening
Cultivation and extraction of cultures of actinomycetes for secondary metabolites
Nine isolates were selected for secondary screening in small scale submerged fermentation system. 200 ml of starch casein broth was dispensed into 500 ml Erlenmeyer flask, to which a loop full of seven days grown isolates were inoculated and incubated on a platform shaker (New Brunswick Scientific), at 200 rpm and room temperature for 10 days (Remya and Vijayakumar, 2008; Dhanasekaran et al., 2009). After ten days of incubation, the content of incubated flask was filtered through 0.2 µm pore size Whatman No. 1 filter paper. Equal volume of ethyl acetate (1:1) was then added to the culture filtrates and shaken vigorously for 1 h and solvent phase that presumably contains antibiotics compound was separated from aqueous phase in a separatory funnel (Assistant, Germany). The ethyl acetate phase that contains antibiotics was evaporated and concentrated in vacuum rota-vapor (BUCHI-Germany) at 100 rev/min and 60°C (Fessenden, 1993; Remya and Vijayakumar, 2008). The dry crude extract was weighted using a balance (SCALTEC, Germany) and kept in small vials at 4°C for further test.
Disc diffusion assay
Antimicrobial activity of the dried crude extract of each isolate was evaluated by using disc diffusion assay (Hassan et al., 2001; Ningthoujam et al., 2009). For this purpose, nutrient agar (NA) (Oxiod) for bacteria and Sabouraud Dextrose Agar (SDA, Oxiod) for yeast/fungi were inoculated with 0.2 ml overnight culture of each standardized test organism; S. aureus ATCC 25923, E. coli ATCC 25922, P. aeruginosa ATCC 25853, S. aureus (clinical isolate), S. boydii clinical isolate, S. pneumonia ATCC 49619, S. typhi ATCC 6539, C. albicans ATCC 62376 and C. neoformance (clinical isolate).
About 20 µl of 50 µg/ml crude extract of each isolate was impregnated with sterile Whatman 0.2 µm pore size antibiotic assay discs (6.0 mm) and placed on the inoculated agar plates. Blank discs impregnated with the solvent were also placed on inoculated plates that served as negative control. This was done in duplicates. The Petri dishes were then kept in a refrigerator at 4°C for 2 h to allow the diffusion of the extracts in the media. The Petri dishes were then incubated at 37°C for 24 h to detect and measure the inhibition diameter around the discs.
Bioassay guided purification of active compound from AAUBA13
The crude extract of AAUBA13 isolate was selected based on inhibition zone diameter and spectrum activity from antimicrobial assay test to detect the antimicrobial component of the crude extract using chromatographic methods (Al-Bari et al., 2006; Gurung et al., 2009).
Thin layer chromatography (TLC)
The TLC plate was cut into 5 × 10 cm and the penal line was drawn on the white side up using pencil and ruler. Consequentially, 10 µl of the crude extract to be separated was spotted in a single small spot of 1.5 cm from the end of the TLC plate using capillary tube. Before inserting the TLC plate, the developing solvent was poured into the solvent tank to cover the bottom of the tank to a depth of 1.0 cm. When the spot was dried, the plate was immersed into solvent tank containing a 1:9 solvent mixture of chloroform: methanol. The solvent was allowed to get to about 90% of the way up, the plate was taken out of the jar with forcipes, and then the solvent front was marked with a pencil immediately and allowed to dry. Spot was visualized with ultraviolet (black) lamp at 254 and 366 nm (Fessenden, 1993; Gurung et al., 2009). Visualization of the spot was also done with vanillin-sulfuric acid spray reagent (Al-Bari et al., 2006; Selvameenal et al., 2009). The spot was circled with pencil. The distance of the spot moved up the plate and that of the solvent was measured in cm. The retention factor (Rf) values of the antimicrobial compound were calculated by dividing distance travelled by the spot to the distance traveled by the solvent (Fessenden, 1993).
Rf = Distance traveled by the compound / Distance traveled by the solvent
To check the antimicrobial activity of single spot from TLC plates, active bioautography was done on nutrient agar plate inoculated with 0.2 ml (3×106) overnight culture of S. aureus (clinical isolates). The spots from TLC plates were cut and placed downward on the center of seeded plate. After 1 h at 4°C in refrigerator, the plate was incubated for 24 h at 37°C. The inhibition of the test organism indicated the spot has antimicrobial activity (Pandy et al., 2004).
Column chromatography
For the single spot from TLC plate, active column chromatography was performed to purify and detect the components of the antimicrobial compound based on their polarity with solvents. The column (Sorbisil 60 mesh; column dimensions 2.5 cm inner diameter × 30 cm length) was packed very tightly with silica gel as slurry with the desired solvent (ethyl acetate). 100 mg of the active crude extract was dissolved in a minimum amount of ethyl acetate and applied directly on top of the packed column. Thereafter, 10 ml of the eluting gradient chloroform: methanol solvent was added to the top of the column. The proportion of crude extract to silica gel in the solvent mixture is 1:30. The column was developed and the flow rate of the system was adjusted per minute. The elution process was conducted by adding different solvents, namely, ethanol, petroleum ether, acetone and methanol in the middle of the process (Fessenden, 1993). Thirteen different fractions of the compound were collected (each of 5 ml) from the column chromatography.
Preparative thin layer chromatography
Preparative TLC plate was prepared by suspending 160 g of Merksilca gel GF 254 powder in 260 ml of deionized water that was vigorously shaken for 45 s in 500 ml Erlenmeyer flask, with rubber stopper. The thickened slurry was poured into 10 × 20 cm plate and spread to prepare 1.0 mm trailing edge. The plate was air dried until it turned white for 45 min and activated at 120°C for half an hour. Ten microliters of the fraction to be separated was applied in a single small spot of 1.5 cm from the end of TLC plate using capillary tube. Before inserting the TLC plate, the developing solvent was poured into the solvent tank to cover the bottom of the tank to a depth of 1.0 cm. When the spot was dried, the plate was immersed into solvent tank containing chloroform: methanol system (1:9). The spots on the chromatography were visualized in UV chamber 245 and 366 nm (Al-Bari et al., 2006). The obtained band was then scratched and dissolved with ethyl acetate and centrifuged at 3000 rpm for 15 min. Supernatant was collected in a pre-weighed vial and kept for evaporation (Selvameenal et al., 2009).
In vitro antimicrobial activity of purified compound from AAUBA13
The antimicrobial potential of 13 fractions of the crude extract from AAUBA13 was tested using disc diffusion techniques.
Identification and characterization of selected isolates
Selected actinomycetes isolates: AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA19, AAUBA30 and AAUBA31 were described by some cultural, morphological, physiological and biochemical characterization to identify the isolates at genus level based on Bergey's Manual of Systematic Bacteriology, Volume four (Williams et al., 1989a,b).
Cultural (macroscopic) characterization
Cultural characteristics of the isolates were investigated by growing the isolates on starch casein agar, starch nitrate agar, glycerol asparagine agar, glucose asparagine agar and actinomycetes isolation agar. A loop full of each isolate from 7 days old culture was taken and inoculated onto each of the medium by streak plating technique and incubated at 30°C for 7 days. The experiment was done in duplicates and colony morphology was noted with respect to color of aerial mycelium and substrate mycelium, colony diameter and diffusible pigment (Williams and Cross, 1971; Remya and Vijayakumar, 2008). The results were recorded.
