Full Length Research Paper

ABSTRACT
Aluminium (Al) toxicity is an important stress factor for plants in acidic environments. Currently, there exists a dearth of research information on sustainable approaches that can mitigate the problem of Al toxicity in sub Saharan Africa. It is often assumed that since plants have the ability to synthesize their native antioxidants, they may not need supplementation to cope with stresses. To challenge this assumption, the present study explored the effects of 50 and 200 µM concentrations of lycopene (ly) on the growth and productivity of Amaranthus hybridus subjected to 3 × 10-2 mM concentration of Al at pH 4.6 and 25°C. High performance liquid chromatography (HPLC) was used to determine the peak absorbance at 375 nm and the thiobarbithuric acid (TBA) method was used to determine antioxidant activity of ly. Pre- and post-antioxidant treatments were hydroponically applied to A. hybridus seedlings before and after Al treatment for 72 h. Results show that post-lycopene treatments significantly alleviated Al stress in A. hybridus more than pre-lycopene treatments. It is concluded that though A. hybridus could synthesize its own antioxidants, it is susceptible to Al toxicity-induced stress and post-lycopene supplementation could alleviate the stress situation and enhance growth and productivity. However, A. hybridus plants appear not to need lycopene while not under Al stress.
Key words: Amaranthus hybridus, growth and productivity, hydroponics, aluminium toxicity, stress amelioration, lycopene.
INTRODUCTION
Soil acidity limits agricultural production globally especially in the Sub-Saharan African region where food security is a global concern. Different kinds of stresses, mainly Al stress, generated from acid soils affect plant growth and result in food shortage and production (Carvalho et al., 1980; Roy et al., 1988; Foy, 1992). Trivalent Al (Al3+) is one of the most toxic forms of soluble Al and is known to damage root cells at sites in the apoplast and cytosol that rapidly inhibit root growth (Kochian, 1995; Hede et al., 2001). Al is the third most abundant metallic element and is readily available to plants when the soil pH drops below 5.5. Some agricultural practices, such as leaching of nitrogen below the plant root zone, inappropriate use of nitrogenous fertilizers and increased excessive accumulation of organic matter, are major sources of soil acidification (Mohan et al., 2008). Fatur et al. (2002) explained that Al toxicity could be transferred to animals and man by consuming plants that accumulated Al. Amaranth species are erect or spreading fast growing annuals, with heights varying between 0. 3 and 2 m, ideal for plant studies in the tropic. Reports indicate that plants grown in pH 6.4 soil were significantly taller with more branches, leaves and greater leaf area than plants grown in pH 5.3 or 4.7 soil. Soil acidity impeded both top and root growth of the plant (Singh and Whitehead, 1992; Palada and Chang, 2003).
The study of Al toxicity has elicited divergent views, if not controversies, over the years and this may continue for a long time. It has been generally observed that plants grown in acid soils due to Al solubility, at low pH, have reduced root systems and exhibit a variety of nutrient-deficiency symptoms, with a consequent decrease in yield. In many countries with naturally acid soils, which constitute about 40% of world arable soil (Le Noble et al., 1996), Al toxicity is a major agricultural problem, and is intensively studied in plant systems. The effects of aluminium on plant growth, crop yield, up take and nutrients distribution in vegetative and reproductive parts are still not fully understood (Mossor-Pietraszewska, 2001). In their own contribution, Aniol and Gustafson (1984) observed that genetic variation in the response to Al toxicity has been found not only among plant species but also among cultivars within species. Other views on the complex nature of Al toxicity studies have been expressed (Kinraide, 1991; Lazof and Holland, 1999; Zatta et al., 2002; Kidd and Proctor, 2001; Exley, 2004). The role of Al in rhizotoxicity is a complex one and in spite of the enormous amount of information gathered from several studies, a lot of things are yet to be known about the complex process. A renowned authority in rhizotoxicity, Kinraide (1991), remarked that Al hydrolysis creates considerable uncertainty in the study of Al rhizotoxicity. A strict attribution of toxicity to Al3+ is not possible because Al3+ is always in equilibrium with its hydrolysis products.
Osaki et al. (1997) classified some tropical plants based on their Al tolerance. Plants in which growth was reduced by low and high Al applications were designated as Al-sensitive plant and Al-medium tolerant plants, respectively, while plants in which growth was not affected or was stimulated by Al application were designated as Al-tolerant plant and Al-stimulated plants, respectively. Plants tolerant to or stimulated by Al were further classified based on the criteria of Al accumulation: (1) Al-excluders (2) Al root-accumulators and (3) Al-accumulators. Al toxicity is always associated with H+ which is often ignored by researchers. Some workers have observed that the solubility of Al is appreciable only at pH values below 5.5. Thus, toxicity to plants occurs only at these low pH values, with the possible exception of the toxicity of Al(OH)34- at higher pH values (Kinraide, 1990). Despite this, and in contrast to the large amount of literature on Al toxicity, very little attention has been given to H+ toxicity, even though the latter is well known to be directly detrimental to root growth (Kidd and Proctor, 2001; Koyama et al., 2000). As in Al toxicity, H+ toxicity is most severe in solutions of low ionic strength and low cation concentrations, and increasing the concentration of Ca2+ and other cations in the external solution reduces or even abolishes the detrimental effects of acidity (Marschner, 1991). An evaluation of these low-pH effects is necessary for greater understanding and correct interpretation in studies of Al toxicity, but this is rarely undertaken (Lazof and Holland, 1999; Samac and Tesfaye, 2003).
Ryan et al. (1993) found the root apex to be the most Al-sensitive zone and Sivaguru and Horst (1998) identified the distal transition zone (DTZ) as the specific site of Al sensitivity. The mechanisms of Al-induced inhibition of root growth, however, are still not clearly established (Kochian, 1995; Barcelo et al., 1996; Matsumoto, 2000; Barcelo and Poschenrieder, 2002; Kochian et al., 2004). Pioneer work by Clarkson (1969) revealed Al induced alterations of root development, indicating that cell division is a primary site for Al-induced root growth inhibition. Further studies reporting Al-induced inhibition of cell division in root tips and the observation that Al binds to nucleic acids supported the view of Al-induced inhibition of root cell proliferation as a primary target for Al-toxicity (Matsumoto et al., 1976; Yamamoto et al., 2001). Lycopene (Ly) and other carotenoids partake in the crucial process of photosynthesis by absorbing energy across a wider spectrum of light and then transferring the absorbed energy to chlorophyll in order to drive the events of photosynthesis (Polivka and Frank, 2010).
