Full Length Research Paper

ABSTRACT
Plant growth promoting rhizobacteria (PGPR) are known to influence plant growing both by direct and/or indirect mechanisms. This study aimed to establish PGPR profile of 15 bacteria isolated from maize (Zea mays L.) rhizosphere in Benin. These rhizobacteria were screened in vitro for the plant growth promoting traits like production of indole acetic acid (IAA), ammonia (NH3), hydrogen cyanide (HCN), catalase, exopolysaccharides and antifungal activity against phytopathogenic fungi for example Fusarium verticillioides, that is an important maize pathogenic. Most rhizobacteria strains were found to produce catalase (100%), exopolysaccharides (100%), ammonia (86.66%), hydrogen cyanide (80%) and indole acetic acid (60%). Pseudomonas putida, Pseudomonas fluorescens and Azospirillum lipoferum have highly produced many of the investigated metabolites. Streptomyces hygroscopicus, Streptomyces fasciculatus, Pseudomonas aeruginosa, P. putida, P. fluorescens and A. lipoferum inhibited mycelial growth of F. verticillioides and Aspergillus ochraceus. P. fluorescens and P. aeruginosa were highly antagonistic against F. verticillioides (52.24% of mycelial growth inhibition) and A. ochraceus (58.33% of mycelial growth inhibition). These results suggest the possibility to use these rhizobacteria as biological fertilization to increase maize yield and the biological control of F. verticillioides and A. ochraceus.
Key words: Rhizobacteria, Plant growth promoting rhizobacteria (PGPR), antifungal activities, biological control, Benin.
Abbreviation: IAA, Indole acetic acid; HCN, hydrogen cyanide; PGPR, plant growth promoting rhizobacteria; ISR, Induced Systemic Resistance; EPS, exopolysaccharides; IPA, indole pyruvic acid.INTRODUCTION
Agriculture is increasingly dependent on the use of chemical fertilizers, growth regulators and pesticides to increase yield. Continued use of such chemical causes wide spread problems in environmental pollution, animal and human health, destruction of natural biological communities and modifies natural nutrient recycling (Karuppiah and Rajaram, 2011). One of the solutions to these problems is the use of bioresources to replace the chemical products. To face this situation, the use of bio-resources to replace these synthesis products is increasingly considered. Among these bio-resources, the use of Rhizobacteria attracts more and more scientists’ attention. The rhizosphere is the area around and under the influence of plant roots and it is known to promote the growth of different prokaryotic and eukaryotic microorganisms (Wahyudi et al., 2011; Cardon and Gage, 2006). Rhizospheric microorganisms can have positive, negative or neutral effects on plant growth and can be classified into following categories bacteria, fungi, actinomycetes, protozoa and algae among many others (Paul and Clark, 1996). Indeed, among these microorganisms, bacteria are the most studied and well known probably because they grow quickly and they have the ability to use a wide range of substances as carbon or nitrogen sources (Karuppiah and Rajaram, 2011). Some of the bacteria found in the rhizosphere include plant growth promoting rhizobacteria (PGPR). The term PGPR comprises of many bacterial species belonging to Pseudomonas, Azospirillum, Azotobacter, Klebsiella, Enterobacter, Alcaligenes, Arthrobacter, Burkholderia, Bacillus and Serratia genus (Saharan and Nehra, 2011) and which improve plant growth.
The mechanisms used by PGPR to promote plant growth are not fully understood. PGPR rhizobacteria use one or more direct or indirect mechanisms to improve the growth and health plants. The direct promotion by PGPR entails either providing the plant with a plant growth promoting substances that are synthesized by the bacterium or facilitating the uptake of certain plant nutrients from the environment. The indirect promotion of plant growth occurs when PGPR lessen or prevent the deleterious effect of one or more phytopathogenic microorganisms. These mechanisms can be triggered simultaneously or individually at different stages of the plant development (Ahmad et al., 2008). Several direct mechanisms such as atmospheric nitrogen (N2) fixation, minerals (for example phosphate) solubilization, antagonism against phytopathogenic microorganisms by production of siderophores, and the ability to produce or change the concentration of the phytohormones (indole-3-acetic acid, gibberellic acid, cytokinines and ethylene) have been reported (Nelson, 2004). Examples of indirect mechanisms include production of antibiotics, lytic enzymes, hydrogen cyanide, catalase, siderophores and competition for space and nutrients. These mechanisms include those involved in the host plant, pathogens andharmful microbes’ biological control (Khan et al., 2006). Thus, the plant health may not be affected in presence of pathogens microorganisms and favorable conditions to cause the diseases, because PGPR destroy the pathogen molecular signal or open plant induced systemic resistance (ISR).