Microscopic characterization
The microscopic examination was carried out by cover slip culture and Gram staining methods to study the morphology of the isolates. Cover slip culture method was done according to previous works (Williams and Cross, 1971; Tiwarty, 2009) by inserting sterile cover slip at an angle of 45° in the starch casein agar medium. A loop full of isolates was taken from 7-day old culture and inoculated, at the insertion of the cover slip on the medium and incubated at 30°C for 7 days. The cover slip was carefully removed using sterile forceps and placed upward on a glass slide. The growth on the cover slip was fixed with few drops of absolute methanol for 15 min and washed with tap water and flooded with crystal violet reagent for 1 min followed by washing and blot drying. It was then examined through microscope (Wagtech, England) under magnification of ×1000 in oil immersion. The morphology of spore chains and hyphae of substrate and aerial mycelia were observed and the picture was captured by U-LH100HG Florescent microscope (OLYMPUS BXSI, JAPAN) in the Applied Microbiology Laboratory, Faculty of Life Science, AAU. The observed characteristics were compared with the actinomycetes morphology provided in Bergey’s manual of Systematic Bacteriology (Williams et al., 1989a,b).
Physiological characterization
Physiological characters of the isolates were studied on the basis of pH tolerance, temperature tolerance, resistance towards sodium chloride and utilization of carbon and nitrogen sources.
pH tolerance
A loop full of the test isolate from 7 days old culture was taken and serially diluted from 10-1 - 10-6 in sterile distilled water; it was agitated with vortex and about 0.1 ml of the suspension was taken and inoculated with spread plate technique onto starch casein agar media which was adjusted to pH levels of 5, 6, 7, 8, 9, 10, 11, and 12. The experiment was done in duplicates and colony was counted with log colony forming unit after incubating the isolates at 30°C for 7 days. The result was recorded (Laidi et al., 2006).
Temperature tolerance
Temperature tolerance of the isolates was determined on nutrient agar plates (Oxide). A loop full of the test isolate from 7 days old culture was taken and serially diluted from 10-1 - 106 in sterile distilled water; it was agitated with vortex and 0.1 ml of the suspension was taken and inoculated with spread plate technique. The experiment was done in duplicate and colony was counted with log colony forming unit after incubating the isolates at 15, 25, 30, 37 and 45°C. The results were recorded (Laidi et al., 2006).
Growth with sodium chloride
The isolates were tested for their levels of tolerance to sodium chloride on nutrient agar (Oxide) supplemented with 5, 7 and 10% sodium chloride. Agar plates were inoculated with test isolates with streak plate technique. The experiment was done in duplicates. The plates were incubated at 30°C for 7 days and observations were made to record the highest concentration of salt that allows the growth (Santhi et al., 2010).
Utilization of carbon and nitrogen source
Different carbon and nitrogen sources were tested, in the basal medium consisting of (g/L) K2HPO4, 1.0; MgSO4.7H2O, 0.5; CaCl2.2H2O, 0.04; FeSO4.7H2O, 0.005; ZnSO4.7H2O, 0.0005; 15 g agar and 1.0% of each of the carbon and nitrogen sources such as D-mannose, sucrose, D-galactose, D-glucose, L-arabinose, D-mannitol, cellobiose and D-fructose were used. Nitrogen sources such as L-arganine, yeast extract, peptone, (NH4) SO4 and (NH4) H2PO4 and pH were adjusted to 7. The carbon and phosphate sources were sterilized separately and added just prior to inoculation. Each isolate from 7 days culture was inoculated with streak plate technique. The experiment was done in duplicates and the plates were incubated at 30°C together with basal medium as negative control. The growth was read after 7, 14, and 21 days and the results were recorded as abundant (When growth on tested Carbon or Nitrogen in basal medium greater than growth on basal medium), moderate (when growth on tested carbon or nitrogen in basal medium is significantly better than growth on basal medium), and good (when growth on tested carbon or nitrogen in basal medium is less than growth on basal medium) (Shirling and Gottlieb, 1966; Oskay et al., 2004; Pandy et al., 2005).
Biochemical characteristics
Biochemical characteristics of the isolates were studied. They include gelatin hydrolysis, starch hydrolysis, and esculin degradation.
Starch hydrolysis
Starch hydrolysis was done using starch agar plates having a composition of soluble starch, 20 g; beef extract, 3 g; peptone, 5 g; agar, 15 g, and distilled water, 1 L. The isolates were taken from 7 days old culture. They were streaked on the media and incubated at 30°C for 7 days together with uninoculated plates that serve as a control. The iodine solution was flooded onto the plates to see the clear zone of hydrolysis around the colony (Aneja, 2005; Remya and Vijayakumar, 2008).
Gelatin hydrolysis
This test was done on sterile nutrient gelatin media having a composition of beef extract, 3 g; peptone, 5 g, gelatin, 120 g, and distilled water, 1 L. Each isolate was taken from 7 days old culture and stabbed into nutrient gelatin tubes with sterile needle. The tubes were incubated for 10 days at 30°C together with uninoculated tube used as control. After incubation, the tubes were placed in refrigerator, at 4°C for 15 min (Aneja, 2005; Sundaramoorthi et al., 2011). The refrigerated gelatin tubes were examined to see whether the medium was liquid for positive test or solid for negative test.
Esculin degradation
Esculin degradation was determined after Kutzner (1976). The isolates were taken from 7 days old culture and streaked into Esculin agar slants having a composition of yeast extract, 0.3 g; ferric ammonium citrate, 0.05 g, agar, 0.75 g; 0.1% of esculin, and 50 ml distilled water; they were incubated at 30°C for 7 days. Observations were made between the periods of incubation to check the blackening of the medium. Positive tests were confirmed compared to the control which was a dark brown substrate.
Data analysis
Data analysis was made by Statistical Package for Social Sciences (SPSS) windows version 17 in terms of the mean of the growth inhibition zone value obtained from each of the seven bacterial test pathogens and two fungal test pathogens. The data on temperature and pH tolerance were analyzed by comparing the mean growth in log colony forming unit through analysis of variances (ANOVAs). The mean for each group was compared using Tukey test (HSD).
RESULTS AND DISCUSSION
Cultures of actinomycetes isolated from different habitats
A total of 30 different actinomycete isolates were recovered from rhizosphere of different plants and soil samples collected from Debrezit and Holeta farm lands and garden soils of Arat Kilo campus. Sixty seven percent of the actinomycetes were isolated from rhizosphere of plants. Rouatt et al. (1951) and Geetanjali and Jain (2016) also reported that greater percentages of actinomycetes are found in rhizosphere soils, whereas 33% of the isolates were recovered from farm soil by using starch casein agar and actinomyctes isolation agar medium supplemented with Amphotracin B (50 μg/ml) (Table 1).
