Full Length Research Paper

ABSTRACT
INTRODUCTION
Finger millet [Eleusine coracana (L.) Gaertn.], which is a small cereal crop that is indigenous to East Africa, is cultivated in arid and semi-arid areas of eastern Africa and south Asia (Sood et al., 2016). It is tetraploid (2n = 4x = 36) and belongs to the Poaceae family and Chloridoideae sub-family (Gimode et al., 2016). Finger millet is a highly nutritious cereal high in proteins, vitamins and minerals. Finger millet is one of the potential grain crops for food and nutritional security, climate resilient farming and agricultural diversification. Despite its significance as a subsistence crop, little attention has focused on the crop’s improvement programs, probably since it is considered with minimal monetary significance in contrast to maize, wheat and rice. Finger millet production and yield is constrained by lack of improved varieties, weeds, diseases and pests, limited uses, unpredictable markets, limited research and moisture stress in dry areas (Oduori, 2005). Varieties with new traits emerging as one will help the plant cope with adverse challenges associated with biotic and abiotic stress. The world’s increasing demand for finger millet offers a golden opportunity to develop efficient, quick and reproducible strategies and techniques for important finger millet local varieties addressing complex traits such as grain quality, biotic and abiotic stress resiliencies. Over the years, conventional breeding has been used for traits improvement in finger millet with some success. However, conventional plant breeding is tedious, time consuming and mostly dependent on environment (Miah et al., 2013). As an alternative, biotechnological techniques such as genetic engineering and genome editing techniques, which relies on the availability of in vitro plant regeneration systems, provides a powerful tool for genetic manipulation of finger millet. Genetic engineering of finger millet for improved varieties is stymied because of absence of a proficient plant tissue culture strategy with high regeneration frequency. To the authors’ knowledge, there is no report on tissue culture and transformation for African finger millet cultivars. Just like other cereals, finger millet regeneration can be accomplished through somatic embryogenesis and organogenesis. Somatic embryogenesis of finger millet is one of the most preferred approach due to production of large numbers of plantlets and its application for genetic transformation technology. Several factors such as the explant source, their developmental stage and plant growth regulators affect this morphogenetic route (Sudhakar et al., 2004). Finger millet has been considered a recalcitrant crop to tissue culture and genetic transformation. Only limited reports are available to date on regeneration and genetic transformation of finger millet finger millet varieties particularly of African origin (Gupta et al., 2017). In these reports, the low rate of embryo initiation, maturation, germination and development into plantlets often remain a major challenge. The present work reports a robust and reliable procedure for the establishment of an efficient and reproducible regeneration through system somatic embryogenesis of finger millet using shoot tips that will be useful for the genetic improvement of this crop.
MATERIALS AND METHODS
Plant material and explant preparation
Seeds of finger millet: GBK-043137, GBK-043128, GBK-043124, GBK-043122, GBK-043094 and GBK-043050 were obtained from Kenya Agricultural and Livestock Research Organization gene bank, at Muguga, Kenya. The seeds were soaked for 30 min in sterile distilled water to dehusk them, followed by surface sterilization with 70% ethanol and 20% sodium hypochlorite containing a few drops of Tween 20 for 20 min. Surface sterilized seed were rinsed three times with sterile distilled water and germinated aseptically on Murashige and Skoog (MS) basal medium (Murashige and Skoog, 1962), supplemented with 3% sucrose, 0.3% gelrite and pH 5.8. The culture bottles were incubated at 25±2°C in the dark for germination for three days.
Callus induction and somatic embryo development
Shoot tips (4 to 6 mm) from in vitro germinated plants were aseptically excised and cultured on callus induction medium (CIM) for six weeks in the dark. The CIM comprised of MS supplemented with 30% sucrose and various plant growth hormones (PGRs) either singly or in combinations comprising of NAA (2.0, 2.5, 3.0 and 4.0 mg/l); 2,4-D + BAP (1.5 + 0.5, 2.0 + 1.0, 2.5 + 1.5, 3.0 + 1.5 mg/l); 2,4-D + KN (1.5 + 0.5, 2.0 + 0.5, 2.5 + 1.0, 3.0 + 1.0 mg/l) and NAA + KN (1.5 + 0.5, 2.0 + 0.5, 2.5 + 1.0, 3.0 + 1.0 mg/l). Explants cultured on MS media without plant PGRs were taken as control. Cultures were incubated at 25±2°C in the dark for about four weeks to promote the formation of callus. The explants were subcultured to a fresh media after every two weeks. Calli that formed were transferred onto embryo induction medium (EIM) containing MS basal salts supplemented with 30 g/l sucrose and 1.75 mg/l of BAP and incubated at 25±2°C in dark for 8 weeks until differentiation into embryo-like structures were observed. The calli were subcultured onto fresh EIM medium after every two weeks.