The present study aimed to establish the PGPR profile of 15 rhizobacteria isolated from maize rhizosphere in Benin, in order to select the best PGPR.
MATERIALS AND METHODS
Microorganisms
The bacteria strains (15) used, were Azospirillum lipoferum, Pseudomonas putida, Pseudomonas aeruginosa, Pseudomonas fluorescens, Streptomyces hygroscopicus, Streptomyces rimosus, Streptomyces fasciculatus, Bacillus coagulans, Bacillus thuringiensis, Bacillus pumilus, Bacillus polymixa, Bacillus licheniformis, Bacillus lentus, Bacillus circulans and Bacillus firmus. These bacteria were previously isolated from the maize rhizosphere in Benin (West Africa), and were biochemically characterized by standard methods of Cappuccino and Sherman (1992) and Aneja (2003). The bacteria cultures were maintained on Muller Hinton (MH) broth with 10% of glycerol at -20°C. At the onset of the experiments all the strains were grown on their respective selective medium: Pseudomonas on King’s B medium, Bacillus on Nutrient agar, Azospirillum on NFb-medium and Streptomyces on Kuster-glycérol medium.
Screening of rhizobacteria for different metabolites
Indole acetic acid production
The production of indole acetic acid (IAA) was detected by adapting method described by Brick et al. (1991). Bacteria were grown in nutrient broth supplemented with L-Tryptophan (500 µg/ml) at 36 ± 2°C for 48 h. Fully grown cultures were centrifuged at 3000 rpm for 30 min. Two (2) ml of the supernatant were mixed with 2 drops of orthophosphoric acid and 4 ml of Salkowski reagent (50 ml of 35% perchloric acid, 1 ml of 0.5 M ferric chloride solution). The appearance of a pink color after 25 to 30 min, indicates IAA production. According to the color intensity, three point scale (+ = low intensity; ++ = Average intensity and +++ = high intensity) were used for analyze the results.
Hydrogen cyanide production
All rhizobacteria were investigated for the production of hydrogen cyanide (HCN) by the method of Lorck (1948). Bacterial streaks were made on nutrient agar enriched with glycine (4.4 g/l) poured into Petri dishes. The top of the plate was covered with a Whatman filter paper No. 1 soaked in 2% sodium carbonate in 0.5% picric acid solution. After sealed with parafilm, those plates were incubated at 36 ± 2°C for 4 days. The positive reaction of HCN production is shown by apparition of orange to red color. According to the color intensity, three point scale (+ = low intensity; ++ = Average intensity and +++ = high intensity) were used to analyze the results.
Catalase production
Catalase is an enzyme which hydrolyzes the hydrogen peroxide (H2O2) to oxygen and water. With a sterile pipette, a bacterial colony was dispersed in a drop of hydrogen peroxide previously deposited on a clean dry slide. The positive reaction is shown by an immediate release of oxygen bubbles forming foam solution (Riegel et al., 2006).
Exopolysaccharides production
The exopolysaccharides (EPS) production was estimated by modified method of Damery and Alexander (1969). Bacteria strains were inoculated into Erlenmeyer flask containing 25 ml of yeast extract mannitol (YEM) broth supplemented with 1% of carbon source (glucose). The inoculated flasks were incubated at 36 ± 2°C on a gyrorotatory shaker at 150 rpm for 72 h. After incubation, the bacterial suspension was centrifuged at 3500 rpm for 30 min and the supernatant was mixed with two volumes of chilled acetone. The crude polysaccharide developed was collected by centrifugation at 3500 × g for 30 min. The EPS was washed with distilled water and acetone (alternately), and weighed after overnight drying at 105°C.