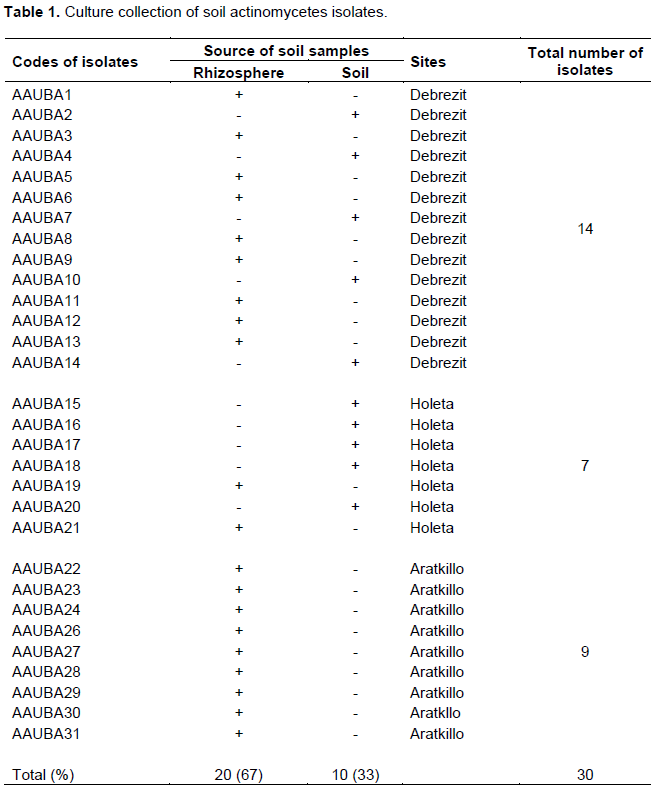
Primary screening of the antimicrobial producing isolates
Out of the 30 actinomycetes isolates subjected for primary screening, 18 (60%) isolates showed varying levels of antimicrobial activities against the five test microorganisms (Table 2). Upon primary screening, nine isolates were identified based on spectrum activity against test organisms, and the potent isolates were isolated from the rhizosphere soils. The present results in agreement with that of Abo-Shadi et al. (2010) reported that microorganisms isolated from rhizosphere soil could be an interesting source of antimicrobial bioactive substance. Ramakrishnan et al. (2009) also reported that rhizosphere soil can serve as an effective source of antimicrobial compounds. This ratio is much higher than the 22 and 34% reported by Abo-Shadi et al. (2010) and Oskey et al. (2004), respectively. This difference may be attributed to the differences in inhibiting antibiotic resistant of test organisms, the genetic differences of the antibiotic producing isolates, and their capacity to produce more than one secondary metabolite.
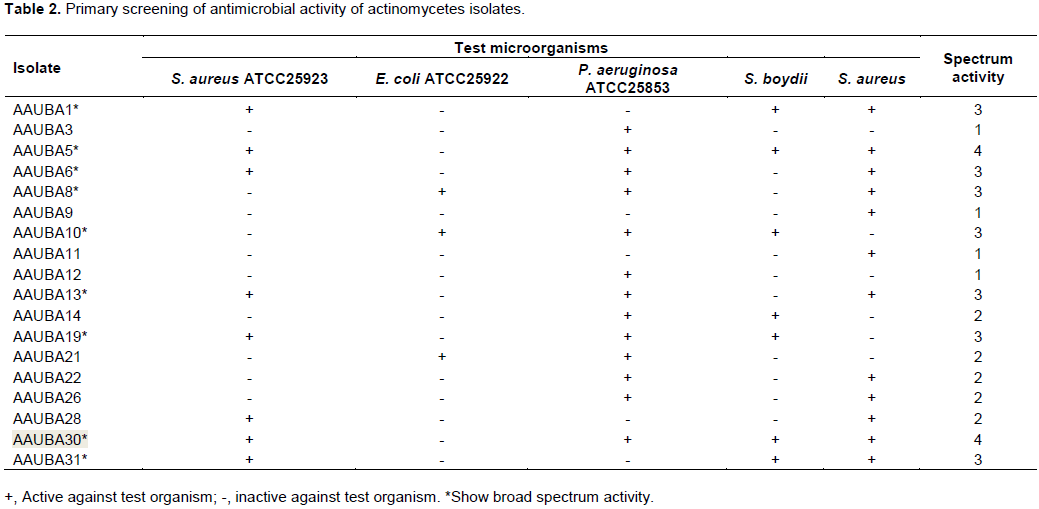
Out of the different actinomycetes isolates screened for antimicrobial activities, 2 isolates (11%) AAUBA5 and AAUBA30, showed a wide spectrum of antibiosis against 4 test organisms except E. coli ATCC 25922. Likewise, 7 (39%), 5 (28%) and 4 (22%) of isolates were found to inhibit any 3, 2, and 1 of the test organisms, respectively. Antimicrobial activities of isolates that inhibited 3 test organisms were AAUBA1, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA19 and AAUBA31. The antimicrobial activities of AAUBA14, AAUBA21, AAUBA22, AAUBA26 and AAUBA28 were found to inhibit 2 test organisms and the rest isolates inhibited 1 test organism (Table 2).
The antimicrobial activities of isolates also showed variations, in inhibiting the test organisms. Consequently, most of the actinomycetes of isolates 13 (72%) inhibited P. aeruginosa ATCC 25853, followed by antimicrobial activities of 12 actinomycetes isolates (67%) and antimicrobial activities of 8 actinomycetes isolates (44%) that inhibited S. aureus (clinical isolate) and S. aureus ATCC25923, respectively. Likewise, antimicrobial activities of 7 actinomycetes isolates (39%) showed a pattern of suppression on S. boydii (clinical isolate). The most resistant test isolate was E. coli ATCC 25922 that was inhibited by only 3 actinomycetes isolates (17%), namely, AAUBA8, AAUBA10, and AAUBA21 (Table 2).
Many isolates (39%) also showed a wide pattern of dual inhibition of P. aeruginosa ATCC 25853 and S. aureus (clinical isolate) and S. aureus ATCC25923 and S. aureus (clinical isolate). Few actinomycetes isolates (17%) had dual inhibition capacity against E. coli ATCC 25922 and most of the isolates (28%) had multiple inhibition capacity against S. aureus ATCC25923 and P. aeruginosa ATCC 25853, S. aureus ATCC25923 and S. boydii (clinical isolate). Some isolates (17%) had multiple inhibition capacity against P. aeruginosa ATCC 25853 and S. boydii (clinical isolates).
From 9 actinomycetes isolates that showed a wide spectrum of activity, antimicrobial activities of actinomycetes isolate (AAUBA10) were active against only Gram negative microorganism and antimicrobial activities of eight actinomycetes isolates (AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA13, AAUBA19, AAUBA30, and AAUBA31) were against both Gram positive and Gram negative microorganisms. Of the nine actinomycetes isolates, AAUBA5, AAUBA6, AAUBA8,AAUBA10, AAUBA13, AAUBA19, and AAUBA30 were active against P. aeruginosa ATCC 25853; AAUBA1, AAUBA5, AAUBA6, AAUBA13, AAUBA19, AAUBA30 and AAUBA31 were active against S. aureus ATCC25923; AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA30 and AAUBA31 were active against S. aureus (clinical isolate); AAUBA1, AAUBA5, AAUBA10, AAUBA19, AAUBA30 and AAUBA31 were against S. boydii (clinical isolate); AAUBA8 and AAUBA10 were active against E. coli ATCC 25922 (Table 2).
Secondary screening of selected actinomycetes crude extract by disc diffusion assay
Based on the results of primary screening, 9 active actinomycetes isolates were selected for fermentation. The active actinomycete isolates include AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA19, AAUBA30 and AAUBA31. The results of ethyl acetate crude extracts of the actinomycetes for antimicrobial activities are shown in Table 3. The crude extracts of nine actinomycetes isolates showed activities against a minimum of 1 test organism and maximum of 8.
