Full Length Research Paper

ABSTRACT
Common bean (Phaseolus vulgaris) is a rich source of protein, vitamins, and micronutrients and is an important crop for food security throughout Latin America, Asia, and Africa. Among tropical and subtropical regions of the world, where the majority of beans are grown, yield losses due to the pathogenic fungus Pseudocercospora griseola, causing angular leaf spot (ALS), can be as high as 80%. The strategic use of marker-assisted selection (MAS) to pyramid multiple resistance genes into a single genetic background with preferred morphological and cooking characteristics is being implemented by six research groups throughout East Africa that make up the African Bean Consortium (ABC). Identifying unique markers that are polymorphic among multiple parents is a major source of marker attrition. In this study, an illustration of how 22 DNA sequences physically linked to the Phg-2 ALS resistance locus were identified using the PhaseolusGenes marker database and checked for amplification and polymorphism among 16 ABC breeding parents are given. Only three polymorphic markers could be identified following this procedure; one (g796), showed a polymorphism present only in the ALS resistance donor, Mexico 54. The PCR protocol developed to identify the g796 polymorphism was validated among five laboratories. Furthermore, co-segregation analysis of the marker and ALS resistance phenotype in a population of 100 F2 individuals from the cross between French bean (that is, green bean) genotype Amy and ALS resistance donor Mexico 54 showed the marker is genetically linked (3 cM) to the Phg-2 locus, in addition to being physically linked. This study suggests that in the near future, genetic resequencing data of diverse common bean accessions, compiled within an easily accessible database, will facilitate identification of markers for MAS, marker/trait association, and candidate gene identification.
Key words: Angular leaf spot resistance, Phaseolus vulgaris, marker-assisted selection.
Abbreviation: ABC, African Bean Consortium; ALS, angular leaf spot; BLAST, basic local alignment search tool; InDel, insertion-deletion; MAS, marker-assisted selection; PAMP, pathogen associated molecular pattern; PCR, polymerase chain reaction; PTI, PAMP triggered immunity; RAPD, random amplification of polymorphic DNA; SCAR, sequence-characterized amplified fragment; SNP, single-nucleotide polymorphism; SSR, simple sequence repeat; STS, sequence-tagged site.INTRODUCTION
MATERIALS AND METHODS
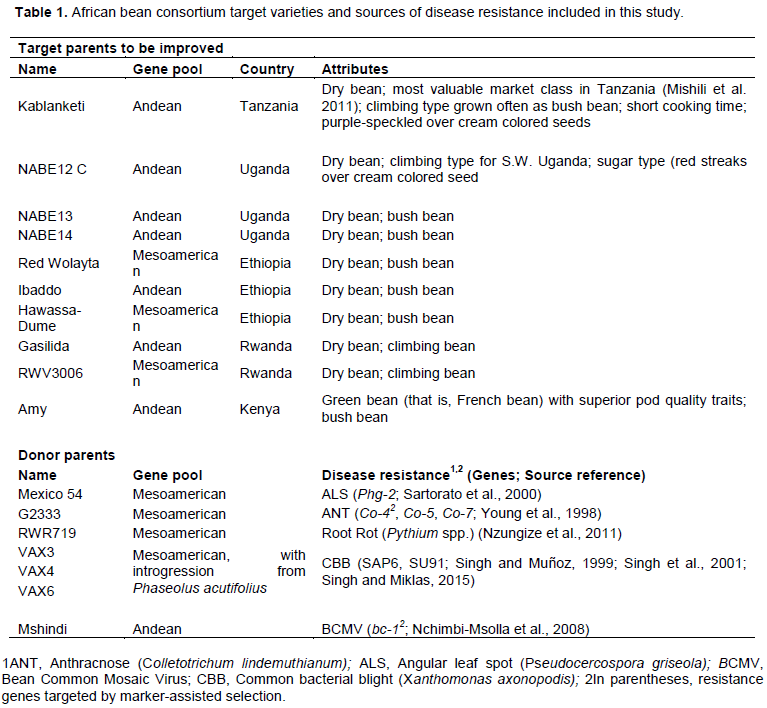
RESULTS

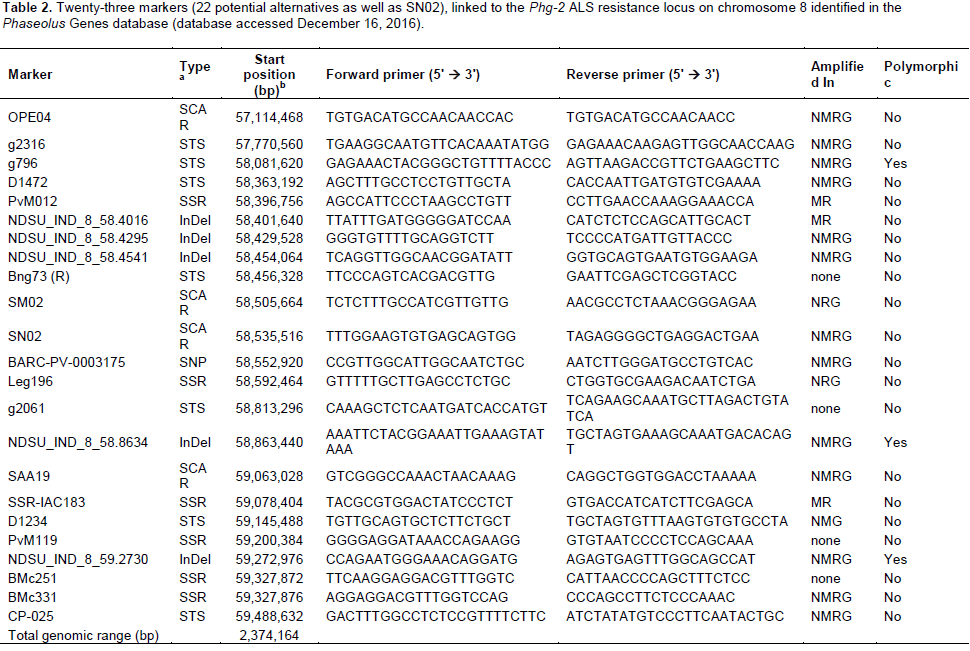

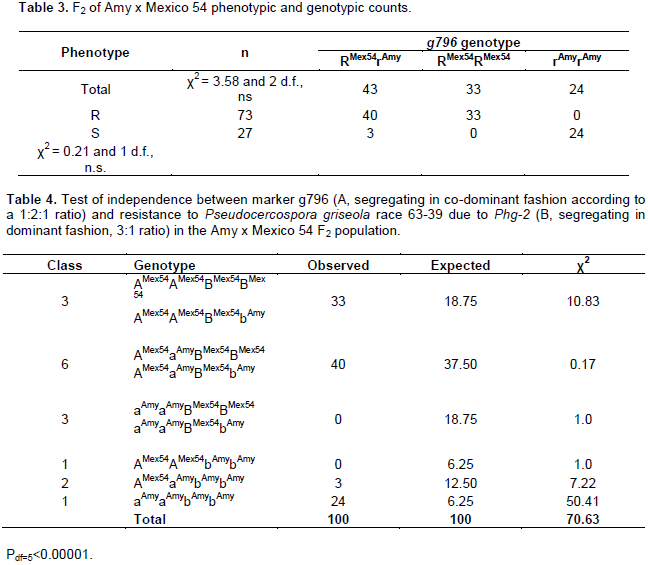
DISCUSSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ABBREVIATIONS
REFERENCES
|
Adam-Blondon A, Sévignac M, Bannerot H, Dron M (1994). SCAR, RAPD and RFLP markers tightly linked to a dominant gene (Are) conferring resistance to anthracnose in common bean. Theoretical Applied Genetics 88:865-870. |
|
|
Altschul S, Gish W, Miller W, Myers E, Lipman D (1990). Basic local alignment search tool. Journal of Molecular Biology 215:332-333. |