Ammonia production
The production of ammonia by rhizobacteria was tested by peptone water method (Cappuccino and Sherman, 1992). The strains were inoculated in peptone water (10 ml) and incubated for 48 to 72 h at 36 ± 2°C. After incubation, Nessler’s reagent (0.5 ml) was added to each tube. The ammonia production was showed by development of brown to yellow color. According to the color intensity, three point scale (+ = low intensity; ++ = average intensity and +++ = high intensity) were used to analyze the results.
Antifungal activity of rhizobacteria
The antifungal activity of rhizobacteria against some phytopathogenic fungi namely Fusarium verticillioides, Aspergillus ochraceus, Aspergillus tamarii and Aspergillus wentii was assessed by dual culture method according to Kumar et al. (2002). A disk (5 mm diameter) was cut out from a young culture of the fungal pathogen and placed in the middle of a Petri dish of potato dextrose agar (PDA). Ten (10) µl of a rhizobacteria suspension (approx. 108 CFU/ml) were spotted 2 cm from and on opposite sides of the fungus infected disk. The control Petri dishes were realized by monocultures of each pathogen. The Petri dishes were incubated at 26 ± 1°C and checked for zones of inhibition of mycelium growth after seven (7) days when the fungal mycelium had reached the edge of the Petri dish. When the pathogen grew over the rhizobacteria, we concluded that rhizobacteria did not have antifungal activity. On the other hand, when fungal growth was restricted by the rhizobacteria, the rhizobacteria’s antifungal activity (percentage of inhibition) was calculated by the following formula:
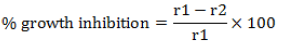
Where, r1 = diameter of pathogen growth in monoculture (control); r2 = diameter of pathogen growth in dual culture.
Data analysis and photo manipulation
The data were analyzed by using Microsoft Office Excel software version 2003. The graphs were realized by using the same software. Adobe Photoshop version 7.0 was used for photos treatment.
RESULTS
Production of plant growth promoting metabolites (PGPM) by rhizobacteria strains
Fifteen (15) rhizobacteria strains were evaluated for production of the plant growth promoting metabolites as catalase, ammonia, hydrogen cyanide, exopolysaccharides and indole acetic acid (IAA) as well as for antifungal properties. All the 15 strains had produced catalase, and exopolysaccharides (Figure 1), while 86.66, 80 and 60% of the strains had produced ammonia, hydrogen cyanide and IAA, respectively. Nine of the strains were found to have antifungal properties (Figure 1). Of the four bacteria genera (Bacillus, Streptomyces, Pseudomonas and Azospirillum) used for production of PGP metabolites searching, all the strains of Pseudomonas and Azospirillum were found to produce catalase, ammonia, hydrogen cyanide, IAA and exopolysaccharides. The production of plant growth promoting metabolites (PGPM) by different genera of the 15 rhizobacteria strains tested in this study is presented in Figure 2. All the species of Pseudomonas and Azospirillum tested were found to produce catalase, ammonia, hydrogen cyanide, indole acetic acid and exopolysaccharides.