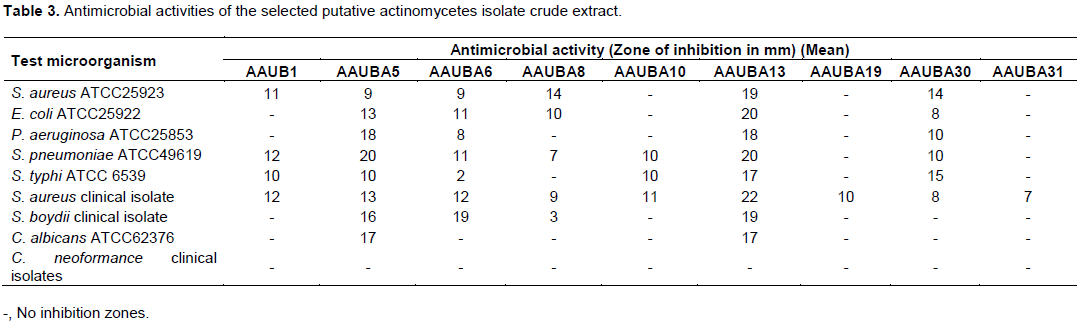
Isolate AAUBA30 crude extract was found to inhibit 6 test organisms while isolate AAUBA8 crude extract inhibited 5 test organisms. Isolate AAUBA1 crude extract was found to inhibit 4 test organisms except E. coli ATCC25922, P. aeruginosa ATCC25853, S. boydii (clinical isolate), C. albicans ATCC62376 and C. neoformance (clinical isolates). Isolate AAUBA10 crude extract inhibited S. pneumonia ATCC49619, S. typhi ATCC and S. aureus (clinical isolate). Isolates AAUBA19 and AAUBA31 crude extract inhibited only S. aureus (clinical isolate) (Table 3).
The crude extract of isolates showed variations in spectrum against test microorganisms. Consequently, AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA19, AAUBA30 and AAUBA31 crude extracts were found to inhibit Gram positive bacteria S. aureus (clinical isolate) including AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13 and AAUBA30; AAUBA1, AAUBA5, AAUBA6, AAUBA8, AAUBA13 and AAUBA30 showed inhibition spectrum against Gram positive S. pneumoniae ATCC49619 and S. aureus ATCC25923, respectively. Likewise, AAUBA1, AAUBA5, AAUBA6, AAUBA10, AAUBA13 and AAUBA30 suppressed Gram negative S. typhi ATCC 6539; AAUBA5, AAUBA6, AAUBA8, AAUBA13 and AAUBA30 and AAUBA5, AAUBA6, AAUBA13 and AAUBA30 isolates inhibited Gram negative E. coli ATCC25922 and P. aeruginosa ATCC25853; AAUBA5, AAUBA6, AAUBA8 and AAUBA13 inhibited Gram negative S. boydii (clinical isolate). Two isolates of crude extract (AAUBA5 and AAUBA13) inhibited C. albicans ATCC62376 (Table 3).
Upon the result of primary and secondary screening methods it seems that from the 9 effective isolates, more isolates were active against Gram positive bacteria than Gram negative bacteria (Tables 2 and 3). This might be due to the morphological differences on cell wall components between those two types of microorganisms. Gram negative bacteria have an outer lipopolysaccharide membrane, besides peptidoglican (Pandey et al., 2002). Hence, their cell wall is impermeable to lipophilic solutes. Porins constitute a selective barrier to the hydrophilic solutes (Nokaido and Vaara, 1985), antimicrobial compound rather than positive bacteria. In the antimicrobial bioassay screening process (Table 3), larger inhibition zone diameter was observed by crude extract of isolate AAUBA13 (22 mm) against S. aureus (clinical isolate) followed by S. pneumoniae ATCC49619 and E. coli ATCC25922 (20 mm) and the least was shown by crude extract of isolate AAUBA6 (2 mm) against S. typhi ATCC 6539. The differences in the ability to produce the clear zone were presumably dependent on the secondary metabolites that were produced by test isolates. This assumption was supported by Dharmawan et al. (2009) and Singh et al. (2016) who state there is variation in the diameter of clear zone because every isolate produces different types of secondary metabolites. Different types of secondary metabolites have different chemical structure, compounds and chemical concentration.
The resistance test isolates were S. aureus ATCC25923, E. coli ATCC25922, P. aeruginosa ATCC25853, S. pneumoniae ATCC49619, S. typhi ATCC 6539 and S. boydii (clinical isolate) that were resistant to one of the antagonistic isolates. The most sensitive one was S. aureus (clinical isolate) that was inhibited by all isolates crude extract; they vary in inhibition zone diameter. However, the most resistant test isolate was C. neoformance (clinical isolates) against all antagonistic isolates, followed by C. albicans ATCC62376 except against AAUBA5 and AAUBA13 crude extract (Table 3).
Accordingly, crude extract of isolates AAUBA5 and AAUBA13 showed a wider spectrum of antimicrobial activities. In this study, isolate AAUBA13 crude extract was found to be the best isolate from antimicrobial bioassay method; it is shown by its broad-spectrum activity with high mean zone of inhibition (22 mm) more than AAUBA5 crude extract and others. This isolate crude extract might produce more than one antimicrobial metabolites that make them effective inhibitor to Gram positive and Gram negative bacterial pathogen and also effective inhibitor to C. albicans ATCC62376 fungal pathogen (Gurung et al., 2009).
The antimicrobial activity of the crude extract of AAUBA13 isolate against the test pathogenic bacteria was positive with variations in inhibition zone diameter ranging from 17 to 22 mm. Among the Gram positives bacteria, S. aureus was found to be more sensitive followed by S. pneumonia ATCC49619 and S. aureus ATCC25923 in decreasing order. In case of Gram negative bacteria, E. coli ATCC25922 was more sensitive followed by S. boydii (clinical isolate), P. aeruginosa ATCC25853, and S. typhi ATCC in decreasing order. However, no activity was observed against the test fungus, C. neoformance (clinical isolate) (Table 3).
The antimicrobial activities of the 9 isolates through primary and secondary screening revealed different results. In the primary screening, isolates AAUBA5, AAUBA6, AAUBA8, AAUBA10, AAUBA13, AAUBA19 and AAUBA30 (Table 2) were found to inhibit P. aeruginosa ATCC 25853, but the fermented products of ABA8, AAUBA10 and AAUBA19 could not inhibit P. aeruginosa ATCC 25853 (Table 3). In the secondary screening, the fermented products of isolates AAUBA5, AAUBA6, AAUBA8, AAUBA13 and AAUBA30 were found to inhibit E. coli ATCC 25922, but in the primary screening, AAUBA5, AAUBA6, AAUBA13 and AAUBA30 crude extracts could not inhibit E. coli ATCC 25922 (Table 2). This might be due to the difference in the morphology of actinomycetes when grown in solid and liquid media as filamentous mycelia and fragmented mycelia, respectively. The present results are in agreement with those obtained by Pandey et al. (2004) who reported that some of the active actinomycetes isolates in the primary screening did not show any activity; but in the secondary screening some showed little activity and some showed improved activity. Bushell (1993) also reported that during the screening of the novel secondary metabolite, actinomycetes isolates failed in their antibiotic activity in liquid culture.