|
|
Aranda L, Porch TG, Bassett MJ, Lara L, Cregan PB (2014). Genetics and mapping of the Cl Gene for circumlineated pattern in common bean using AFLP-based bulk segregant analysis and SNP-based bidirectional selective genotyping. Journal of the American Society for Horticultural Science 139:213-218. |
|
|
Araya CM, Alleyne AT, Steadman JR, Eskridge KM, Coyne DP (2004). Phenotypic and genotypic characterization of Uromyces appendiculatus from Phaseolus vulgaris in the Americas. Plant Disease 88:830-836. |
|
|
Ariani A, Berny JC, Gepts P (2016). Genome-wide identification of SNPs and copy cumber variation in common bean (Phaseolus vulgaris L.) using genotyping-by-sequencing (GBS). Molecular Breeding 36:87-88. |
|
|
Ashraf M, Foolad MR (2013) Crop breeding for salt tolerance in the era of molecular markers and marker-assisted selection. Plant Breeding 132:10-20. |
|
|
Beaver J, Osorno J (2009). Achievements and limitations of contemporary common bean breeding using conventional and molecular approaches. Euphytica 168:145-175. |
|
|
Carvalho GA, Paula TJ, Alzate-Marin AL, Nietsche S, Barros EG, Moreira MA (1998). Inheritance of resistance to angular leaf spot of common bean in AND 277 to race 63–23 of Phaeoisariopsis griseola and identification of a RAPD marker linked to the resistance gene. Fitopatologia Brasileira 23:482-485. |
|
|
Coimbra-Gonçalves GK, Gonçalves-Vidigal MC, Coelho RT, Valentini G, Vidigal Filho PS (2016). Characterization and mapping of anthracnose resistance gene in Mesoamerican common bean cultivar Crioulo 159. Crop Science 56:2904-2915. |
|
|
Collard BCY, Mackill DJ (2008). Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philosophical Transactions of the Royal Society B: Biological Sciences 363(1491):557-572. |
|
|
Cortés AJ, Chavarro MC, Blair MW (2011). SNP marker diversity in common bean (Phaseolus vulgaris L.). Theoretical and Applied Genetics 123:827-845. |
|
|
Costa M, Tanure J, Arruda K, Carneiro J, Moreira M, Barros EG (2010). Development and characterization of common black bean lines resistant to anthracnose, rust and angular leaf spot in Brazil. Euphytica 176:149-156. |
|
|
Duncan RW, Gilbertson RL, and Singh SP (2012). Direct and marker-assisted selection for resistance to common bacterial blight in common bean. Crop Science 52:1511-1521. |
|
|
Durham KM, Xie W, Yu K, Pauls KP, Lee E, Navabi A (2013). Interaction of common bacterial blight quantitative trait loci in a resistant inter-cross population of common bean. Plant Breeding 132:658-666. |
|
|
Geffroy V, Sévignac M, De Oliveira J, Fouilloux G, Skroch P (2000). Inheritance of partial resistance against Colletotrichum lindemuthianum in Phaseolus vulgaris and co-localization of QTL with genes involved in specific resistance. Molecular Plant Molecular Interactions 13:287-296. |
|
|
Geffroy V, Sicard D, de Oliveira J, Sévignac M, Cohen S (1999). Identification of an ancestral resistance gene cluster involved in the coevolution process between Phaseolus vulgaris and its fungal pathogen Colletotrichum lindemuthianum. Molecular Plant Molecular Interactions 12:774-784. |