Shoot proliferation and root formation
Mature embryos were cultured on shoot induction medium (SIM) comprising of MS supplemented with 30% sucrose and different concentrations of BAP (1, 1.5, 1.75 and 2 mg/L). The SIM was also used for elongation with subculture every two weeks up to four subcultures. The number of shoots formed were counted every four weeks and recorded. Shoots formed were cultured on root induction media (RIM) comprising of MS medium supplemented with 30% sucrose and 1.0 KN+0.25 NAA, 1.0 BAP + 0.25 NAA, 1.0 KN + 0.25 2,4-D and 1.0 BAP + 0.25 2,4-D for root formation and development for four weeks. The developed roots were observed and recorded.
Hardening and acclimatization
Rooted plantlets were rinsed with sterile distilled water to remove the excess media. The plantlets were transferred to the greenhouse for acclimatization on four media regimes: forest soil only, cocopeat, forest soil + sand + manure (2:1:1) and forest soil + sand + fertilizer (4:2:0.05). The plants were watered on regular intervals. Data on number of leaves, plant height and colour was collected after 2, 4 and on the sixth week from the day of hardening off. Survival of plantlets was recorded after 3 weeks [Survival plantlet (%) = (surviving plantlets/total plantlets) x 100).
Statistical analysis
All data were expressed as mean ± standard. The data were analysed using ANOVA with Minitab statistical computer software v.17. Means were separated using the Fisher’s protected LSD test at a confidence level of 95% (p ≤ 0.05).
RESULTS
Effects of plant growth regulators on callus induction
It was possible to geminate seeds of all the selected varieties in vitro using the MS hormone free medium, albeit with different gemination efficiencies. GBK-043137 exhibited the highest gemination efficiency of 75% followed by GBK-043050, GBK-043128, GBK-043094 and GBK-043124 with seed gemination efficiencies of 71, 66, 65 and 63%, respectively. Variety GBK-043122 had the least seed germination efficiencies of 40% (Table 1). There was significant difference between GBK-043137, GBK-043050, GBK-043128, GBK-043094 and GBK-043124, and GBK-043122 in terms of seed germination efficiencies on MS hormone free medium (Table 1) (p<0.05). However, for varieties GBK-043137, GBK-043050, GBK-043128, GBK-043094 and GBK-043124, there was no significant difference, in terms of seed germination efficiencies (Table 1) (p>0.05). All geminated plants exhibited normal phenotype (Figure 1a). It was possible to initiate calli in all the finger millet varieties at all the different concentrations and combinations of NAA, 2,4-D, BAP and kinetin, however with different callus formation frequencies. Different combinations and concentrations of NAA (2.0, 2.5, 3.0 and 4.0 mg/l); 2,4-D + BAP (1.5 + 0.5, 2.0 + 1.0, 2.5 + 1.5, 3.0 + 1.5 mg/l); 2,4-D + kinetin (1.5 + 0.5, 2.0 + 0.5, 2.5 + 1.0, 3.0 + 1.0 mg/l), NAA + kinetin (1.5 + 0.5, 2.0 + 0.5, 2.5 + 1.0, 3.0 + 1.0 mg/l) and control were tested for their ability to induce calli and determine best combination and concentration for calli induction of the finger millet varieties. All the treatments produced calli. The callus induction mean percentage ranged from 4.33±0.33 to 12.33±0.33 out of 60 explants based on the type and concentration of PGRs in the medium (Table 2). The highest callus formation was observed on medium supplemented with 2.5 mg/l 2,4-D + 1.5 mg/l BAP at 12.33±0.33 for variety GBK-043128 (Table 2a). This combination and concentration also had the best response for callus induction on all other varieties when compared with the others. GBK-043094 was the best responding variety with regards to callus induction in all media combinations and concentrations. Medium supplemented with NAA was the worst performing media with 4.33±0.33 callus formation. This medium formed callus like structures at the tip of explant and developed shoots within three days which were fast growing. These explants were subcultured; they turned brown after a week and eventually died off. The controls did not form any callus (Table 2a and b). Explants formed calli that were friable, soft, watery, white in colour, they developed gradually, slowly and were non-regenerative with unorganized morphology, destitute of nodular structures (Figure 1b). Following a month of culture, the greater part of the calli turned out to be light yellow in colour and embryogenic in nature with nodular development (Figure 1c). Explants did not show any change turned brown and eventually black and they died within three weeks of culture.