Detection and purification of the active compound from AAUBA13
Thin layer chromatography and column chromatography
The crude extract of AAUBA13 was run on a TLC plate and a single band (spot) with blue color was detected at Rf value 0.73. The column chromatography experiment showed 13 different fractions. This was already indicated by the color of the bands on the silica gel plate. Fractions were tested for antimicrobial activity. Accordingly, fractions 8 was identified as best active fraction and concentrated to be further characterized as given in Table 5.
Preparative thin layer chromatography
Fraction 8 was further purified using preparative chromatography technique; it resulted in single visible band when visualized with UV light, and upon scratching of these bands out from the glass plate a pure compound was obtained.
The antimicrobial form AAUBA13 showed Rf value of 0.73 in chloroform: methanol (1:9); 0.48 ethanol: petroleum ether (1:1) solvent systems and 0.57 Chloroform: ethyl acetate (1:1) solvent system (Table 4).
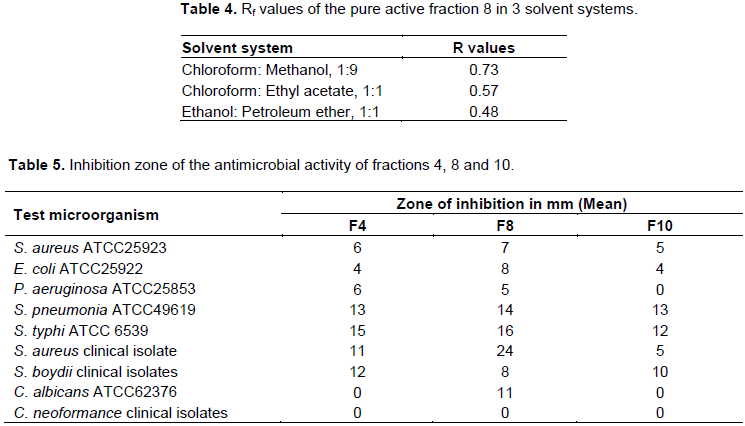
In vitro antimicrobial activities of the fractions
From the 13 fractions, only 3 were found to be active against all tested microorganisms (Table 5). Accordingly, fraction F8 displayed large inhibition zone than that of fractions F4 and F10. The highest antimicrobial inhibition activity was observed against S. aureus clinical isolate with a clear zone diameter of 24 mm; the lowest was observed against P. aeruginosa ATCC25853 with diameter of 5 mm. Fraction 4 showed activity against all test bacteria. Fraction F10 showed less activity against all test pathogens and this fraction has no activity against P. aeruginosa ATCC25853 compared to fractions F4 and F8. Only F8 has activity against C. albicans ATCC 62376. However, all fractions did not show antifungal activity against C. neoformance (clinical isolates).
In this work, the antimicrobial compound from isolate AAUBA13 crude extract separated into individual component with chromatography and test confirms the crude extract was a mixture of different compound. The active fraction eluted at fractions 4, 8 and 10. This indicates the active components have polar and non-polar nature. Fraction which moves or elutes faster (F4) and is retained least on the adsorbed silica is most likely non-polar and fraction that moves slower (F10) and is retained more on the adsorbent silica gel is polar (Atta et al., 2009).
The antimicrobial activity of the purified fraction from isolate AAUBA13 crude extract showed variations in inhibition zone diameter from 5 to 24 mm against test microorganisms. Maximum inhibition zone diameter for crude extract was 22 mm against S. aureus (clinical isolate) (Table 3), whereas the purified fraction showed inhibition zone diameter of 24 mm against S. aureus clinical isolates (Table 5). This indicates that the activity of antimicrobial compound increases as the compound becomes pure. This agrees with the work of Kavitha and Vijayalakshmi (2007). However, like that of crude extracts, purified compound did not show activity against C. neoformance. Therefore, the antimicrobial compound obtained from AAUBA13 crude extract was not effective against C. neoformance. On the other hand, the crude extract as well as purified compound obtained from AAUBA13 crude extract showed activity against C. albicans ATCC62376.
Identification and characterization of selected isolates
Morphological characterization
Among the isolates, AAUBA5 and AAUBA10 have a hook (retinaculiaperti) like structure, whereas isolates AAUBA1, AAUBA6, AAUBA8, AAUBA13, AAUBA30 and AAUBA31 showed (rectiflexible) spore chains (Table 6), with smooth spore chains of 3 or more and branched mycelium from the cover slip culture. The microscopic examination emphasized that the spore chain with rectiflexible and retinaculiaperti with smooth spore surfaces were typical characteristics of genus Streptomycetes (Williams et al., 1989a,b).
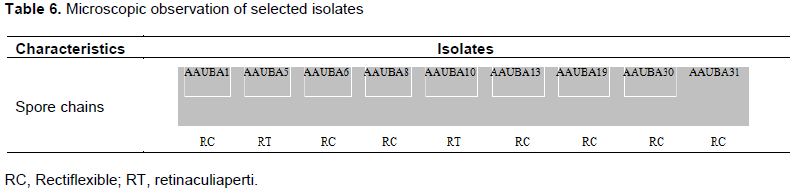
Cultural characterization
The isolates were found to grow in all media with some variations. However, abundant growth of the isolates was observed on starch casein agar medium, moderate growth was observed on actinomycetes isolation agar and good growth was observed on glycerol asparagine agar medium. Diffusible pigment was observed only on isolate AAUBA6 on starch casein agar medium and glycerol asparagine agar medium. The isolates were characterized by different colony diameter on starch casein agar medium at pH 7 (Table 7).
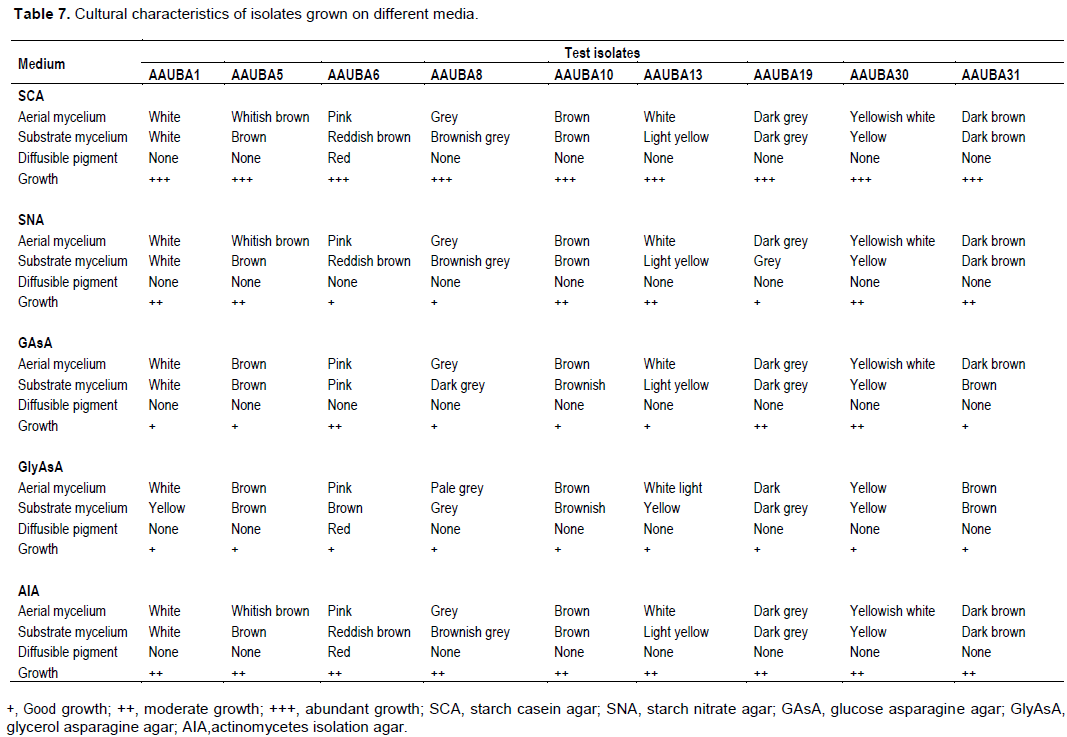
The color of the aerial and substrate mycelia varied depending on the type of the media used. The color of the substrate mycelia was also recorded (Table 7). The cultural (Macroscopic) characteristics of aerial and substrate mycelium of each isolate range from white, grey, blue, yellow, brownish, pink to dark grey. No diffusible pigment was produced except in AAUBA6 isolate, where red diffusible pigment was observed. These results reveal that the actinomycetes isolates were related to the genus Streptomyces (Cross, 1989; Lechevalier, 1989; Locci, 1989).