|
|
Gepts P, Aragão FJ, De Barros E, Blair MW, Brondani R, Broughton W, Galasso I, Hernández G, Kami J, Lariguet P, McClean P (2008). Genomics of Phaseolus beans, a major source of dietary protein and micronutrients in the tropics. Genomics of Tropical Crop Plants 113-143. Springer New York. |
|
|
Gepts P, Osborn TC, Rashka K, Bliss FA (1986). Phaseolin-protein variability in wild forms and landraces of the common bean (Phaseolus vulgaris): evidence for multiple centers of domestication. Economic Botany 40:451-468. |
|
|
Gonçalves-Vidigal M, Cruz A, Garcia A, Kami J, Vidigal Filho P (2011). Linkage mapping of the Phg-1 and Co-14 genes for resistance to angular leaf spot and anthracnose in the common bean cultivar AND 277. Theoretical and Applied Genetics 122:893-903. |
|
|
Gonçalves-Vidigal MC, Cruz AS, Lacanallo GF, Vidigal Filho PS, Sousa LL (2013). Co-segregation analysis and mapping of the anthracnose Co-10 and angular leaf spot Phg-ON disease-resistance genes in the common bean cultivar Ouro Negro. Theoretical and Applied Genetics, 126:2245-2255. |
|
|
Guzmán P, Gilbertson RL, Nodari R, Johnson WC, Temple SR (1995). Characterization of variability in the fungus Phaeoisariopsis griseola suggests coevolution with the common bean (Phaseolus vulgaris). Phytopathology 85:600-607. |
|
|
International Center for Tropical Agriculture (CIAT) (1987). Standard system for the evaluation of bean germplasm. Centro Internacional de Agricultura Tropical, Cali, Colombia. |
|
|
Johnson W, Guzmán P, Mandala D, Mkandawire A, Temple S (1997). Molecular tagging of the bc-3 gene for introgression into Andean common bean. Crop Science 37:248-254. |
|
|
Keller B, Manzanares C, Jara C, Lobaton JD, Studer B (2015). Fine-mapping of a major QTL controlling angular leaf spot resistance in common bean (Phaseolus vulgaris L.). Theoretical and Applied Genetics 128:813-826. |
|
|
Kelly JD, Gepts P, Miklas PN, Coyne DP (2003). Tagging and mapping of genes and QTL and molecular marker-assisted selection for traits of economic importance in bean and cowpea. Field Crops Research 82:135-154. |
|
|
Kwak M, Gepts P (2009). Structure of genetic diversity in the two major gene pools of common bean (Phaseolus vulgaris L., Fabaceae). Theoretical and Applied Genetics 118:979-992. |
|
|
Mahuku GS, Iglesias AM, Jara C (2009). Genetics of angular leaf spot resistance in the Andean common bean accession G5686 and identification of markers linked to the resistance genes. Euphytica 167:381-396. |
|
|
Mahuku GS, Henrıquez MA, Montoya C, Jara C, Teran H, Beebe S (2011). Inheritance and development of molecular markers linked to angular leaf spot resistance genes in the common bean accession G10909. Molecular Breeding 28:57-71 |
|
|
Melotto M, Afanador L, and Kelly J (1996). Development of a SCAR marker linked to the I gene in common bean. Genome 39:1216-1219. |
|
|
Meziadi C, Richard MM, Derquennes A, Thareau V, Blanchet S, Gratias A, Pflieger S, Geffroy V (2016). Development of molecular markers linked to disease resistance genes in common bean based on whole genome sequence. Plant Science 242:351-7. |