Ly is synthesized in plants that produce the yellow and red colours in leaves and fruits most frequently during ripening. Cazzonelli (2011) described that ly and other carotenoids play crucial roles in protecting chloroplasts of plants as well by regulating the amount of light absorbed by plants. In essence, excess light could destroy the photosynthetic apparatus without the presence of carotenoids. Additionally, plants regulate the amount of light being absorbed by absorbing reactive oxygen species (ROS) derived from triplet chlorophyll. Such behaviour is mediated by the action of ly and other carotenoids contained in plants (Cazzonelli, 2011). At the molecular level, Al stress is known to cause drastic changes in the expression patterns of genes, some of which are quite important in the mediation of responses to oxidative stress (Maron et al., 2008). It is thus inherent that exposure of plants to Al elicits the production of ROS, which may damage cellular components if antioxidant defences are jeopardized (Darko et al., 2004; Sharma and Dubey, 2007). Other pertinent functions of ly include the attractive colours it imparts to leaves, fruits and flowers, thus facilitating plant pollination and seed dispersal (Walter et al., 2010). Ly is one of the most potent natural antioxidants (Miller et al., 1996) and has been suggested to prevent carcinogenesis and athero-genesis by protecting critical biomolecules including lipids, low-density lipoproteins (LDLs), proteins and DNA (Hodis et al., 1995; Agarwal and Rao, 1998; Rao and Agarwal, 1998). Several studies have indicated that ly is an effective antioxidant and free radical scavenger. Ly, because of its high number of conjugated double bonds, exhibits higher singlet oxygen quenching ability when compared to β-carotene or α-tocopherol. In in vitro systems, ly was found to inactivate hydrogen peroxide and nitrogen dioxide (Rao and Agarwal, 2000). Using pulse radiolysis techniques, Rao and Agarwal (2000) demonstrated its ability to scavenge nitrogen dioxide (NO2·), thiyl (RS·) and sulphonyl (RSO2·) radicals. Ly is highly lipophilic and is most commonly located within cell membranes and other lipid components. It is, therefore, expected that in the lipophilic environment ly will have maximum ROS scavenging effects. Oxidative modification of LDLs is hypothesized to be the key step in the atherogenic process, and LDL-associated anti-oxidants provide protection against this oxidation (Rao and Agarwal, 2000). In vitro ly and other carotenoids are able to inhibit oxidation of LDLs (Agarwal and Rao, 1998).
According to Gong et al. (2005), environmental stresses increase the formation of ROS that oxidize membrane lipids, protein and nucleic acids. Plants with high levels of antioxidants, either constitutive or induced, have greater resistance to oxidative damage (Sudhakar et al., 2001). Aftab et al. (2010) noted that all the treatments in which Al was applied to experimental plants had raised the activities of antioxidant enzymes. With increasing amounts of Al in the soil medium, a concomitant increase in the activities of catalase (CAT), peroxidase (POX) and superoxidase dismutase (SOD) antioxidant enzymes were observed in the exposed plants (Boscolo et al., 2003; Siminovicova et al., 2004; Sharma and Dubey, 2007; Zhen et al., 2007; Aftab et al., 2010). The fact that despite the increase in the activities of elevated levels of the antioxidant enzymes under Al toxicity, as many workers have reported; Al sensitive plants are still damaged, even under low Al con-centrations, suggests that both the constitutive and induced high levels of antioxidants may be insufficient to combat the stress situation. Such a situation will undoubtedly lead to crop failure and aggravation of food security problems in the sub Saharan region. It is against this backdrop, and to contribute to the study of the complex problem of Al toxicity, that the present study was undertaken, with the focus of exploring the possible effects of supplemented antioxidant (ly) on both the vegetative and reproductive growth of A. hybridus, subjected to mild Al induced stress.
MATERIALS AND METHODS
Soil analysis
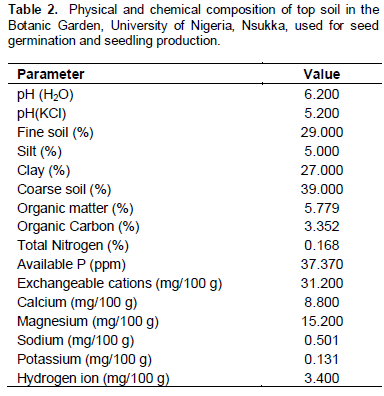
The top soil for raising the nursery was collected from the Botanic garden, University of Nigeria, Nsukka. A sample of the soil was air dried and taken to the Department of Soil Science Laboratory, University of Nigeria, Nsukka for analysis. The analysis was done using the standard method of the Association of Official Analytical Chemists (2005).
Lycopene extraction and purification
Fresh, matured and ripe, De Rica cultivar, tomatoes weighing 10 kg were purchased from Nsukka local market. The tomatoes were washed with tap water and then ground with a national electric blender without addition of water. This was allowed to run until a homogenous paste was obtained. Water was separated from the homogenous paste using a large separating funnel (20 cm) with filter paper wrapped within. The residue obtained was air dried at room temperature. The filtrate was discarded after filtration. The red paste obtained after filtration was extracted with 1.25 L of methanol and 2.5 L of petroleum ether and vortexed to aid the extraction of ly. Pressure was not allowed to build up in the container through the release of excess gas produced from the mixture. After extraction, petroleum ether was evaporated to dryness. The tomato paste extract was purified by dispersing into 2-propanol (1: 5, wt/wt) at 60°C for 1 h. Then, 37 wt. % KOH solution (1: 4, v/ v) was added and the mixture was stirred at 50°C for 2 h. The mixture was finally washed with distilled water and filtered to obtain ly crystals following the methods of Yaping et al. (2002). The procedure was repeated to re-extract more ly from the mixture. Extracted ly was stored at 2°C and protected from light to prevent transformation to inactive isomers.
Determination of antioxidant activity
The antioxidant activity of ly was determined by obtaining its thiobarbithuric acid (TBA) value. One hundred milligrams (100 mg) of ly were thoroughly mixed with 5 mL aliquot of ethanol and poured into a 5 mL vial. A control was set up by pouring 5 mL aliquot of ethanol into another vial. From each vial, 1 mL of sample solution was measured out and mixed with 2 mL of 20% trichloroacetic acid (TCA) solution and 2 mL TBA solution. The mixture was placed in a boiling water bath for 10 min. After cooling, it was centrifuged at 3,000 rpm for 10 min and absorbance of the supernatant was measured at 532 nm. The readings were taken thrice. The antioxidant activities were calculated as below:
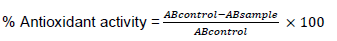
Where, ABsample = Absorbance of sample and ABcontrol = Absorbance of control (Hanachi and Golkho, 2009).
High performance liquid chromatography analysis (HPLC)
The percentage (%) purity of the extracted ly was determined using HPLC in the Department of Pure and Industrial Chemistry, UNN. UV spectrometry of each antioxidant was determined using UV-visible spectrophotometer. Five millilitre aliquot of ly dissolved in ethanol was poured into a 1 cm vial and its absorbance was
measured in the spectrophotometer using ethanol as a blank. Triple readings were taken for each sample. Spectra of ly standard in ethanol was plotted to confirm the peak absorbance of the extracted ly.
Stock preparations
Fresh 1 M stock solutions of lycopene and AlCl3 as well as full Hoagland’s nutrient solution were prepared daily, as shown below. They were stored at 4°C in a refrigerator before use.
Lycopene stock solution
One gram of ly was properly mixed with 10 mL of ethanol before the addition of 990 mL of distilled water. A 1% alcohol dilution of ly was used in this study following the protocol of Fiskesjo (1981); who showed that 1% alcohol dilutions of lipophilic solutes were not toxic to Allium roots.
AlCl3 stock solution (1 M)
This was prepared by dissolving 133.5 g of AlCl3 in little distilled water and the volume made up to 1000 mL with same. The pH was kept at 4.6.
Al treatment concentration
Al treatment concentration of 3 x 10-2mM was achieved through serial dilution and pH 4.6 was through adjustments with H2SO4.
Hoagland’s nutrient solution
This was prepared using the formulation of Hoagland and Arnon (1950 revised).
Determination of actual Al concentration in solution
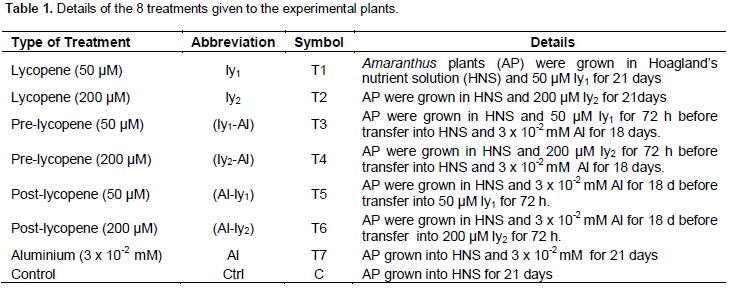
Treatments
Details of the eight treatments are presented in Table 1.