Physiological characteristics
pH and temperature tolerance
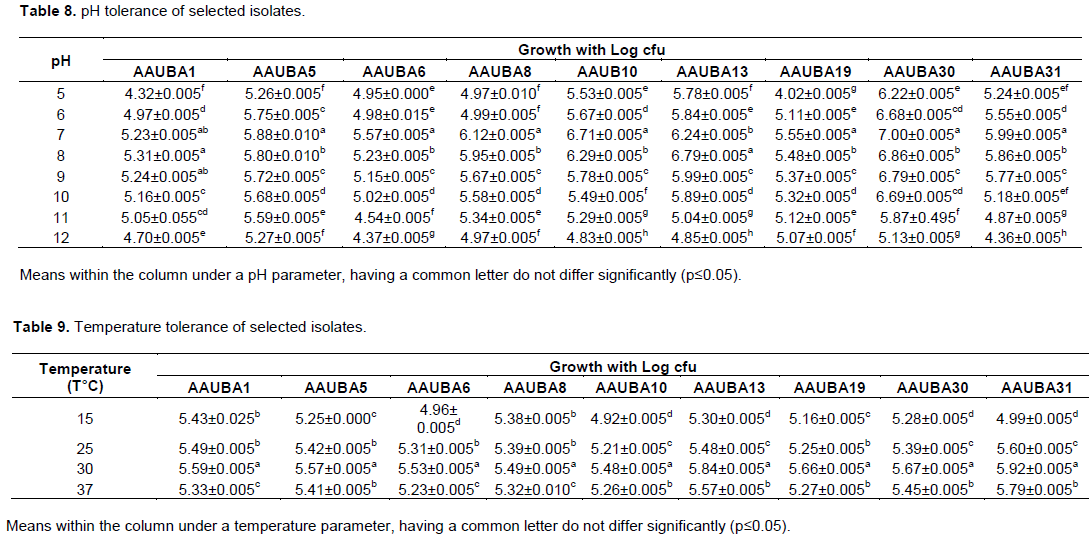
The isolates were found to grow well relatively in wider range of pH 5 to 12; maximum growth was observed at pH 7 and for isolate AAUBA1 and AAUBA13, it was pH 8. Growth tends to significantly decrease as the pH levels increase and decreases (Table 8). The isolates did grow at temperatures of 15 to 37°C, in which number of colony and colony diameter were higher than the rest of the temperature ranges. However, all isolates did not grow at temperature of 45°C (Table 9).
Growth with NaCl
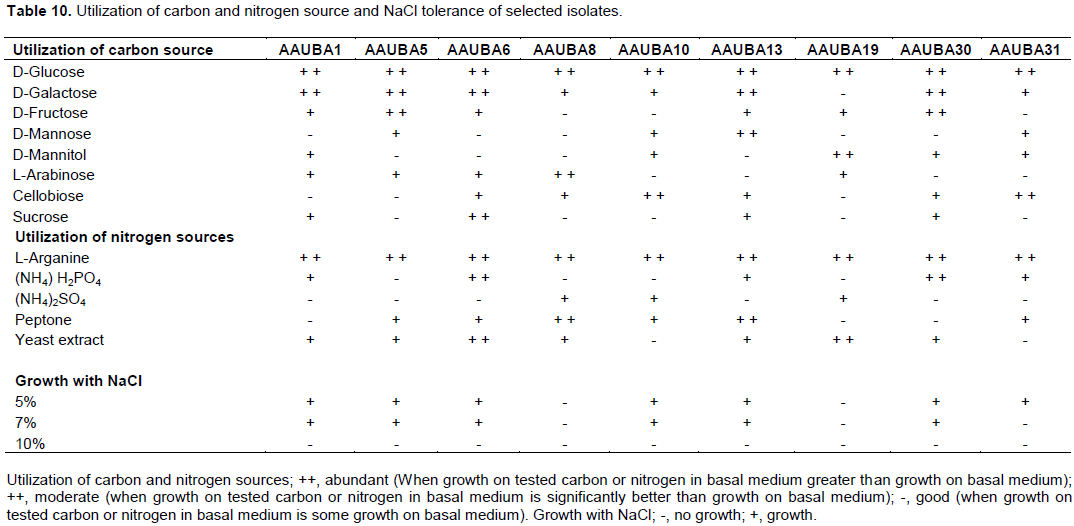
All isolates were grown at a concentration of salt supplemented with 5 and 7% except isolates AAUBA8 and AAUBA19 (Table 10). However, all isolates did not grow at a concentration of salt supplemented on 10% sodium chloride. AAUBA31 did not grow in a salt concentration supplemented with 7% of sodium chloride (Table 10).
Utilization of carbon and nitrogen sources
The isolates found to grow on a wider range of carbon and nitrogen sources were used with little variations. Table 10 shows that the isolate can utilize different carbon and nitrogen sources.
Consequently, all the isolates utilized D-glucose and L-arganine as a carbon and nitrogen source, respectively, followed by D-galactose, D-fructose, cellobiose, D-mannitol and L- arabinose, and D- mannose and sucrose as a source of carbon in decreasing order; yeast extract, peptone, (NH4) H2PO4 and (NH4)2SO4 as a source of nitrogen in decreasing order. Isolate AAUBA6 was found to utilize best both carbon and nitrogen source used, followed by isolate AAUBA13. The least utilization was shown on isolates AAUBA10 and AAUBA31. Isolates AAUBA6, AAUBA13 and AAUBA30 were best in the utilization of the carbon source used (Table 10).
Cultural characteristics of the 9 isolates varied based on the type of the media used most likely due to the nutritional versatility of the isolates. The utilization of a variety of carbon sources by this actinomycete isolates will help in adaptation to a variety of inoculation sites and wide soil types (Yadav et al., 2009). Concerning utilization of a variety of nitrogen source, the isolates produced abundant aerial mycelium on organic nitrogen source medium and good growth was observed on inorganic nitrogen source. This is also in agreement with the report of Petrova and Vlahov (2007).