Regeneration of plants from callus
The rapidly growing friable calluses were subcultured onto MS medium supplemented with 1.75 mg/l of BAP for somatic embryo induction. Somatic embryogenic structures started to form after two weeks culture onto the EIM. The greenish compact and moderate developing calli that developed organized structures and inevitably displayed tissue differentiation were regarded as embryogenic (Figure 1d), as opposed to the non-embryogenic calli that were white or cream, friable and quickly developing (Figure 1e). Some calli of the finger millet varieties turned green and formed globular, heart, torpedo and cotyledonary stages of embryos which appeared progressively after 4 weeks on EIM medium. There was 100% transition from callus to somatic embryos for all finger millet varieties. After four weeks on EIM in light, the embryo maturation was observed with formation of small shoots (Figure 1f).
Shoot elongation and multiplication
The highest shoot formation frequency was achieved using MS supplemented with 1.75 mg/l on GBK-043094 and GBK-043137 both with a mean of 25.07±0.64 and 25.33±2.95, respectively (Table 3). This concentration of MS and 1.75 mg/l was also highly significant (P≤0.05) for GBK-043128, GBK-043124, GBK-043050, GBK-043094 and GBK-043137 with mean number of shoots at 23.93±0.74, 22.53±1.03, 23.93±0.74, 25.07±0.64 and 25.33±2.95 respectively, however a lower concentration of 1.5 mg/l was highly significant for GBK-043122. The lowest performing shooting media was 1.0 mg/l on GBK-043050 and 2.0 mg/l on GBK-043050 with a mean number of shoots of 9.80±1.33 and 9.80±2.40 respectively. The height of plants varied from 3 to 6 cm.
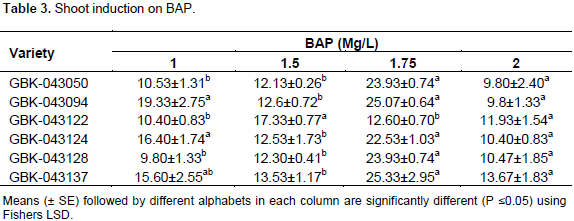
Root induction
Shoots (2 to 3 cm in height) cultured on MS media containing 1.0 KN + 0.25 NAA, 1.0 BAP + 0.25 NAA, 1.0 KN + 0.25 2,4-D and 1.0 BAP + 0.25 2,4-D showed root induction in all six finger millet varieties, after 4 weeks of incubation. The roots started forming within two weeks and proper root system was achieved in four weeks (Figure 1g) with high significant differences observed using 1.0 BAP + 0.25 NAA for varieties GBK-043128, GBK-043124, GBK-043094 and GBK-043050 with mean of 14.60±2.23, 15.00±2.2, 15.00±2.2 and 13.93±2.03, respectively. MS supplemented with 1.0 KN + 0.25 NAA was significantly higher for GBK-043137 and GBK-043122 with a mean of 10.87 ± 0.75 and 11.07 ± 0.93 respectively (Table 4). The best rooting hormone combinations not only had highest rooting but also 100% of the plants formed roots as compared to rest of the combinations.