All the species of Streptomyces and Bacillus have produced catalase and exopolysaccharides, while 87.5 and 66.7% of the Bacillus and Streptomyces species have respectively produced ammonia. Only 75 and 66.7% of Bacillus and Streptomyces species have respectively produced hydrogen cyanide, whereas 62.5% of Bacillus species have produced IAA but none of Streptomyces species have produced any IAA
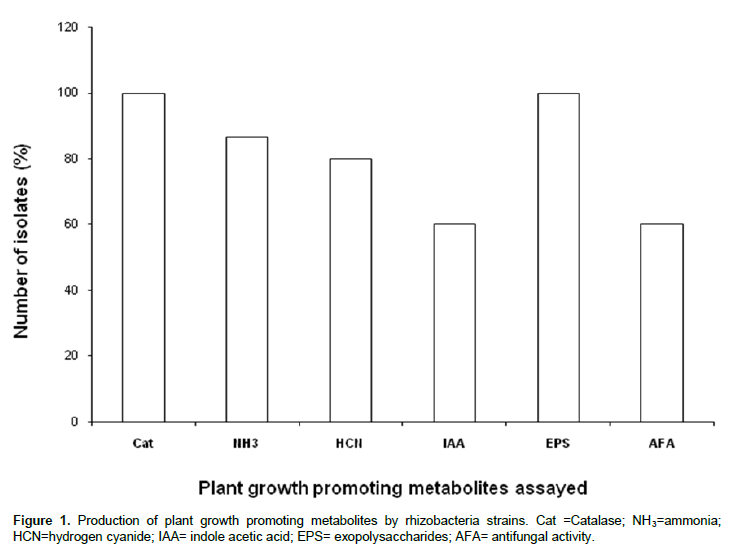
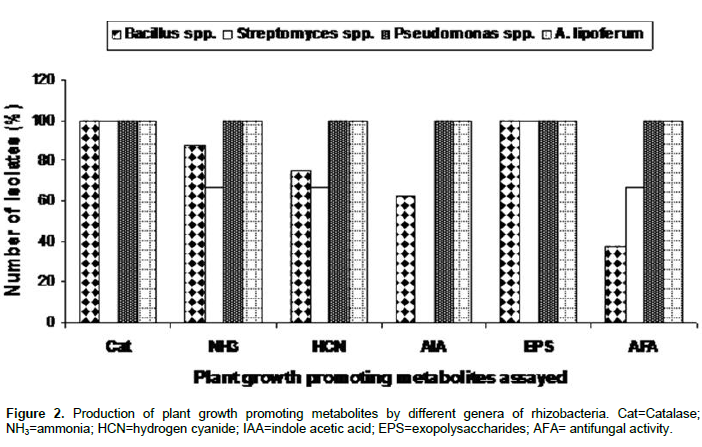
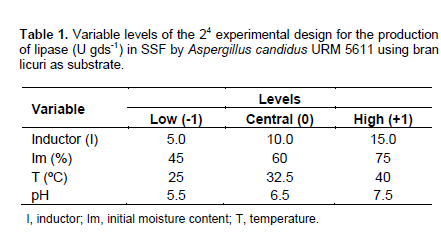
Table 1 provides a detailed profile of different rhizobacteria species for the parameters previously evaluated (catalase production, ammonia, cyanide hydrogen, indole acetic acid and exopolysaccharides) and an estimate the levels production of these parameters. When reading this table, we can observe that the different rhizobacteria do not have the same potential for production of PGP-metabolites. Thus, Pseudomonas putida and P. fluorescens highly produce ammonia, while Azospirillum lipoferum and B. thuringiensis have an average ammonia production. On the contrary, S. rimosus and B. lentus do not produce ammonia. The other strains lowly produced ammonia.
A. lipoferum, P. fluorescens and P. putida showed high production level of hydrogen cyanide and indole acetic acid. S. rimosus, B. lentus, B. circulans, S. fasciculatus, S. rimosus, S. hygroscopicus, B. lentus, B. firmus and B. circulans have not produced the hydrogen cyanide and indole acetic acid. Apart from P. fluorescens that displays high production of exopolysaccharides, all the other rhizobacteria had low or average produced exopolysaccharides.
Antagonism assay of rhizobacteria strains against phytopathogenic fungi
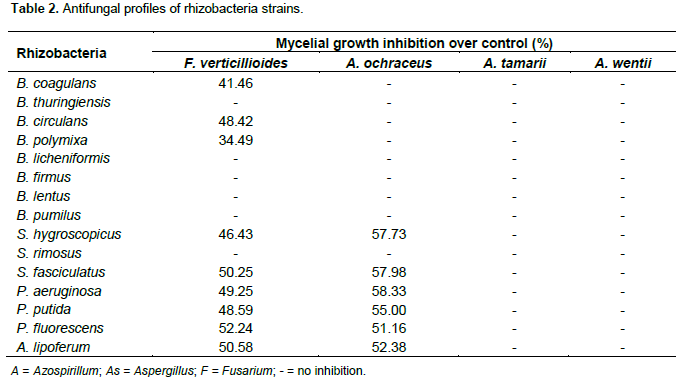
The antifungal properties of the rhizobacteria strains used in this study were tested against plant pathogenic: Fusarium verticillioides, Aspergillus Ochraceus, A. Tamarii and A. wentii. The growth of F. verticillioides was inhibited by S. hygroscopicus, S. fasciculatus, P. aeruginosa, P. putida, P. fluorescens and A. lipoferum to varying degrees. An inhibition zone was observed in Petri dishes for A. ochraceus co-inoculated with P. aeruginosa, S. hygroscopicus, S. fasciculatus, P. putida, P. fluorescens and A. lipoferum (Table 2). B. thuringiensis, B. licheniformis, B. firmus, B. lentus and S. rimosus did not inhibit the growth of any of the pathogenic fungi tested (Table 2). No antagonism was observed between rhizobacteria strains and A. tamarii and A. Wentii (Figure 3d). In opposite, S. hygroscopicus, S. fasciculatus, P. aeruginosa, P. putida, P. fluorescens and A. lipoferum have inhibited the normal A. ochraceus mycelial growth.