Biochemical test
All of the isolates showed clear zone when flooded with iodine solution indicating that they are capable of hydrolyzing the starch; they were also found to hydrolyzed gelatin. All isolates showed a dark brown to black complex on esculin medium, which is a positive reaction to esculin degradation.
The biochemical tests, physiological properties of local isolates, as well as its carbon and nitrogen source utilization characteristics can also be compared to those of the actinomycetes described in Bergey’s Manual of Determinative Bacteriology, showing these isolates were found to be classified under genus Streptomyces (Lechevalier et al., 1989; Williams et al., 1989)
CONCLUSION
The finding of this study showed that the antimicrobial compound obtained from AAUBA13 crude extract has an antibacterial activity and also an antifungal activity. The data, in general, showed that the antimicrobial compounds obtained from AAUBA13 demonstrate broad spectrum and a remarkable antimicrobial activity against bacterial and C. albicans ATCC62376.
Actinomycetes isolates recovered from rhizosphere samples showed the potential to produce antimicrobial bioactive compounds. It is also suggested that the other isolates should be further processed to fully realize their antibiotic property on different test microorganisms. There is need for further studies to optimize the production conditions of the bioactive compounds from the potent actinomycetes isolates.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The first author would like to thank Addis Ababa University, University of Gondar, for financial support.
REFERENCES
|
Abo-Shadi MA, Sidkey NM and Al-Mutrafy AM (2010). Antimicrobial Agent Producing Microbes from some Soils' Rhizosphere in Al-Madinah Al-Munawwarah, KSA. Journal of American Science, 6:915-925. |
|
|
Adegboye MF, Babalola OO (2013). Actinomycetes: a yet in exhaustive source of bioactive secondary metabolites. Microbial Pathogens and Strategies for Combating Them, Science, Technology and Education (Mendez-Vilas A. Ed.). Formatex, pp. 786-795. |
|
|
Al-Bari AA, Mohammed AS, Sazedur RM, Ashik MM (2006). Characterization and Antimicrobial activities of a Phthalic Acid Derivative produced by Streptomycetes bangladeshiensis. A Novel Species Collected in Bangladish. Research Journal of Medicine and Medical Sciences, 1:77-81. |
|
|
Aneja KR (2005). Experiments in microbiology plant Pathology and Biotechnology. 4th ed. New Age International Publishers. New Delhi. pp. 74-128. |
|
|
Arifuzzaman M, Khatun M R, Rahman H (2010). Isolation and screening of actinomycetes from Sundarbans soil for antibacterial activity. African Journal of Biotechnology, 9:4615-4619. |
|
|
Atta HM, Dabour SM, Desoukey SG (2009). Sparsomycin Antibiotic Production by Streptomyces Spp. AZ-NIOFD1: Taxonomy, Fermentation, Purification and Biological Activities. American-Eurasian Journal of Agricultural & Environmental Science, 5:368-377. |
|
|
Awad MH, El-Shahed KY I E-Nakkadi AEM (2009). Isolation, Screening and Identification of Newly isolated soil Streptomycetes (Streptomycetes. NRC-35) for β-Lactamase inhibitor production. World Applied Sciences Journal, 7:637-646. |
|
|
Baltz RH (2007). Antimicrobials from Actinomycetes: Back to the future. Microbe, 2:125-131. |
|
|
Berdy J (2005). Bioactive microbial metabolites. The Journal of Antibiotics, 58:1-26. |
|
|
Biniam W (2008). Bioactive compounds from alkaliphilic Actinomycetes. M.Sc. Thesis, Department of Biology (Microbiology), Addis Ababa University, Addis Ababa. |
|
|
Brock TD, Madigan MT (1991). Biology of Microorganisms. 6th ed. Prentice-Hall International Inc., USA. pp. 198-223. |
|
|
Bushell ME (1993). A method for increasing the success rate of duplicating antibiotic activity in agar and liquid cultures of Streptomycetes isolates in new antibiotics screens. Journal of Fermentation and Bioengineering, 76:89-93. |
|
|
Cross T (1998). Growth and examination of actinomycetes some guidelines. In. Bergey' manual of systematic bacteriology, vol.4, Williams and Wilkins company, Baltimore. pp. 1120-1126. |
|
|
Dancer SJ (2004). How antibiotics can make us sick: the less obvious adverse effects of antimicrobial chemotherapy. The Lancet Infectious Diseases, 4:611-619. |
|
|
Demain AL, Elander R (1999). The β-lactam antibiotics: past, present, and future. Antonie Van Leeuwenhoek, 75(1-2):5-19. |
|
|
Dhanasekaran D Thajuddin N, Panneerselvam A (2009). Distribution and Ecobiology of Antagonistic Streptomycetes from Agriculture and Coastal Soil in Tamil Nadu, India. Journal of Culture Collections, 6:10-20. |
|
|
Dharmawan IWE, Retno K, Made SP (2009). Isolation of Streptomyces spp. In Bali Barat National Park and inhibition test to five diarrheagenic Escherichia coli strain. Biodiversitas Journal of Biological, 13:1-6. |
|
|
Enright MC (2003). The evolution of a resistant pathogen-the case of MRSA. Current Opinion in Pharmacology, 3:474-79. Fernando P (2006). The historical delivery of antibiotics from microbial natural products-can history repeat? Biochemical Pharmacology, 71:981-990. |
|
|
Fessenden JR (1993). Organic Laboratory Techniques. Books/Cole Publishing Company Pacific Grove, California. pp. 189-240. |
|
|
Geetanjali, Jain P (2016). Antibiotic production by rhizospheric soil microflora - a review. International Journal of Pharmaceutical Sciences and Research, 7(11):4304-4314. |
|
|
Gurung DT, Sherpa C, Agrawal VP, Lekhak B (2009). Isolation and Characterization of Antibacterial Actinomycetes from Soil Samples of Kalapatthar, Mount Everest Region. Nepal Journal of Science and Technology, 10:173-182. |
|
|
Hackl E Boltenstern S, Bodrossy L, Sessitsch A (2004). Comparison of diversities and compositions of bacterial populations inhabiting natural forest soils. Applied and Environmental Microbiology, 7:5057-5065. |
|
|
Hassan MA, El-Naggar MY, Said WY (2001). Physiological factors affecting the production of an antimicrobial substance by Streptomyces violatus in batch cultures. Egyptian Journal of Biology, 3:1-10. |
|
|
Kavitha and Vijayalakshmi M (2007). Studies on Cultural, Physiological and Antimicrobial activities of Streptomycetesrochei. Journal of Applied Sciences Research, 3(12):2026-2029. |
|
|
Kornwendisch F, Kutzner HJ (1992). The family Streptomycetaceae. In. The Prokaryotes. (Balows A, Truper HG, Dworkin M, Harder W Schleifer KH Eds). Springer-Verlag, New York. pp. 921-995. |
|
|
Kumbhar C, Watve M (2013). Why antibiotics: A comparative evaluation of different hypotheses for the natural role of antibiotics and an evolutionary synthesis. Natural Science, 5:26-40. |
|
|