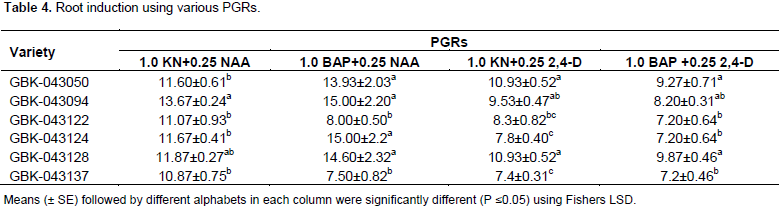
Hardening and acclimatization
Well-developed plantlets (5 to 6 cm in height) with more than two true leaves from all the regenerated varieties were hardened successfully in pots using four media regimes: forest soil only, cocopeat, forest soil + sand + manure in a ratio and forest soil + sand + fertilizer in 2:1:1 and 4:2:0.05 ratios with 97% survival rate (Figure 1h to i). The number of leaves for all the in vitro regenerated finger millet varieties varied between 6.43±1.30 and 11.92±0.96. The number of leaves for all the in vitro regenerated finger millet varieties was statistically, not significantly different at two and four six weeks for soil while all the rest were different (Table 5) across the media regimes. Following seven days of incubation in a growth chamber, all the pots were moved to regular environmental conditions with a 100% survival rate with a 100% survival rate. The somatic embryo-derived plantlets grew well and exhibited phenotypic homogeneity as compared to seed-derived field-grown finger millet plants (Figure 1g and h). The height of the plants at two weeks varied from 3 to 6 cm. The shortest plants at this period were those hardened on soil, sand and manure which were also significantly different on GBK-043124 (Table 6). At four weeks, the tallest plants were produced using cocopeat which was not significantly different across varieties (Figure 1b). The media with the tallest plants at six weeks was soil, sand and manure (52.33±5.56) on variety GBK-043128 which was significantly different while soil + sand + fertilizer produced the shortest plants (Table 6).
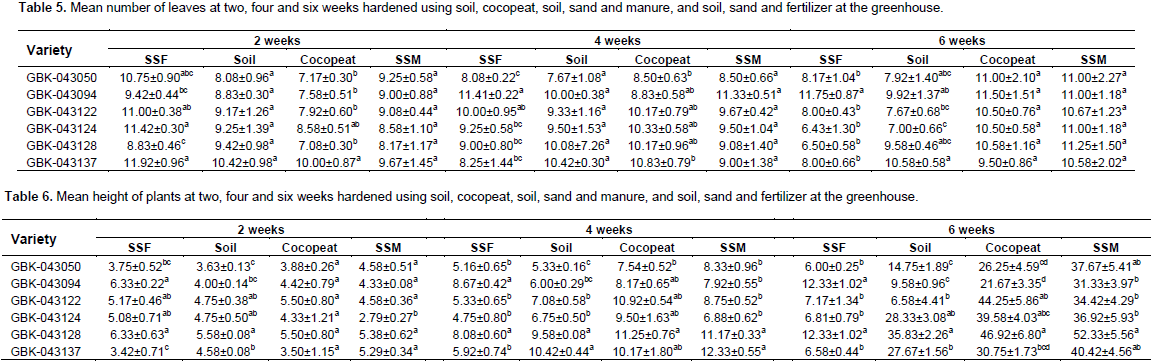
DISCUSSION
In this study, shoot apical meristems derived from mature seeds of several finger millet varieties germinated in culture used for the successful induction of embryogenic calli and subsequent plant regeneration. Although, six Kenyan farmers preferred finger millet varieties were used, this regeneration system can be extended into a range of agronomically important African finger millet varieties. Explants derived from mature seeds are considered an excellent source material for biotechnological application due to easy storage and accessibility to large amounts of uniform quality explant material (Sudhakar et al., 2004). Shoot apical meristems have been used successfully in the regeneration systems in cereals as starting material to obtain stable transformation in barley, wheat, maize, sorghum and millet (Sticklena and Orabya, 2005). The use of shoot apex in regeneration is critical since it can divide to produce viable new organs such as leaves, stems and adventitious roots (Itoh et al., 2006). Shoot apex is also most beneficial for its quick development therefore it can allow rapid development of plants (Ceasar and Ignacimuthu, 2008; Dey et al., 2012). Auxins have been shown to play an important role in inducing callus (Anjaneyulu et al., 2011). Therefore, in the current study, it was used alone or in combination with cytokinins. It was observed that by increasing levels of NAA across varieties resulted in the formation of callus-like structures at the base of the tip of explant then it started elongating and eventually shoots were formed which was higher as compared to other plant growth regulators. This trend is consistent with the works of Ceasar and Ignacimuthu (2008) who observed that after 5 weeks, NAA induced callus remained non-responsive. The previous reports on cereals tissue culture dealt with different auxins and cytokinins at different concentrations also showed the superiority of 2,4-D over the other auxins. The presence of NAA growth regulator was also found to be inhibitory for plant regeneration as well as callus proliferation. Medium supplemented with NAA was the worst performing media with 4.33±0.33 to 10.00±0.58 callus formation (Table 2a). It is therefore not understood why NAA performed poorly on the finger millet varieties tested. More studies should be done to investigate this callus proliferation as well as plant regeneration inhibitory performance.