.png)
P. aeruginosa, S. hygroscopicus and S. fasciculatus have induced the mycelial growth inhibition of A. ochraceus respectively with the rate of 58.33, 57.98 and 57.73% (Table 2). Many rhizobacteria strains have presented the antagonism against F. verticillioides. Indeed, all rhizobacteria except B. thuringiensis, B. licheniformis, B. firmus, B. lentus (Figure 3c), B. pumilus and S. rimosus have inhibited the mycelial growth of F. verticillioides. S. fasciculatus (50.25%) (Figure 3b), A. lipoferum (50.58%) and P. fluorescens (52.24%) have strongly inhibited mycelial growth of F. verticillioides. It was remarked that all rhizobacteria display antagonistic effect to A. ochraceus which have also inhibited F. verticillioides.
DISCUSSION
Understanding the bacteria mechanisms of root coloni-zation and plant growth is crucial to improve the biological control methods against the plant pathogens and efficient use of plant growth promoting rhizobacteria (Kloepper et al., 1980). Indeed, PGPR properties investi gating the different rhizobacteria in the study revealed interesting results that illuminate their PGPR profiles. All rhizobacteria strains used in this study produced catalase. These strains are in the genera Bacillus, Pseudomonas, Azotobacter and Streptomyces. Previous studies by Rathaur et al. (2012) showed that Bacillus spp., Pseudomonas spp., and Azotobacter spp., isolated from Withania somnifera (Ashwagandha), were also able to produce catalase. The ability to produce catalase gives the bacteria an added advantage because it makes them potentially more resistant to environmental, mechanical and chemical stresses (Rathaur et al., 2012). The ability of PGPR to produce ammonia is considered as an important measure to influence indirectly plant growth (Karuppiah and Rajaram, 2011). About eighty six percent (86%) of tested rhizobacteria had produced ammonia (87.5% of Bacillus spp., 66.6% of Streptomyces spp., 100% of Pseudomonas spp., and Azospirillum spp.,). P. putida and P. fluorescens displayed high production of ammonia, while Azospirillum lipoferum and B. thuringiensis had average production. In studies by other researchers, ammonia production was detected in 95% of strains isolated from the rhizosphere of rice, mangroves and soils contaminated by effluent (Samuel and Muthukkaruppan, 2011, Joseph et al., 2007). B. subtilis MA-2 and P. fluorescens MA-4 known to be real producers of ammonia were shown to significantly increase the biomass of medicinal and aromatic plants such as geranium (Mishra et al., 2008). All the studied strains have produced low to moderate amount of exopolysaccharides, except P. fluorescens that displayed high production.
Exopolysaccharides (EPS) produced by rhizobacteria join soil aggregates and change their porosity (Alami et al., 2000). Thus, the soil porosity which is directly associated to the transfer of water from the soil to plant roots, is influenced by bacterial activity. Bacterial EPS produced on root surfaces also contribute to the maintenance of the water flow required for photosynthetic activity and plant growth.
In addition, EPS can limit or retard the soil dessiccation under water stress conditions. Conversely, in water excess (flooding), EPS help to prevent the dispersion of clay soil (Henao and Mazeau, 2009). Only 60% of the strains were able to produce IAA (100% of Pseudomonas spp. and Azospirillum spp., 62.5% of Bacillus spp., 0% of Streptomyces spp.,).