Laidi RF, Kansoh AL, Elshafi AM, Cheikh B (2006). Taxonomy, identification and biological and activity of a novel isolate of streptomycestendae. Arab Journal of Biotechnology, 9:427-436. |
|
|
Lalitha MK (2004). Manual on Antimicrobial Susceptibility Testing. Department of Microbiology, Christian medical college, Tamil Nadu. pp. 1-47. |
|
|
Lechevalier HA (1989). A practical guide to genetic identification of actinomycetes. In. Bergey's manual of Systematic Bacteriology. (Williams, S.T., Sharp, M.E. and Holt, J.P. Eds). The William and Wilkins company, Baltimore. pp. 2344-2347. |
|
|
Livermore DM (2003). Bacterial resistance: origins, epidemiology and impact. Clinical Infectious Diseases, 36:11-23. |
|
|
Locci R (1989). Streptomycetes and related genera. In. Bergey's manual of systematic bacteriology. (Williams, S.T., sharp, M.E. and Holt, J.P. Eds). The William and Wilkins company, Baltimore. pp. 2344-2347 |
|
|
Miyadoh S (1993). Research on antibiotic screening in Japan over the last decade: a producing microorganism approach. Actinomycetologica, 7:100-106. |
|
|
Moges K (2009). Antimicrobial compound from thermophlicactinomycete isolated from mushroom compost. M.Sc. Thesis, Department of Biology (Microbiology), Addis Ababa University, Addis Ababa. |
|
|
Ningthoujam DS, Sanasam S, Nimaichand S (2009). Screening of Actinomycete Isolates from Niche Habitats in Manipur for Antibiotic Activity. American Journal of Biochemistry and Biotechnology, 5:221-222. |
|
|
Nokaido H, Vaara M (1985). Molecular basis of bacterial outer membrane permeability. Microbiological Reviews, 49:1-32. |
|
|
Okeke IN, klugman KP, Bhuta, ZA, Duse AG, Jenkins P, Brien TF, Pablos- mendez A, Laxminarayan R (2005). Antimicrobial resistance in developing countries. Part II: strategies for containment. The Lancet Infectious Diseases, 5:568-580. |
|
|
Oskay M, Tamer A, Azeri C (2004). Antibacterial activity of some actinomycetes isolated from farming soils of Turkey. African Journal of Biotechnology 3:441-446. |
|
|
Pandey B, Ghimirel P, Prasad V, Thomas M, Chan Y and Ozanick S (2002). Studies of the antimicrobial activity of the actinomycetes isolated from the Khumby region of Nepal. Department of Bacteriology, University of Wisconsin Madison, Wisconsin. |
|
|
Pandey B, Grimier P, Agrawal VP (2004). Studies on the antibacterial activity of actinomycetes isolated from the Khumbu region of Nepal. Journal Biology Science, 23:44-53. |
|
|
Parungao MM, Maceda EBG, Villano MAF (2007). Screening of Antibiotics Producing Actinomycetes from Marine, Brackish and Terrestrial Sediment of Samal Island, Philippines. Journal of Research in Science, Computing and Engineering 4:29-38. |
|
|
Petrova D, Vlahov, S (2007). Taxonomic characterization of the thermophlicactinomycete strain 21E-produce of termostabel collagenase. Journal of Culture Collections, 5:3-9. |
|
|
Ramakrishnan J, Shunmugasundaram M, Narayanan M, (2009). Streptomyces sp. SCBT isolated from rhizosphere soil of medicinal plants is antagonistic to pathogenic bacteria. Iranian Journal of Biotechnology, 7:75-81. |
|
|
Remya M, Vijayakumar R (2008). Isolation and Characterization of marine antagonistic actinomycetes from west coast of India. Medicine and Biology,15:13-19. |
|
|
Rondon M, August P, Bettermann AD, Brady SF, Grossman TH, Liles MR, Loiacono, KA, Lynch, BA, Macneil C, Minor C, Tiong M, Osborne J, Clardy J, Handelsman J Goodman R (2000). Cloning the soil metagenome: a strategy for accessing the genetic and functional diversity of uncultured microorganisms. Applied and Environmental Microbiology 66:2541-2547. |
|
|
Rouatt JW, Lechevalier M, Waksman SA (1951). Distribution of antagonistic properties among actinomycetes isolated from different soils. Antibiotics & Chemotherapy (Northfield, Ill.), 1:185-92. |
|
|
Saadoun I, Gharaibeh R (2003). The Stereptomyces flora of Badia region of Jordan and its potential as a source of antibiotics active against antibiotic resistant bacteria. Journal of Arid Environments, 53:365-371. |
|
|
Sanglier JJ, Haag TA, Huck T A (1993). Novel bioactive compounds from actinomycetes. A short review (1988-1992). Research in Microbiology, 144:633-642. |
|
|
Santhi VS, Jose PA, Solomon RDJ (2010). Isolation and characterization of antagonistic actinomycetes from mangrove sediments. International Journal of Current Research, 3:020-023. |
|
|
Selvameenal L, Radhakishnan M, Balagurunathan R (2009). Antibiotic Pigment from Desert Soil Actinomycetes; Biological Activity, Purification and Chemical Screening. Indian Journal of Pharmaceutical Sciences, 71:499-504. |
|
|
Shirling EB, Gottlieb D (1966). Methods for characterization of Streptomycetes species. International Journal of Systematic and Evolutionary Microbiology, 16:313-340. |
|
|
Singh V, Haque S, Singh H, Verma J, Vibha K, Singh R, Jawed A, Tripathi CKM (2016). Isolation, screening and identification of novel isolates of Actinomycetes from India for Antimicrobial Application. Frontiers in Microbiology, 7:1921-1928. |
|
|
Sundaramoorthi C, Vengadesh PK Gupta S, Karthic K, Tamilsehli V (2011). Production and characterization of antibiotics from soil–isolated actinomycetes. International Research Journal of Pharmacy, 2:114-118 |
|
|
Tiwarty KB (2009). Protocol for actinomycetes studies in RLABB.2008/9. Central department of microbiology, institution of science and technology. Tribhuven University, kiripur. p. 21. |
|
|
Tortora GJ, Funke BR, and Case CL (2010). Microbiology: An Introduction, 10th ed. Pearson Benjamin Cummings, USA. pp. 530-589. |
|
|
Vijayakumar R, Muthukumar C, Thajuddin N, Panneerselvam A, Saravanamuthum R (2007). Studies on the diversity of actinomycetes in the Palk Strait region of Bay of Bengal, Inidia. Actinomycetologica, 21:59-65. |
|
|
Williams ST, Cross T (1971). Actinomycetes. In. Methods in Microbiology. (Booth, C. Ed.). Academic press, London. pp. 29-65. |
|
|
Williams ST, Goodfellow M, Alderson G (1989a). Genus Streptomycetes. In. Bergey's Manual of Systematic Bacteriology. (Williams, S.T., Sharpe, M.E. and Holt, J.P. Eds). The Williams and Wilkins Company, Baltimore. pp. 2452-2490. |
|
|
Williams ST, Sharpe ME, Holt JG (1989b). Bergey's Manual of Systematic Bacteriology. Vol. 4, William and Wilkins Co., Baltimore. |
|
|
Wohlleben W, Mast Y, Stegmann E, Ziemert N (2016). Antibiotic drug discovery. Microbial Biotechnology, 9(5):541-548. |
|
|
Yadav AK, Kumar R, Saikia, R, Bora TC, Arora DK (2009). Novel copper resistant and antimicrobial Streptomyces isolated from Bay of Bengal, India. Journal of Medical Mycology, 19:234-240. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