Plant growth regulators auxins alone and cytokinins or in combination plays a very important role inducing callus and its proliferation (Thomas and Maseena, 2006). The degree of embryonic callus was improved by a combination of 2,4-D and BAP at 2.5 mg/l 2,4-D +0.5 mg/l of BAP for GBK-043137, GBK-043128, GBK-043122 and GBK-043050. A higher level of 3.0 mg/L 2,4-D + 1.5 mg/L was significant for GBK-043124 and GBK-043094. This is in agreement with the works of Anjaneyulu et al. (2011) who used 2 mg/l ,4-D + 1.0 mg/l of BAP obtaininghighest callus formation. Growth regulators at varying concentrations cause variability in plantlet regeneration. The callus obtained on both varieties across the hormones at earlier stages was white and compact but as it approached about six weeks, some were yellow and brown watery and soft. This agrees with the works of Yemets et al. (2013) who obtained three types: (1) callus white, compact, well-structured callus; (2) yellow or lemon, less compact, globular callus; (3) yellow, watery, soft callus. They were able to identify that the first type of callus was superior and was produced only when NAA was added to the 2,4-D + KIN combinations. This research confirms that the superior callus can also be obtained by combination of 2,4-D and BAP which is variety independent. In all the cases, use of MS alone which was used as control turned brown then died off without producing any callus. Two step embryo maturation containing cytokinin has proven to be effective for plantlet regeneration (Xie and Hong, 2001; Seabrook and Douglass, 2001). This was done using BAP and it was found out that the optimum for both varieties was at 1.75 mg/l. This is similar to the works of Patil et al. (2009) who found out that the best performing concentration was 2.0 mg/l and higher concentrations than this had negative effect on shoot induction. A combination of both cytokinins and auxins was used for root regeneration and it was observed that 1.0 KN + 0.25 NAA was significantly higher as compared to the rest of treatments for GBK-043137; this is in agreement with the works of Anjaneyulu et al. (2011) though the average number was lower in this study for varieties GBK-043094 1.0 BAP+0.25 NAA produced significantly higher shoots and it has the highest number. In both treatments, there was 100% root formation. Soil, sand and fertilizer regime were initially the best performing at two weeks in terms of leaf number and height because plants are able to acclimatize better but it was overtaken by cocopeat which did not retain water easily and at the first weeks, the plants showed signs of witling. However, with frequent watering, the plants improved significantly and at four weeks, they were the healthiest plants that displayed green colour and their size of stem was also the largest. The plants were acclimatized and hardened successfully with 97% survival rate. This high survival rates could be due to well-developed root system and the greenhouse conditions. The plants that died (80%) were those hardened on cocopeat; this could be due to the low capacity to retain water as compared to other media. Plants were regenerated after eight weeks of culture.
CONCLUSION
An effective, simple and reproducible system for somatic embryogenesis and regeneration of Kenyan finger millet varieties was developed using shoot apical meristems. This was achieved by combining auxin and cytokinin at appropriate concentrations and combinations to stimulate somatic embryogenesis from shoot explants and plant regeneration stages. The study has also proved that finger millet is recalcitrant to regeneration; its tissue culture is genotype dependent and also heavily influenced by environmental conditions under which the explants is grown. This reproducible protocol will form the basis for the improvement of this crop by transgenic technologies.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
We are grateful to Kenya Agricultural and Livestock Research Organization, Gene Bank, for providing the finger millet seed used. Support for this research was made possible through a capacity building competitive grant (RU/2016/DRRG/01/009) provided by Regional Universities Forum for Capacity Building in Agriculture (RUFORUM).