A. lipoferum, P. fluorescens and P. putida have produced large amounts of IAA. Several bacterial species for example Bacillus spp., Pseudomonas spp., Azotobacter spp., Azospirillium spp., Phosphobacteria spp., Glucanoacetobacter spp., Aspergillus spp. and Penicillium spp. have been reported to produce IAA (Shobha and Kumudini, 2012, Wahyudi et al., 2011). IAA production by PGPR varies according to species and strains origin. This may also be influenced by growth conditions, stage of development and the substrate availability (Ashrafuzzaman et al., 2009). The IAA produced by rhizobacteria plays a major role in roots elongation and can directly promote roots growth by cells elongation and cells division stimulation or indirectly influences the ACC (Amino cyclopropane-1-carboxylate)-deaminase activity in rhizobacteria (Rathaur et al., 2012). In addition, Patten and Glick (2002) reported that low levels of IAA can stimulate roots elongation, while high levels of IAA stimulate the lateral and adventitious roots formation. The ACC-deaminase product by PGPR, hydrolyzes plant’s ACC, an immediate precursor of plant ethylene hormone, which prevents production of toxic ethylene levels (Patten and Glick, 2002). The ability of rhizobacteria to use tryptophan added to their culture medium is an important parameter to detect and quantify their IAA production. Tryptophan is the main precursor of the bacterial IAA biosynthesis via the Indole Pyruvic Acid (IPA) pathway (Patten and Glick, 1996). IAA is part of many secondary metabolites produced by rhizobacteria in abundance during the stationary phase, even in the absence of tryptophan (Wahyudi et al., 2011).
Another important trait of PGPR, that may indirectly influence the plant growth, is the production of hydrogen cyanide. Eighty percent (80%) of rhizobacteria tested produced hydrogen cyanide (100% of Pseudomonas spp., Azospirillum spp., 75% of Bacillus spp., 66.6% of Streptomyces spp.). In a study by Karuppiah and Rajaram (2011), 75% of Bacillus spp., isolated produced hydrogen cyanide, whereas only 40% of rhizobacteria isolated from bean (Phaseolus vulgaris L.) in France by Kumar et al. (2012) produced the same secondary metabolite. Many rhizobacteria displayed antagonism against phytopathogenic fungi: S. hygroscopicus, S. fasciculatus, P. aeruginosa, P. putida, P. fluorescens and A. lipoferum significantly (51.16 to 58.33%) inhibited the mycelial growth of A. ochraceus. All rhizobacteria except B. thuringiensis, B. licheniformis, B. firmus, B. lentus, B. pumilus and S. rimosus inhibited the F. verticillioides mycelia growth by 34.49 to 52.24%. Fatima et al. (2009) showed that Azospirillum WPR-42 and Azospirillum WM -3 isolated from wheat (Triticum turgidum) inhibited mycelial growth of Rhizoctonia solani by 55 and 75%, respectively. Wahyudi et al. (2011) also demonstred that isolates of Bacillus spp. inhibited mycelial growth of Sclerotium rolfsii and F. oxysporum by 8.33 and 25%, respectively. None of the rhizobacteria tested in this study were antagonistic against A. tamarii and A. wentii. This suggests that the modes of action and types of antifungal metabolites produced vary from a rhizobacteria to another (Williams and Asher, 1996). The reduction of fungal growth by some PGPR and the in vitro inhibition zones may be probably due to antifungal substances and/or lytic enzymes released by PGPR. Indeed, the synthesis of a wide range antibiotics is most common character associated with the ability of PGPR to preclude the plant pathogens proliferation (Mazurier et al., 2009). The antibiosis mechanism is well-known and perhaps most importantly used by PGPR to limit the pathogens’ invasion. It consists of direct inhibition of the pathogen growth through the antifungal metabolites and/or antibiotics production (Adam, 2008). B. subtilis strains produce a variety of powerful antifungal metabolites such as zwittermycine-A, kanosamine (Peypoux et al., 1999) and lipopeptides belonging to surfactin, iturin and the fengycin families (Rahman et al., 2007). Some antifungal molecules, produced by Pseudomonas spp, such as hydrogen cyanide (HCN), viscosamide, pyoluteorine, 2,4-diacetylphloroglucinol, pyrrolnitrin, phenazines and butyrolactone are also implicated in the biocontrol (Haas and Defago, 2005).