Growing Amaranthus hybridus
Viable A. hybridus seeds from the Amaranthus germplasm maintained in the Botanic garden, University of Nigeria, Nsukka (which were originally obtained from National Institute for Horticultural Research, Ibadan Nigeria) were planted in baskets containing top soil obtained from the Botanic garden, University of Nigeria, Nsukka. Both the physical and chemical constituents of the top soil were analysed in the Soil Science Analytical Lab., University of Nigeria, Nsukka; the results are presented in Table 2. The baskets were placed in the screen house in the Botanic garden. Seeds were watered with full strength Hoagland’s nutrient solution. After 54 days of planting, 120 healthy seedlings each were transplanted into separate one hundred and twenty, 50 Cl transparent plastic bottles, each containing full strength Hoagland’s nutrient solution and laid in a randomized complete block design (RCBD) in the screen house of the Botanic garden. Five plants per treatment, replicated three times, were used to monitor the growth of plants. Each A. hybridus plant was held upright in each bottle using sterilized cotton wool in such a way that shoots propped out of the container while the roots were immersed into the nutrient solution. The 120 bottles were wrapped with opaque black water-proof cellophane bags to prevent the roots from receiving light. Ten days after stabilization in the nutrient solution, the 8 treatments were applied, with Hoagland’s nutrient solution as the source of nutrient as detailed in Table 1. In total, all individual plants received treatments for 21 days. All treatments were renewed daily since the hydroponic method used was non continuous flow, to ensure uniformity of treatment and adequate supply of nutrients. At the end of this period, the experiment was terminated. Fresh and dry weights of shoots, roots and inflorescences were recorded. Other growth parameters such as number of leaves, plant height, root length, number of inflorescence, and length of inflorescence, were evaluated. The temperature of the screen house was monitored using a thermometer to ensure that the plants grew under normal temperatures.
Statistical analysis
Data collected from growth studies were analysed with one-way analysis of variance (ANOVA). Least significant difference (LSD) was used to separate means at P≤0.05 level of significance. SPSS v16, Microsoft excel 2010 and Gen-Stat packages were used for computation, data analysis and graphics.
RESULTS
Soil analysis
The result of the analysis of the soil from the Botanic garden used in the raising the A. hybridus nursery showed no presence of Al (Table 2).
Antioxidant activity
Antioxidant activity of the extracted ly was found to be 71.9%. This was significantly different from that of the ethanol blank, which exhibited a lower percentage activity of 18.5%. According to Rhee (1978), the TBA test is a colorimetric technique that measures the absorbance of a red chromogen formed between TBA and malondialdehyde (MDA). Peak absorbance of the extracted ly at 375 nm was 1.266.
Hydrogen ion concentration
Adopting the formula of Stephenson (2010); the H+ concentration of 3 x 10-2 mM of Al used for the studies (pH 4.6) was calculated to be 2.5 x 10-5 mol L-1 while that of extracted ly (pH 5.8) was calculated to be 1.5 x 10-6 mol L-1.
Determination of actual Al concentration in solution
The actual Al concentration in solution based on the 3 × 10-2 mM Al used for the study was found to be 1.85 mg/L, using the Aluminon protocol.
Vegetative and reproductive data
Average growth of A. hybridus plants was stunted after subjection to Al stress. Visible symptoms such as yellowing, wilting and loss of leaves, as well as inhibition of root growth were characteristic of the experimental plants (Figures 13 and 15).
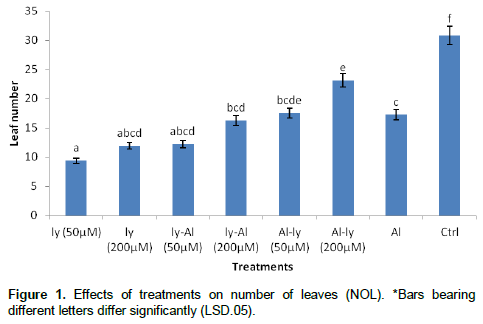
Number of leaves (NOL)
All the treatment significantly reduced the NOL when compared with the control. Al treatment reduced the NOL by 44% in comparison with the control. There was a significant difference between the NOL produced by the Al treatment (T7) and that of ly (50 μM) (T1) and Al-ly (200 μM) (T6). There was no significant difference between T7 and all the other treatments (Figure 1).
Plant height (PLH)
Similar to the situation in Figure 1, all the treatments in Figure 2 significantly reduced PLH in comparison with the control(C) (Figure 12). For T7, reduction of PLH was 36.5% when compared with C. Reduction of PLH by T7 differed significantly with that of T1 and C. The response of T6 did not differ significantly with that of Al-ly (50 μM) (T5) and T7 but differed with all the other treatments.
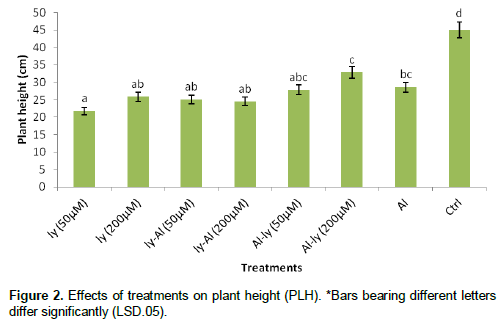
Length of inflorescence (LOI)
Figure 3 show that T7 decreased LOI by 37% in comparison with C. The response of T7 differed significantly with that of T1, ly (200 μM) (T2) and ly-Al (50 μM) (T3), but failed to differ significantly with those of ly-Al (200 μM) (T4), T5 and T6. Also, the response of T4, T5 and T6 did not differ significantly.
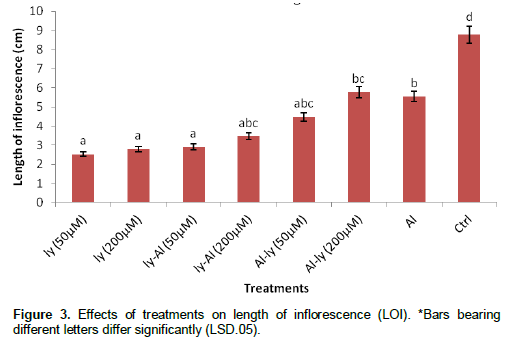
Number of inflorescence (NOI)
All the treatments in Figure 4 differed significantly with C by reduction in the NOI produced. T7 reduced the NOI by 33.5% in comparison with C. Response of the plants to T7 treatment differed significantly with that of T1, T2 and T4. It did not differ significantly with that of T3, T5 and T6. Also, the response of T6 differed significantly with that of T1, T2, T3 and T4.
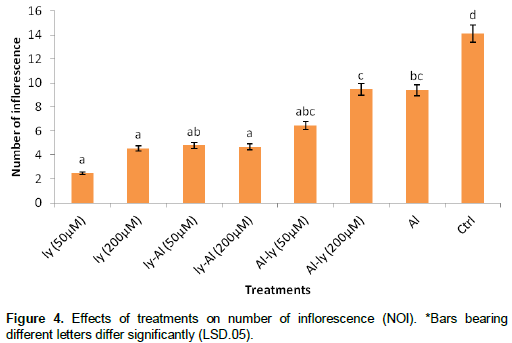
Fresh weight of inflorescence (FWI)
As shown in Figure 5, all the treatments significantly reduced the FWI. For T7, the level of reduction in comparison with the control was a staggering 69.9%. The response of T7 differed significantly with that of T1, T2 and T3, but it did not differ significantly with that of T4, T5 and T6. Also while T6 differed significantly with T1, T2 and T3, it did not differ significantly with T4, T5 and T7.