REFERENCES
|
Anjaneyulu E, Attitalla H, Hemalatha S, Eharath Raj S, Balaji M (2011). An efficient protocol for callus induction and plant regeneration in finger millet (Eleusine coracana L.) World Applied Sciences Journal, 12(7):919-923. |
|
|
Ceasar SA, Ignacimuthu S (2008). Efficient somatic embryogenesis and plant regeneration from shoot apex explants of different Indian genotypes of finger millet (Eleusine coracana (L.) Gaertn.). In Vitro Cellular & Developmental Biology-Plant, 44:427-435. |
|
|
Dey M, Bakshi S, Galiba G, Sahoo L, Panda SK (2012). Development of a genotype independent and transformation amenable regeneration system from shoot apex in rice (Oryza sativa spp. indica) using TDZ. 3 Biotechnology, 2:233-240. |
|
|
Gimode D, Odeny DA, de Villiers EP, Wanyonyi S, Dida MM, Mneney EE, Muchugi A, Machuka J, de Villiers SM 2016). Identification of SNP and SSR markers in finger millet using next generation sequencing technologies. PLoS one, 11(7): 1-21. |
|
|
Gupta SM, Arora S, Mirza N, Pande A, Lata C, Puranik S, Kumar J, Kumar A (2017). Finger millet: a "certain" crop for an "uncertain" future and a solution to food insecurity and hidden hunger under stressful environments. Frontiers in Plant Science, 8(643):1-14. |
|
|
Itoh JI, Sato Y, Nagato Y, Matsuoka M (2006). Formation, maintenance and function of the shoot apical meristem in rice. Plant Molecular Biology, 60:827-842. |
|
|
Miah G, Rafii MY, Ismail MR, Puteh AB, Rahim HA, Asfaliza R, Latif MA (2013). Blast resistance in rice: a review of conventional breeding to molecular approaches. Molecular Biology Reports, 40(3): 2369-2388. |
|
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, 15:473-497. |
|
|
Oduori COA (2005). The importance and research status of finger millet in Africa. Paper Presented at The McKnight Foundation Collaborative Crop Research Program Workshop on Tef & Finger Millet: Comparative Genomics of the Chloridoid Cereals at the Biosciences for East and Central Africa (BECA) ILRI, Nairobi, Kenya, 28 – 30 June 2005. |
|
|
Patil SM, Sawardekar SV, Bhave SG, Sawant SS, Jambhale ND, Gokhale NB (2009). Development of somaclones and their genetic diversity analysis through RAPD in finger millet (Eleusine coracana L. Gaertn.). Indian Journal of Genetics and Plant Breeding, 69: 132-139. |
|
|
Seabrook JEA, Douglass LK (2001). Somatic embryogenesis on various potato tissues from a range of genotypes and ploidy levels. Plant Cell Reports, 20:175-182. |
|
|
Sood S, Kumar A, Babu BK, Gaur VS, Pandey D, Kant L, Pattnayak A (2016). Gene discovery and advances in finger millet [Eleusine coracana (L.) Gaertn.] Genomics-An important nutri-cereal of future. Frontiers in Plant Science, 7(1634):1-17. |
|
|
Sticklena MB, Orabya HF (2005). Shoot apical meristem: a sustainable explant for genetic transformation of cereal crops. In Vitro Cellular and Developmental Biology-Plant, 41:187-200. |
|
|
Sudhakar D, Duc LT, BuiBa-Bong BB, Tinjuangjun P, Maqbool SB, Valdez M, Jefferson R, Christou P (2004). An efficient rice transformation system utilizing mature seed-derived explants and a portable, inexpensive particle bombardment device. Transgenic Research, 1:289-294. |
|
|
Thomas TD, Maseena EA (2006). Callus induction and plant regeneration in Cardiospermum halicacabum (L.) an important medicinal plant. Scientia Horticulturae. 108:332-336. |
|
|
Veluthambi K, Gupta K, Sharma A (2003). The current status of plant transformation technologies. Current Science, 84: 368-380. |
|
|
Xie DY, Hong Y (2001). Regeneration of Acacia mangium through somatic embryogenesis. Plant Cell Reports, 20:34-40. |
|
|
Yemets AI, Bayer GA, Blume YB (2013). An effective procedure for in vitro culture of Eleusine coracana (L.) Gaertn. and its application. ISRN Botany, 21: 503-510. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