CONCLUSION
This study focused on establishing the PGPR profiles of bacteria isolated from maize rhizosphere in Benin. Most of the rhizobacteria strains were found to produce catalase, exopolysaccharides, ammonia, hydrogen cyanide and indole acetic acid. P. putida, P. fluorescens and A. lipoferum produced most of the investigated metabolites. These results suggest the possibility to use these rhizobacteria as biological fertilization to increase the maize yield. S. hygroscopicus, S. fasciculatus, P. aeruginosa, P. putida, P. fluorescens and A. lipoferum inhibited mycelial growth of F. verticillioides and A. ochraceus. P. fluorescens and P. aeruginosa were highly antagonistic against F. verticillioides and A. ochraceus and can be used in biological control of these phytopathogenic fungi.
CONFLICT OF INTERESTS
The authors declare that they have no competing interests.
ACKNOWLEDGEMENTS
The authors thanked the International Foundation of Science (IFS research grant No. C/5252-1) and West Africa Agricultural Productivity Programme (WAAPP/PPAAO) for funding this work.
REFERENCES
|
Adam A (2008). The systemic resistance induced in tomato and cucumber and stimulation of the lipoxygenase pathway by non-pathogenic rhizobacteria. Ph.D. Thesis, University of Liège. Liège. Belgium. p. 148. Ahmad F, Ahmad I, Khan MS (2008). Screening of free-living rhizospheric bacteria for their multiple growth promoting activities. Microbiol. Res. 163:173-181. |
|
|
Alami Y, Achouak W, Marol C, Heulin T (2000). Rhizosphere soil aggregation and plant growth promotion of sunflower by an EPS-producing Rhizobium sp. isolated from sunflower roots. Appl. Environ. Microbiol. 66:3393-3398. |
|
|
Aneja KR (2003). Experiments in Microbiology, Plant pathology and Biotechnology. 4th ed. New Age International Publishers, Daryaganj, New Delhi. |
|
|
Ashrafuzzaman M, Hossen FA, Ismail MR, Hoque MA, Islam ZM, Shahidullah SM, Meon S (2009). Efficiency of plant growth-promoting rhizobacteria (PGPR) for the enhancement of rice growth. Afr. J. Biotechnol. 8: 1247-1252. |
|
|
Brick JM, Bostock RM, Silverstone SE (1991). Rapid in situ assay for indoleacetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microbiol. 57: 535-538. |
|
|
Cappuccino JC, Sherman N (1992). Microbiology: A Laboratory Manual. Benjamin (ed). Cumming Publishing Company, New York. USA. pp. 125-179. |
|
|
Cardon ZJ, Gage DJ (2006). Resource exchange in the rhizosphere: molecular tools and the microbial perspective. Anim. Rev. Ecol. Evol. Syst. 37:459-488. |
|
|
Damery JT, Alexander M (1969). Physiological differences between effective and ineffective strains of Rhizobium. Soil. Sci. 108: 209-215. |
|
|
Fatima Z, Saleemi M, Zia M, Sultan T, Aslam M, Rehman R, Fayyaz Chaudhary M (2009). Antifungal activity of Plant Growth-Promoting Rhizobacteria isolates against rhizoctonia solani in wheat. Afr. J. biotechnol. 8:219-225. |
|
|
Haas D, Defago G (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3: 307-319. |
|
|
Henao LJ, Mazeau K (2009). Molecular modelling studies of clay-exopolysaccharide complexes: soil aggregation and water retention phenomena. Mat. Sci. Engin. 29: 2326-2332. |
|
|
Joseph B, Ranjan Patra R, Lawrence R (2007). Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). Int. J. Plant. Prod. 2: 141-152.Karuppiah P, Rajaram S (2011). Exploring the Potential of Chromium Reducing Bacillus sp. and there Plant Growth Promoting Activities. J. Microbiol. Res. 1:17-23. |