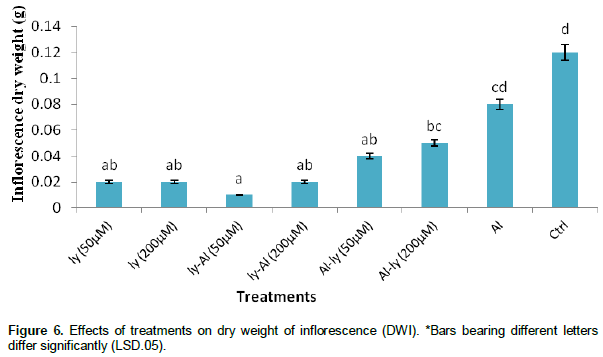
Dry weight of inflorescence (DWI)
The response of all the treatments in Figure 6 differed significantly with that of C except for T7. For T7, the response of the plants, in comparison with the control, indicate a 31.3% decrease in DWI. The response of T7 did not differ significantly with that of T6 and C, but it differed significantly with that of the rest of the treatments. While response of plants to T6 differed significantly with responses to T3 and C, it did not differ significantly with that of T1, T2, T4, T5 and T7.
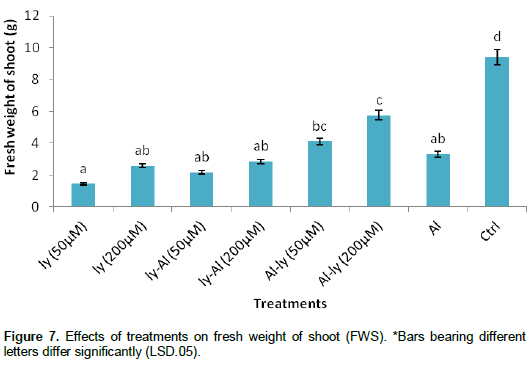
Fresh weight of shoot (FWS)
Figure 7 shows are similar to the other parameters, all the treatments significantly reduced the FWS in comparison with C. The level of reduction, for T7, was as high as 65%. This response differed significantly with responses from T6 and C but did not differ significantly with the rest of the treatments. T6 did not differ significantly with T5, but differed significantly with the rest of the treatments. While response to T5 differed significantly with that of T1 and C, it did not differ significantly with the rest of the treatments.
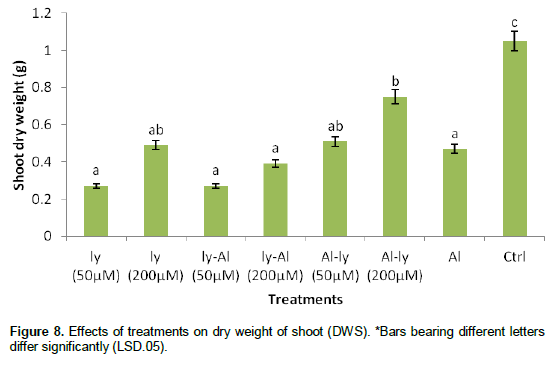
Dry weight of shoot (DWS)
Similarly, all the treatments significantly reduced the DWS when compared with C (Figure 8). A comparison between T7 and C showed a 54.9% level of reduction in DWS. T7 differed significantly from T6 but did not differ significantly with the rest of the treatments. While the response of T6 differed significantly from that of T1, T3, T4, and T7, it did not differ significantly with that of T2 and T5.
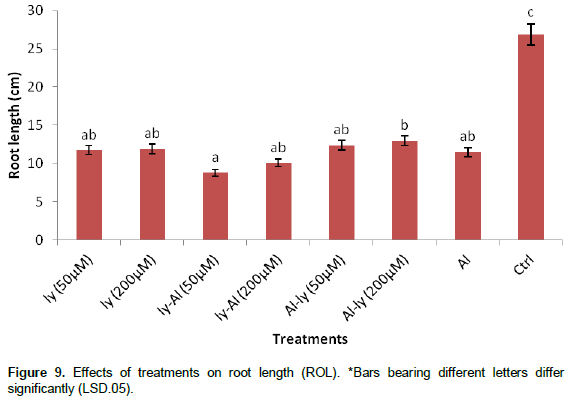
Root length (ROL)
The response of the plants to the treatments with respect to ROL (Figure 9), indicate that C (Figure 14) differed significantly with the rest of the treatments. T7 reduced root length by 57% in comparison with C. This response did not differ significantly with that of all the other treatments. T6 differed significantly with only T3.
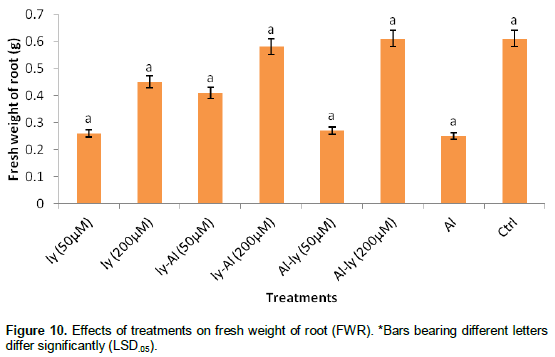
Fresh weight of root (FWR)
The response of all the treatments measured in terms of FWR (Figure 10) turned out to be insignificant in comparison with each other (including C). Though T7 in comparison with C reduced FWR by 59.9%, but this turned out to be insignificant. Same is true for T1 and T5.
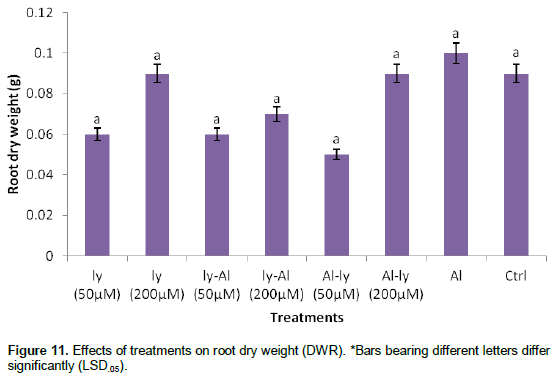
Dry weight of root (DWR)
Similar to the situation with FWR (Figure 10), the response of plants to all the treatments with regards to DWR (Figure 11) turned out to be completely insignificant. Incidentally, the response of the plants to T7 treatment was higher than that of all the treatments (including C) though this did not differ significantly with the other treatments. This shows that T7 treatment resulted to increase in DWR more than any other treatment. This was 13.5% higher than that of C. Also, T2 and T6 resulted to 1% increase in DWR in comparison with C.