|
|
Miklas PN, Kelly JD, Beebe SE, Blair MW (2006). Common bean breeding for resistance against biotic and abiotic stresses: from classical to MAS breeding. Euphytica 147:106-131. |
|
|
Mishili FJ, Temu A, Fulton J, Lowenberg-DeBoer J (2011). Consumer preferences as drivers of the common bean trade in Tanzania: a marketing perspective. Journal of International Food and Agribusiness Marketing 23:110-127. |
|
|
Morrell PL, Buckler ES, Ross-Ibarra J (2012). Crop genomics: advances and applications. Nature Reviews Genetics 13:85-96. |
|
|
Namayanja A, Buruchara R, Mahuku G, Rubaihayo P, Kimani P (2006). Inheritance of resistance to angular leaf spot in common bean and validation of the utility of resistance linked markers for marker assisted selection outside the mapping population. Euphytica 151:361-369. |
|
|
Nchimbi-Msolla S, Misangu R, Mabagala R, Magayane F, Kweka S, Butler LM, Miklas PN, Myers JR (2008). 'Mshindi' kablanketi dry bean for East Arica. Annual Report Bean Improvement Cooperative 51:278-279. |
|
|
Nietsche S, Borem A, Carvalho GA, Rocha RC, Paula TJ (2000). RAPD and SCAR markers linked to a gene conferring resistance to angular leaf spot in common bean. Journal of Phytopathology 148:117-121. |
|
|
Nzungize J, Gepts P, Buruchara R, Male A, Ragama P, Busogoro JP, Baudoin JP (2011). Introgression of Pythium root rot resistance gene into Rwandan susceptible common bean cultivars. African Journal of Plant Science 5:193-200. |
|
|
Oblessuc PR, Baroni RM, Garcia AA, Chioratto AF, Carbonell SA, Camargo LE, Benchimol LL (2012). Mapping of angular leaf spot resistance QTL in common bean (Phaseolus vulgaris L.) under different environments. BMC genetics 13(1):50. |
|
|
Oblessuc PR, Cardoso Perseguini JMK, Baroni RM, Chiorato AF, Carbonell SAM (2013). Increasing the density of markers around a major QTL controlling resistance to angular leaf spot in common bean. Theoretical and Applied Genetics 126:2451-2465. |
|
|
Oblessuc PR, Francisco C, Melotto M (2015). The Co-4 locus on chromosome Pv08 contains a unique cluster of 18 COK-4 genes and is regulated by immune response in common bean. Theoretical and Applied Genetics 128:1193-1208. |
|
|
Okii D, Tukamuhabwa P, Kami J, Namayanja A, Paparu P, Ugen M, Gepts P (2014). The genetic diversity and population structure of common bean (Phaseolus vulgaris L.) germplasm in Uganda. African Journal of Biotechnology 29:2935-2949. |
|
|
Pastor-Corrales MA, Jara C, Singh SP (1998) Pathogenic variation in, sources of, and breeding for resistance to Phaeoisariopsis griseola causing angular leaf spot in common bean. Euphytica 103:161-171. |
|
|
Perseguini JMKC, Oblessuc PR, Rosa JRBF, Gomes KA, Chiorato AF, Carbonell SA, Garcia AA, Vianello RP, Benchimol-Reis LL (2016). Genome-wide association studies of anthracnose and angular leaf spot resistance in common bean (Phaseolus vulgaris L.). PLoS ONE,11:e0150506. |
|
|
Queiroz VT, Sousa CS, Costa MR, Sanglad DA, Arruda KMA, Souza TLPO, Ragagnin VA, de Barros EG, Moreira MA (2004). Development of SCAR markers linked to common bean angular leaf spot resistance genes. Annual Report Bean Improvement Cooperative 47:237-238. |