|
|
Khan MS, Zaidi A, Wani PA (2006). Role of phosphate solubilizing microorganisms in sustainable agriculture-a review. Agron Sustain. Dev. 27:29-43. |
|
|
Kloepper JW, Schroth MN, Miller TD (1980). Effects of rhizosphere colonization by plant growth promoting rhizobacteria on potato plant development and yield. Ecol. Epidemiol. 70:1078-1082 |
|
|
Kumar A, Kumar A, Devi S, Patil S, Payal C, Negi S (2012). Isolation, screening and characterization of bacteria from Rhizospheric soils for different plant growth promotion (PGP) activities: an in vitro study. Recent Res. Sci. Technol. 4:01-05. |
|
|
Kumar NR, Arasu VT, Gunasekaran P (2002). Genotyping of antifungal compounds producing plant promoting rhizobacteria, Pseudomonas fluorescens. Curr. Sci. 82:1463-466. |
|
|
Lorck H (1948). Production of hydrocyanic acid by bacteria. Physiol. Plantarum. 1:142-146. |
|
|
Mazurier S, Corberand T, Lemanceau P, Raaijmakers JM (2009). Phenazine antibiotics produced by fluorescent pseudomonads contribute to natural soil suppressiveness to Fusarium wilt. ISME J. 3: 977-991. |
|
|
Mishra PK, Mishra S, Selvakumar G, Bisht SC, Kundu S, Bisht JK, Gupta HS (2008). Characterization of a psychrotrophic plant growth promoting Pseudomonas PGERs17 (MTCC 9000) isolated from North Western Indian Himalayas. Annal. Microbiol. 58: 1-8. |
|
|
Nelson LM (2004). Plant growth promoting rhizobacteria (PGPR): Prospect for new inoculants. Crop Managt. doi: 10. 1094/CM-2004-0301-05-RV. |
|
|
Patten CL, Glick BR (1996). Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 42:207-220. |
|
|
Patten CL, Glick BR (2002). Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 68:3795-3801. |
|
|
Paul EA, Clark FE (1996). Soil Microbiology and Biochemistry. Academics Press, San Diego: CA, USA, p. 340. |
|
|
Peypoux F, Bonmatin JM, Wallach J (1999). Recent trends in the biochemistry of surfactin. Appl. Environ. Microbiol. 51: 553-563. |
|
|
Rathaur P, Ramteke PW, Waseem W, John SA (2012). Isolation and characterization of nickel and cadmium tolerant plant growth promoting rhizobacteria from rhizosphere of Withania somnifera. J. Biol. Environ. Sci. 6:253-261. |
|
|
Rahman M, Ano T, Shoda M (2007). Biofilm fermentation of iturin A by a recombinant strain of Bacillus subtilis 168. J. Biotechnol. 127: 503-507. |
|
|
Riegel P, Archambaud M, Clavé D, Vergnaud M (2006). Bactérie de culture et d'identification difficiles. Biomérieux, Paris, France. pp. 93-112. |
|
|
Saharan BS, Nehra V (2011). Plant Growth Promoting Rhizobacteria: A Critical Review. LSMR. 21: 1-30. |
|
|
Samuel S, Muthukkaruppan SM (2011). Characterization of plant growth promoting rhizobacteria and fungi associated with rice, mangrove and effluent contaminated soil. Curr. Bot. 2: 22-25. |
|
|
Shobha G, Kumudini BS (2012). Antagonistic effect of the newly isolated PGPR Bacillus spp. on Fusarium oxysporum. Int. J. Appl. Sci. Eng. Res. 1:463-474. |
|
|
Wahyudi AT, Astuti RP, Widyawati A, Meryandini A, Nawangsih AA (2011). Characterization of Bacillus sp. strains isolated from rhizosphere of soybean plants for their use as potential plant growth for promoting Rhizobacteria. J. Microbiol. Antimicrobial. 3:34-40. |
|
|
Williams GE, Asher MJC (1996). Selection of rhizobacteria for the control of Phythium ultimum and Aphanomyces cochiliodes on sugerbeet seedlings. Crop Prot. 15:479-486. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