DISCUSSION
The results of the analysis of the soil used for the raising of the seedlings showed complete absence of Al. This is an indication that the only source of Al in the study came from the Al treatments (T3-T7). Generally, as can be seen from the eleven parameters studied, Al treatment (which was the main focus of the study) significantly reduced both the vegetative and reproductive growths of A. hybridus at the low concentration of 3 x 10-2 mM used. This is an indication that the popular local A. hybridus cultivar, “Inine oma” used for the study is sensitive to low Al concentrations and is placed under the Al-sensitive group based on the classification of some tropical plants by Osaka et al. (1997) according to their Al tolerance capacity. Other workers have reported sensitivities of some plants to low Al concentrations. While Taylor et al. (1998) reported that in cowpea a toxicity threshold was observed at 0.1 μM Al and complete growth inhibition recorded at > 40 μM concentrations; Aftab et al. (2010) observed that application of 0.01 and 0.10 mM Al concentrations on Artemisia annua resulted to a mild stress situation that gave rise to a higher production of artemisinin. This shows that low concentrations of Al could generate stress and reduce growth in some plants while it can enhance growth/ productivity in some others. A comparison between the effects of Al toxicity (T7) in relation to T5 and T6 treatments (which were the post-ly treatments) showed that there were more growth decreases among the vegetative parameters studied than the reproductive parameters. The difference in the decrease of number of leaves, fresh weight of shoot and dry weight of shoot was significant between T7 and T6 treatments, while it was non-significant for plant height, root length and fresh weight of root, respectively. This is thought to be caused by decrease in growth resulting from growing the plants, in T7 treatment for 21 d at 4.6 pH and higher H+ concentration of 2.5 x10-5 mol L-1, while the ones given T5 and T6 treatments were first of all grown in the same 3×10-2 mM Al (T7) for 18 d, at the same 4.6 pH, before they were transferred into the ly (T1) and ly (T2) solutions for only 72 h, at a pH of 5.8 and lower H+ concentration of 1.5 x 10-6 mol L-1. Stressing the plants for 18 d (3 d less than T7) gave results closer to T7 and led to the reduction in growth of all the parameters. Growing them in the ly treatments, (T5 and T6), at a less acidic pH and lower H+ concentration enabled them to recover non-significantly for plant height, root length and fresh weight of root but significantly for number of leaves, fresh weight of shoot and dry weight of shoot. The ameliorative role of ly was dose dependent with ly T2 showing better recovery than ly T1. Probably, if the plants were allowed to grow longer in the two ly concentrations, after Al treatment, better recovery results may have been achieved. This calls for further research to explore the possible effect of other concentrations of ly, as well as different durations of treatment, after Al stress. Aslanturk and Celik (2005) reported that ly had preventive effect on chromosome aberrations particularly at 1 and 3 μM concentrations but the effect decreased at 5 and 10 μM concentrations.
It is pertinent to emphasize that the observed decrease in growth should not be ascribed to the Al3+ in solution alone but also to the presence of relatively higher H+ concentration of 2.5 x10-5 mol L-1 based on the 4.6 pH of the solution. Soil Survey Division Staff- [SSDS] (1993) defined a solution with a pH of 4.6, as very strongly acidic. It is our view that the potential effects of H+ under acidic conditions in rhizotoxicity studies have not been given adequate attention leading to gross exaggeration of the toxic effects of Al on plant roots. Based on the fact that Al3+ becomes available in solutions, under very strong acidic conditions, pH<5.5 (Kinraide, 1990; 1997; Harter, 2007; Silva, 2012), observed that growth defects are often attributed purely to the effect of Al3+, completely ignoring the toxic effects of H+, which at 2.5 x10-5 mol L-1 could on its own totally disrupt growth, even in the absence of Al3+. Koyama et al. (2000) and Kidd and Proctor (2001), noted that despite the large amount of literature on Al toxicity, very little attention has been given to H+ toxicity, even though the latter is well known to be directly detrimental to root growth. Further reports indicated that as in Al toxicity, H+ toxicity is most severe in solutions of low ionic strength and low cation concentrations, and increasing the concentration of Ca2+ and other cations in the external solution reduces or even abolishes the detrimental effects of acidity (Marschner, 1991). Furthermore, it was emphasized that an evalua-tion of these low-pH effects is necessary for greater understanding and correct interpretation in studies of Al toxicity, but regrettably this is rarely undertaken (Lazof and Holland, 1999; Samac and Tesfaye, 2003). Our finding are the actual Al concentration in solution at the 3 × 10-2 mM used for the study, using the Aluminon protocol, was 1.85 mg/L further buttressed the view that H+ concentration of 2.5 x10-5 mol L-1 of the Al solution used at 4.6 pH, played a key role in the observed decrease in yield of A. hybridus. The poor performance of A. hybridus plants treated with sole ly and ly (T1 and T2), as well as the two pre-ly (T3 and T4) treatments, in almost all the parameters studied, could be attributed to the effect of the H+ concentration of 1.5 x10-6 mol L-1 at the 5.8 pH of the solutions which is higher than the H+ concentration of 4 x 10-7 mol L-1 at pH of 6.4 reported to be optimal for the growing of Amaranth plants (Singh and Whitehead, 1992; Palada and Chang, 2003). The 5.8 pH level was classified by SSDS (1993) as strongly acidic.
The observed dose dependent ameliorative effect of ly indicate that the Al stressed plants absorbed the supplemented ly antioxidants apparently because both the intrinsic and possibly stress-induced higher activities of antioxidant enzymes (not monitored) of the Al stressed plants were not enough for the plants to recover from the stress condition caused by Al toxicity. This view is supported by the findings of Gong et al. (2005), who stated that environmental stress increased the formation of reactive oxygen species (ROS) that oxidize membrane lipids, proteins and nucleic acids.
According to Sudhakar et al. (2001), under such conditions it is known that plants with high levels of antioxidants, either constitutive or induced, have been reported to have greater resistance to oxidative damage. Additionally, it has been reported that with increasing amounts of Al in the soil medium, a concomitant increase in the activities of catalase (CAT), peroxidase (POX) and superoxidase dismutase (SOD) antioxidant enzymes were observed in the exposed plants (Boscolo et al., 2003; Siminovicova, 2004; Sharma and Dubey, 2007; Zhen et al., 2007; Aftab et al., 2010). The decrease in growth of the post- Al stressed (T3 and T4 treatments) plants (though not statistically significant in almost all the parameters) were more than those of the pre-Al (T5 and T6) treatments. The possible reasons for this is that growing the non-stressed plants in T1 and T2 treatments for only 72 h before transferring them to Al for 18 d (T3 and T4) had insignificant ameliorative effects for almost all the parameters in comparison with the T1, T2 and Al treatments. As already discussed under T1 and T2 treatment, A. hybridus plants that are not under stress may not need supplemented antioxidants, probably because they are adequately protected by their intrinsic antioxidants. The essence of the antioxidants was to assist in mitigating stress conditions not to serve as growth stimulants. The 5.8 pH initial environment of the T3 and T4 treatments stopped the growth retardation process because of the H+ in solution, despite the fact that the duration of treatment was only 72 h. The transfer of the plants to Al (T7) condition that lasted for 18 d subjected the plants to a greater stress conditions caused by both Al3+ and H+, under a more acidic condition (pH 4.6 and H+ concentration of 5.9), as against the milder acidic condition of pH 5.8 and H+ concentration of 5.1 (for the T3 and T4 treatments). This can account for the non-significant difference between all the T3 and T4 treatments and Al (T7) treatment. In cases like plant height, fresh weight of shoot, dry weight of shoot and root length, growth suppression by Al was insignificantly lower than T3 treatment. The double growth decrease resulting from the initial growth in T1 and later in Al could be responsible for this. Additionally, the lower insignificant growth decrease of T4 over T3, treatment, as can be seen from leaf number, fresh weight of shoot, dry weight of shoot, root length, fresh weight of root; and dry weight of root can be as a result of the higher concentration of T1 in T2 being able to better prevent H+, arising from the 5.8 pH of ly, from retarding the growth of the plants. Further studies will be needed to confirm this assumption.
The response of A. hybridus to the various treatments with respect to dry weight of root merits a separate discussion from the other vegetative parameters. Though the responses to all the other treatments were non-significant, this is the only case where there was growth enhancement based on Al treatment. The reasons why this should occur only in this parameter is not known. The number of roots could not be counted because of the mesh of deformed lateral roots formed due to Al toxicity. Despite the extensive wealth of information available about the effects of Al toxicity on the roots of plants, controversies still persist, meaning that much still need to be understood. Zobel et al. (2007) reported both root growth inhibition and increase in root diameter in roots exposed to Al. It has been suggested that the tolerance level of a genotype may not always be based on the number of primary roots and root length because both parameters may have similar results in stressed and stress-free environments. In this case, root vigour, root growth pattern, total root area, or total root mass of the corresponding genotypes under stressed and stress- free environments may be considered (Famoso et al., 2010; Roy and Bhadra, 2014).