|
|
Reinprecht Y, Yadegari Z, Perry GE, Siddiqua M, Wright LC (2013). In silico comparison of genomic regions containing genes coding for enzymes and transcription factors for the phenylpropanoid pathway in Phaseolus vulgaris L. and Glycine max L. Merr. Frontiers in Plant Science 4:317. |
|
|
Rendón-Anaya M, Montero-Vargas JM, Saburido-Alvarez S, Vlasova A, Capella-Gutiérrez S, Ordaz-Ortiz JJ, Aguilar OM, Vianello-Brondani RP, Santalla M, Delaye L, Gabaldón T, Gepts P, Winkler R, Guigó R, Delgado-Salinas A, Herrera-Estrella A (2017). Genomic history of the origin and domestications of common bean in the Americas unveils its closest sister species. Genome Biology 18:60. |
|
|
Sartorato A, Nietsche S, de Barros EG, Moreira MA (2000). RAPD and SCAR markers linked to resistance gene to angular leaf spot in common beans. Fitopatologia Brasileira 25:637-642. |
|
|
Schmutz J, McClean P, Mamidi S, Wu G, Cannon S (2014). A reference genome for common bean and genome-wide analysis of dual domestications. Nature Genetics 46:707-713. |
|
|
Schwartz HF, Pastor-Corrales MA (1989). Bean production problems in the tropics. 2nd ed. International Center for Tropical Agriculture (CIAT), Cali, Colombia. |
|
|
Singh SP, Miklas PN (2015). Breeding common bean for resistance to common blight: A review. Crop Science 55:971-984. |
|
|
Singh SP, Mu-oz CG (1999). Resistance to common bacterial blight among Phaseolus species and common bean improvement. Crop Science 39:80-89. |
|
|
Singh SP, Mu-oz CG, Terán H (2001) Registration of common bacterial blight resistant dry bean germplasm VAX 1, VAX 3, and VAX 4. Crop Science 41: 275-276. |
|
|
Sousa LL, Gonçalves AO, Gonçalves-Vidigal MC, Lacanallo GF, Fernandez AC (2015). Genetic characterization and mapping of anthracnose resistance of common bean landrace cultivar Corinthiano. Crop Science 55:1900-1915. |
|
|
Souza TL, Ragagnin VA, Dessaune SN, Sanglard DA, Carneiro JE, Moreira MA, Barros EG (2014). DNA marker-assisted selection to pyramid rust resistance genes in "carioca" seeded common bean lines. Euphytica 199(3):303-316. |
|
|
Tryphone GM, Chilagane LA, Protas D, Kusolwa PM, Nchimbi-Msolla S (2013). Marker assisted selection for common bean diseases improvements in Tanzania: Prospects and future needs. InPlant breeding from laboratories to fields. InTech. |
|
|
Vlasova A, Capella-Gutiérrez S, Rendón-Anaya M, Hernández-O-ate M, Minoche AE (2016). Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biology 17:1-18. |
|
|
Voorrips RE (2002). MapChart: Software for the graphical presentation of linkage maps and QTLs. Journal of Heredity 93:77-78. |
|
|
Wortmann CS, Kirkby RA, Eledu CA, Allen DJ (1998). Atlas of common bean (Phaseolus vulgaris L.) production in Africa. International Center for Tropical Agriculture (CIAT), Cali, Colombia. |
|
|
Young RA, Melotto M, Nodari RO, Kelly JD (1998). Marker-assisted dissection of the oligogenic anthracnose resistance in the common bean cultivar, 'G 2333'. Theoretical Applied Genetics 96:87-94. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