On the other hand, it has been reported by other workers that Al in low concentrations could stimulate growth in some aspects of growth and productivity and not in others. Ou-yang et al. (2014) found that with the increasing of AlCl3 concentration up to 1 mM, the fresh weight of cotyledons of Jatropha curcas had a little increase. On the other hand, the fresh weight of hypocotyls decreased gradually with increasing Al concentration up to 3 mM and the fresh weight of radicles showed a similar trend, but when Al concentration was higher than 1 mM, the development of radicles was completely suppressed. According to Aftab et al. (2010), the addition of Al to the soil medium significantly reduced the yield and growth of Artemisia annua plants, but the artemisinin content were higher when 0.10 mM Al was applied (51.8% more), as compared to untreated plants, however a decrease in artemisinin content was noted when further higher doses of Al were applied. The response of the reproductive parameters of length of inflorescence, number of inflorescence, fresh weight of inflorescence and dry weight of inflorescence, indicate that the decrease in the yield of these parameters by Al treatment were less than that of the other treatments, though the differences were not statistically significant. Though the concentration of Al in the different parts of the plant did not form part of this study, it is suspected that a possible reason for this could be reduced Al content of the reproductive parts of the plant which is probably a survival strategy to ensure conservation of the species germplasm, needed for the continuity of the species. The leaves, and not the reproductive parts, have been reported to be the major area of Al accumulation, especially in Al-accumulator species. Ma et al. (1997) remarked that Al ions translocate very slowly to the upper parts of plants and most plants contain no more than 0.2 mg Al g-1 dry mass. However, some plants known as Al accumulators, may contain over 10 times more Al in their leaves without any injury. Watanabe and Osaki (2002) reported that many Al accumulator species, especially woody plants, accumulate more than 10,000 mg Al kg-1 in the leaves, without injury to the plants. Specific classical examples of hyper-accumulators include tea plant (Camellia sinensis, in older leaves), Hydrangea and members of the Rubiaceae family; unfortunately, there is not much information in the literature as to mechanisms, cellular localization and chemical form of Al which accumulates in these plants (Vitorello et al., 2005). The issue of the optimal concentration of ly needed to achieve highly significant ameliorative effect merits discussion. From the results obtained, it is suggestive that the higher dose of (T2) given to plants after Al stress yielded better ameliorative results even for the short duration of 72 h and extending the duration may possibly yield even better result. This merits a follow up investigation. This ameliorative effect of 50 and 200 μM ly on A. hybridus differs markedly from the situation in human and animal antioxidant supplementation results, as perhaps expected. Lowe et al. (1999) reported that low doses (1 to 3 μM) ly or β-carotene protected DNA from damage induced by xanthine/xanthine oxidase. By increasing the concentration to (4 to 10 μM) of the test substances, the opposite effects were observed at concentrations that were higher than physiological concentrations seen in vivo (Hwang and Bowen, 2005).
The bioavailability of ly to the experimental plants in vitro is yet another curious issue. It is not known whether it is the cis or the trans-isomer of ly that could be more bioavailable to A. hybridus under Al stress. Again studies with human and animal models indicate that although the major form of ly in the diet is all trans-isomer, representing about 80 to 97% of total ly in tomatoes and related products (Nguen and Schwarz, 1998; Shi and Le Maguer, 2000), human blood and tissues contain mainly cis-isomers. Other studies have indicated that the cis-isomers are more bioavailable (Tyssandier et al., 2003; Khoo et al., 2011) in humans and animals. This situation may again be different in plants and merits investigation. On the other hand, based on the reports of Lee and Chen (2002) that heating ly at 60 and 80°C favoured the isomerization of ly, it is suspected that in the course of this study, that processing of the tomato paste at 60°C, for the extraction of ly may have converted the naturally occurring trans-isomer to the cis-isomer. It is then hypothesized that it was the cis-isomer that the stressed A. hybridus plants absorbed that resulted to the observed ameliorative effect after Al stress. Perhaps once inside the plants they could be re-converted to the trans-isomer that make up almost the total amount of ly in tomatoes as reported by (Nguen and Schwarz, 1998; Shi and Magner, 2000). This however needs to be investigated through further studies. The report of Krinsky (1998) has provided good insight into the mechanism of the action of ly. Though ly is generally known to be a lipophilic substance; which may raise the possible question about its solubility and hence bioavailability in the 1% ethanol solvent used in this study, following the protocol of Fiskesjo (1981). Van Breemen et al. (2008) reported that besides its radical reactions, ly has also been shown to up-regulate the so-called antioxidant response element (ARE). Cellular enzymes, like glutathione S-transferase, superoxide dismutase or quinone reductase, are activated by ly, resulting in another way of protecting cells against highly reactive oxygen species. Additionally, Linnewiel et al. (2009) found that hydrophilic oxidation products of carotenoids, rather than the intact lipophilic carotenoid molecules, were responsible for the stimulation of the ARE system, detected in vitro using LNCaP and MCF-7 cells.
Oxidized ly derivatives, built, due to the instability of these molecules, are present in tomatoes, as well as in human serum and tissues (Holzapfel et al., 2013). From the foregoing, it is opined that the A. hybridus plants were able to possibly benefit from the lipophilic as well as the hydrophilic oxidation products of ly. This is also an area for further investi-gation in the study of the ameliorative role of supplemented ly in plants.
The present study has raised more questions than answers in this apparent controversial area of study; where many questions are still begging for answers. With further studies, this area of supplementing Al stressed plants, or plants subjected to other forms of environmental stress, either through soil application or spraying of the aerial parts of plants, may be found to contribute positively towards the alleviation of environmental stress in plants, leading to the reduction of the food security challenges in sub Saharan Africa and perhaps other parts of the world.
CONCLUSION
In conclusion, this present study has shown that the local cultivar of A. hybridus (Inine oma), used for this study is an Al sensitive plant. The vegetative parts were more sensitive than the reproductive parts and as such recorded more decrease in growth. A. hybridus plants appear not to need ly while not under Al stress. Pre-lycopene treatment had little or no ameliorating effect on Al stressed plants while post-lycopene treatment recorded more ameliorating effect in most of the parameters studied though only few were statistically significant. The ameliorating effect of ly was found to be dose dependent which calls for further research to find the optimal dose. This area of study has received little or no scientific attention, apparently because plants are known to possess their intrinsic antioxidants for fighting ROS. However, the fact that crop losses (even by elite cultivars) continue to be recorded due to crops succumbing to diverse and complex environmentally based stresses; argues strongly in favour of exploring alternative ways of improving on the capacity of plants to withstand stress situations. Antioxidant supplementation may turn out to be equally beneficial to plants under stress, similar to the enormous role it has been playing in ameliorating stress conditions in man and animals.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interest.
REFERENCES
| Aftab T, Khan MMA, Idrees M, Naeem M, Moinuddin (2010). Effects of aluminium exposures on growth, photosynthetic efficiency, lipid peroxidation, antioxidant enzymes and artemisinin content of Artemisia annua L. J. Phytol. 2(8): 23-37. | ||||
|
Agarwal S, Rao AV (1998). Tomato lycopene and low density lipoprotein oxidation: a human dietary intervention study. Lipids 33: 981-984. Crossref |
||||
|
Aniol A, Gustafson JP (1984). Chromosomelo- cation of genes controlling aluminium tolerance in wheat, rye and triticale. Can. J. Genet. Cytol. 26: 701-705. Crossref |
||||
| Aslanturk OS, Celik TA (2005). Preventive effect of lycopene on chromosome aberrations in Allium cepa. Pakistan J. Biol. Sci. 8(6):482-486 | ||||
| Association of Official Analytical Chemists (2005). Official Methods of Analysis of AOAC International (18th Ed.). Gaithersburg, Maryland, USA. 983 pp. | ||||
|
Barceló J, Poschenrieder C (2002). Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: a review. Environ. Exp. Bot. 48:75-92. Crossref |
||||
|
Barceló J, Poschenrieder C, Vázquez MD Gunsé B (1996). Aluminium phyto-toxicity. A challenge for plant scientists. Fertil. Res.43:217-23. Crossref |
||||
|
Boscolo PRS, Menossi M, Jorge RA (2003). Aluminium induced oxidative stress in maize. Phytochemistry 62:181-189. Crossref |
||||
|
Carvalho MM, Andrew CS, Edwards DG Asher CJ (1980). Comparative performances of six Stylosanthes species in three acid soils. Aust. J. Agric. Res. 31:61-76. Crossref |
||||
|
Cazzonelli CI (2011). Carotenoids in nature: insights from plants and beyond. Funct. Plant. Biol. 38: 833-847. Crossref |
||||
| Clarkson DT (1969). Metabolic aspects of aluminium toxicity and some possible mechanisms for resistance. In: Rorison IH, editor. Ecological Aspects of the Mineral Nutrition in Plants. Oxford: Blackwell. pp. 38-397. | ||||
|
Darko E, Ambrus H, Stefanovits-Banyai E, Fodor J, Bakos F, Barnabas B (2004). Aluminium toxicity, Al tolerance and oxidative stress in an Al-sensitive wheat genotype and in Al-tolerant lines developed by in vitro microspore selection. Plant Sci. 166: 583-591. Crossref |
||||
|
Exley C (2004). The pro-oxidant activity of aluminium. Free Radic. Biol. Med. 36:380-387. Crossref |
||||
|
Famoso AN, Clark RT, Shaff JE, Craft E, McCouch SR, Kochian LV (2010). Development of a novel aluminum tolerance phenotyping platform used for comparisons of cereal aluminum tolerance and investigations into rice aluminum tolerance mechanisms. Plant Physiol. 153(4): 1678-1691. Crossref |
||||
|
Fatur T, Tusek M, Falnoga I, Scancar J, Lah TT, Filipic M (2002). DNA damage and metallothionein synthesis in human hepatoma cells (HepG2) exposed to cadmium. Food Chem. Toxicol. 40: 1069-1076. Crossref |
||||
|
Fiskesjo G (1981). The Allium test as a standard in environmental monitoring. Hereditas 102:99-112. Crossref |
||||
|
Foy CD (1992). Soil chemical factors limiting plant root growth, In: Hatfield J.L., Stewart B.A. (Eds.), Advances in Soil Sciences: Limitations to Plant Root Growth, Vol. 19, Springer Verlag, New York. pp. 97-149. Crossref |
||||
|
Gong H, Zhu X, Chen K, Wang S, Zhang C (2005). Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 169: 313-321. Crossref |
||||
| Hanachi P, Golkho SH (2009). Using HPLC to Determine the Composition and Antioxidant Activity of Berberis vulgaris. Eur. J. Sci. Res. 29(1): 47-54. | ||||
| Harter RD (2007). Acid soils of the tropics. | ||||
| Hede AR, Skovmand B, Lopez-Csati J (2001). Acid soils and aluminium toxicity. In: Reynolds, M.P., Ortiz-Monaterio, J.I., McNab, A. (Eds.), Application of physiology in wheat breeding. D.F. CIMMYT, Mexico. pp. 172-182. | ||||
| Hoagland DR, Arnon DI (1950). The water-culture method for growing plants without soil. California Agricultural Experiment Station Circular 347:1-32. | ||||
|
Hodis HN, Mack WJ, LaBree L, Cashin-Hemphill L, Sevanian A, Johnson R, Azen SP (1995). Serial coronary angiographic evidence that antioxidant vitamin intake reduces progression of coronary artery atherosclerosis. J. Am. Med. Assoc. 273:1849-1854. Crossref |
||||
|
Holzapfel NP, Holzapfel BM, Champ S, Feldthusen J, Clements J, Hutmacher DW (2013). The potential role of lycopene for the prevention and therapy of prostate cancer: from molecular mechanisms to clinical evidence. Int. J. Mol. Sci.14:14620-14646. Crossref |
||||
|
Hwang ES, Bowen PE (2005). Effects of lycopene and tomato paste extracts on DNA and lipid oxidation in LNCaP human prostate cancer cells. BioFactors 23:97-105. Crossref |
||||
|
Khoo HE, Prasad K N, Kong KW, Jiang Y, Ismail A (2011). Carotenoids and Their Isomers: Colour pigments in fruits and vegetables. Molecules 16: 1710-1738. Crossref |
||||
| Kidd PS, Proctor J (2001). Why plants grow poorly on very acid soils: are ecologists missing the obvious? J. Exp. Bot. 52:791-799. | ||||
|
Kinraide TB (1990). Assessing the rhizotoxicity of the aluminate ion, Al(OH)4. Plant Physiol. 93:1620-1625. Crossref |
||||
| Kinraide TB (1991). Identity of the rhizotoxic aluminium species. Plant Soil 134:167-178. | ||||
|
Kinraide TB (1997). Reconsidering the rhizotoxicity of hydroxyl, sulphate and fluoride complexes of aluminium. J. Exp. Bot. 48: 1115-1124. Crossref |
||||
|
Kochian LV (1995). Cellular mechanisms of aluminium toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46:237-260. Crossref |
||||
|
Kochian LV, Hoekenga OA, Pineros MA (2004). How do crop plants tolerate acid soils? Mechanisms of aluminium tolerance and phosphorus efficiency. Ann. Rev. Plant Biol. 55:459-493. Crossref |
||||
|
Koyama H, Kawamura A, Kihara T, Hara T, Takita E, Shibata D (2000). Overexpression of mitochondrial citrate synthase in Arabidopsis thaliana improved growth on a phosphorus-limited soil. Plant Cell Physiol. 41:1030-1037. Crossref |
||||
|
Krinsky NI (1998). The antioxidant and biological properties of the carotenoids. Ann. New York Acad. Sci. 854:443-447. Crossref |
||||
|
Lazof DB, Holland MJ (1999). Evaluation of the aluminium-induced root growth inhibition in isolation from low pH effects in Glycine max, Pisum sativum and Phaseolus vulgaris. Aust. J. Plant Physiol. 26:147-157. Crossref |
||||
|
Lee MT, Chen BH (2002) Stability of lycopene during heating and illumination in a model system. Food Chem. 78:425-432. Crossref |
||||
|
Le Noble ME, Blevins DG, Sharp RE, Cumbie BG (1996). Prevention of aluminium toxicity with supplemental boron. I. Maintenance of root elongation and cellular structure. Plant. Cell Environ. 19: 1132-1142. Crossref |
||||
|
Linnewiel K, Ernst H, Caris-Veyrat C, Ben-Dor A, Kampf A, Salman H, Danilenko M, Levy J, Sharoni Y (2009). Structure activity relationship of carotenoid derivatives in activation of the electrophile/antioxidant response element transcription system. Free Radic. Biol. Med. 47: 659-667. Crossref |
||||
|
Lowe GM, Booth LA, Young AJ, Bilton RF (1999). Lycopene and β-carotene protect against oxidative damage in HT29 cells at low concentrations but rapidly lose this capacity at higher doses. Free Radic. Res. 30:141-151. Crossref |
||||
|
Ma JF, Zheng SJ, Matsumoto H, Hiradate S (1997). Detoxifying aluminum with buck- wheat. Nature 390:569-570. Crossref |
||||
|
Maron LG, Kirst M, Mao C, Milner MJ, Menossi M, Kochian LV (2008). Transcriptional profiling of aluminum toxicity and tolerance responses in maize roots. New Phytol. 179: 116-128. Crossref |
||||
| Marschner H (1991). Mechanisms of adaptation of plants to acid soils. Plant Soil 134:11-20. | ||||
|
Matsumoto H (2000). Cell biology of aluminium toxicity and tolerance in higher plants. Int. Rev. Cytol. 200:1-47. Crossref |
||||
| Matsumoto H, Hirasawa F, Torkai H, Takahasi E (1976). Localisation of absorbed aluminium in pea roots and its binding to nucleic acids. Plant Cell Physiol. 17: 127-137. | ||||
|
Miller NJ, Sampson J, Candeias LP, Bramley PM, Rice-Evans CA (1996). Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 384: 240-246. Crossref |
||||
|
Mohan Murati AV, Suprava J, Kamal KP, Brahma BP (2008). Aluminium induced oxidative stress and DNA damage in root cells of Allium cepa L. Ecotox. Environ. Saf. 70: 300-310. Crossref |
||||
|
Mossor-Pietraszewska T (2001). Effect of aluminium on plant growth and metabolism. Acta Biochemica Polonica 48(3): 673-686. PMid:11833776 |
||||
|
Nguen ML, Schwarz SJ (1998). Lycopene stability during food processing. Proc. Soc. Exp. Boil. Med. 218:101-105. Crossref |
||||
|
Osaki M, Watanabe T, Tadano T (1997). Beneficial Effect of Aluminium on Growth of Plants Adapted to Low pH Soils. Soil Sci. Plant Nutr. 43: 551-563. Crossref |
||||
|
Ou-yang C, Gao S, Mei L, Chung TW, Tang L, Wang S Chen F (2014). Effects of aluminium toxicity on the growth and antioxidant status in Jatropha curcas seedlings. J. Med. Plants Res. 8:178-185 Crossref |
||||
| Palada MC, Chang LC (2003). AVRDC, International Cooperator's Guide Suggested Cultural Practices for Vegetable Amaranth. www.avrdc.org. Accessed July 15, 2014. | ||||
|
Polívka T, Frank HA (2010). Molecular factors controlling photosynthetic light harvesting by carotenoids. Acc. Chem. Res. 43: 1125-1134. Crossref |
||||
|
Rao AV, Agarwal S (1998). Bioavailability and in vivo antioxidant properties of lycopene from tomato products and their possible role in the prevention of cancer. Nutr. Cancer 31:199-203. Crossref |
||||
|
Rao AV, Agarwal S (2000). Role of antioxidant lycopene in cancer and heart disease. J. Am. Coll. Nutr.19: 563-569. Crossref |
||||
|
Rhee KS (1978). Minimization of further lipid peroxidation in the distillation 2-thiobarbituric acid test of fish and meat. J. Food Sci. 43:1776-1778. Crossref |
||||
|
Roy AK, Sharma A, Talukder G (1988). Some aspects of aluminium toxicity in plants. Bot. Rev.54: 145-177. Crossref |
||||
|
Roy B, Bhadra S (2014). Effect of toxic levels of aluminium on seedling parameters of rice (Oryza sativa L.) under hydroponic culture. Rice Sci. 21(4):217-223. Crossref |
||||
|
Ryan PR, DiTomaso JM, Kochian LV (1993). Aluminium toxicity in roots. An investigation of spatial sensitivity and the role of root cap. J. Exp. Bot. 44:437-446. Crossref |
||||
|
Samac DA, Tesfaye M (2003) Plant improvement for tolerance to aluminium in acid soils - a review. Plant Cell Tissue Organ Cult. 75:189-207. Crossref |
||||
|
Sharma P, Dubey RS (2007). Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 26:2027-2038. Crossref |
||||
|
Shi J, Le Maguer J (2000). Lycopene in tomatoes: Chemical and physical properties affected by food processing. Crit. Rev. Biotechnol. 20:293-334. Crossref |
||||
| Shull KE (1960). Suggested modified aluminon method for aluminum determination. J. Am. Water Works Assoc. 52(6):779-785. | ||||
|
Silva S (2012). Aluminium toxicity targets in plants. J. Bot. Article ID 219462. Crossref |
||||
|
Simonovicova M, Tamas L, Huttova J, Mistrık I (2004). Effect of aluminium on oxidative stress related enzymes activities in barley roots. Biol. Plant. 48:261-266. Crossref |
||||
| Singh BP, Whitehead WF (1992). Response of vegetable Amaranth to differing soil pH and moisture regimes. Acta Hort. (ISHS) 318:225-230. | ||||
|
Sivaguru M, Horst WJ (1998). The distal part of the transition zone is the most aluminium-sensitive apical root zone of Zea mays L. Plant Physiol. 116:155-163. Crossref |
||||
| Soil Survey Division Staff (SSDS) (1993). Soil Survey Manual. Agricultural Handbook No.:18 USDA-NRCS. US Government Printing Office, Washington, DC, USA. 438p. | ||||
|
Sudhakar C, Lakshmi S, Giridarakumar S (2001). Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci. 161:613-619. Crossref |
||||
|
Taylor GJ, Blamey FPC, Edwards DG (1998). Antagonistic and synergistic interactions between aluminium and manganese on growth of Vigna unguiculata at low ionic strength. Physiol. Plant 104:183-194. Crossref |
||||
| Tyssandier V, Reboul E, Dumas JF, Bouteloup-Demange C, Armand M, Marcand J, Sallas M, Borel P (2003). Processing of vegetable-borne carotenoids in the human stomach and duodenum. Am. J. Physiol. Gastrointest. Liver Physiol. 284(6):G913-G923. | ||||
|
Van Breemen RB, Pajkovic N (2008). Multi-targeted therapy of cancer by lycopene. Cancer Lett. 269:339-351. Crossref |
||||
|
Vitorello VA, Capaldi FR, Stefanuto VA (2005). Recent advances in aluminium toxicity and resistance in higher plants. Braz. J. Plant Physiol. 17:127-143. Crossref |
||||
|
Walter MH, Floss DS, Strack D (2010). Apocarotenoids: hormones, mycorrhizal metabolites and aroma volatiles. Planta 232:1-17. Crossref |
||||
|
Watanabe T, Osaki M (2002). Mechanisms of adaptation to high aluminium condition in native plant species growing in acid soils: A review. Commun. Soil Sci. Plant Anal. 33(7&8):1247-1260. Crossref |
||||
|
Yamamoto Y, Kobayashi Y, Matsumoto H (2001). Lipid peroxidation is an early symptom triggered by aluminium, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 125:199-208. Crossref |
||||
|
Yaping Z, Suping Q, Wenli Y, Zheng X, Hong S, Side Y, Dapu W (2002). Antioxidant activity of lycopene extracted from tomato paste towards trichloromethyl peroxyl radical 3CCl3O2. Food Chem. 77:209-212. Crossref |
||||
|
Zatta P, Kiss T, Suwalsky M, Berthon G (2002). Aluminium(III) as a promoter of cellular oxidation. Coord. Chem. Rev. 228:271-284. Crossref |
||||
|
Zhen Y, Qi JL, Wang SS, Su J, Xu GH, Zhang MS, Miao L, Peng XX, Tian D, Yang YH (2007). Comparative proteome analysis of differentially expressed proteins induced by Al toxicity in soybean. Physiol. Plant 131:542-554. Crossref |
||||
|
Zobel RW, Kinraide TB, Baligar VC (2007). Fine root diameters can change in response to changes in nutrient concentrations. Plant Soil 297: 243-254. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


