Full Length Research Paper

ABSTRACT
Alternanthera brasiliana is a medicinal plant with several popular use and high production of betalains. The aim of the present research was to establish a protocol for callus induction and betalain biosynthesis in A. brasiliana species, as well as to investigate the effect of different qualities of light in increasing betalain in A. brasiliana callus cultures. In this way, the callus initiation and the improvement of secondary metabolites were induced using varying phytohormones concentrations and under the influence of red, white and blue lights. To start the experiments, the plants were cultivated for 30 days in Murashige and Skoog (MS) medium to grow and form internodes. These were inoculated in MS medium supplemented with different combinations of plant growth regulators to find the best combination for medium callus induction (MCI). After another 30 days, the explants were transferred to a medium for betacyanin induction (MBI) containing thidiazuron (TDZ) and α-naphthalene acetic acid (NAA) under blue, white, and red lights. The best medium for A. brasiliana callus induction with higher production of betalains, was the MS medium supplemented white indole-3-acetic acid (IAA) and 2,4-dichlorophenoxyacetic acid (2,4-D), before inoculation on the MBI. The blue and white lights promoted callus pigmentation (betalains), whereas the red light was not effective at inducing pigmentation in the calli.
Key words: Amaranthaceae, betacyanin, betalains, pigments, spectra of light, secondary metabolites
INTRODUCTION
Seeking an improvement in health and quality of life in recent years, the use of synthetic dyes have been sidetracked, due to the potential hazards they generate, such as allergies and induction of various diseases, that way, natural dyes have gained market due to their bioactive properties and applications as health promoters when included in diets (Martins et al., 2017). Carotenoids, anthocyanin, and betalains are examples of plants withpigment production; physiologically, this production acts as attractants for pollinators and agents to disperse the seeds (Osbourn, 2017). These pigments have great potential and can be used as food dyes because they present not only coloring attributes, but also properties related to antioxidant, antiradical, anti-inflammatory and cytotoxic, radio protective, neuroprotective, antimicrobial, anti-proliferative and hepato-protective properties, enabling his use in food, cosmetic, and pharmaceutical industries (Georgiev et al., 2008; Georgiev et al., 2010; Biswas et al., 2013; Martins et al., 2017). Betalains are nitrogenated, water-soluble vacuolar pigments, produced by most of caryophyllales plants and never co-occur with anthocyanins. They are synthesized from L-tyrosine and L-phenylalanine (Osbourn, 2017)and are immonium derivatives of betalamic acid for which the conjugation system 1,7-diazaheptamethine is the chromophore. The carboxylic groups in the betalain structure provide the acidic characteristic to the molecule, thus, making it different from the alkaloid group, which has basic character (Herbach et al., 2006). The yellow-to-orange-colored betaxanthins release green auto fluorescence and are produced by condensation of betalamic acid and amino acids (or amines). Similarly, the violet betacyanins are O-glycosides (C-5 or C-6) and come from the spontaneous condensation of betalamic acid and cyclo-dopa.
The acylation of betacyanins generates its main derivative, betanin (betanidin-5-O-β-glucoside) (Gandía-Herrero et al., 2010; Reis et al., 2015). The great interest in the market of renewable natural products has drawn attention to the technical and commercial viability of a variety of systems, exploring in vitro and cell and tissue cultured plants as potential bio-factories of phytochemical products, where there is a potential in making the production more reliable, simple and predictable. The ability of plant cells, calli, and tissues cultivated in vitro to produce and accumulate chemicals is important by maintaining carefully a controlled and aseptic environment; these types of cultures can also provide an excellent source for in-depth research of metabolic and biochemical pathway and the possibility to predict how amount can be produced in the cultures during a period of time, something impossible to make in nature (Rao et al., 2002; Namdeo, 2007; Karuppusamy, 2009). Another important point is the use of elicitors (that is, chemical and stressor agents) during cultivation which can change or induce some metabolic pathways that affect, both qualitatively and quantitatively, the plant secondary metabolism (Karuppusamy, 2009; Othman et al., 2016). Physical stimulation using light is one of the elicitor agents widely used in plant tissue culture; due to the phytochromes and cryptochromes, plants can perceive a small fraction of light radiation, changing the gene expression and physiological responses and modifying and even maximizing plant development and levels of secondary metabolites as a form of protection (Katerova et al., 2017).
Red and blue lights have been used to elicit several phenotypic expressions via perception and signal transduction pathways. In the induction of betalain biosynthesis, the use of light was reported in sugar beet (Beta vulgaris), seedlings of Amaranthus caudatus, callus of Portulaca, cell cultures of Chenopodium album, and callus of Alternanthera brasiliana (L.) Kuntze and Adachi 1995; Macedo et al., 1999; Silva et al., 2005; Zhao et al., 2010). A. brasiliana (L.) Kuntz (Amaranthaceae) is a plant broadly distributed in South America, with ability to accumulate pigments as betalains (betacyanins and betaxanthins) and flavonoids. This species is used in traditional medicine for the treatment of several human pathologies as cough, diarrhea, infections, and it has also analgesic activity. Moreover, its activity as an antiproliferative, anti-inflammatory, antiedematous and antioxidant, has been reported. It also shows activity against the herpes simplex virus and, recently, anticonvulsant effect (Lagrota et al.,1994; Macedo et al., 1999, 2011; Facundo et al., 2012; Andreazza et al., 2013; Schallenberger et al., 2017). Based on the foregoing, A. brasiliana offers a promising object for plant biotechnology studies. Thus, the aim of the present research was to establish a protocol for callus induction and betalain biosynthesis in A. brasiliana species, as well as to investigate the effect of different qualities of light in increasing betalain in A. brasiliana callus cultures.
MATERIALS AND METHODS
Callus induction
The experimental A. brasiliana plants were grown for 30 days in a growth chamber under a photon flux density of 22 mmol m-2 s-1 for a 16-h photoperiod at 25 ± 2°C. Internode segments of 0.1 to 0.2 cm in length were placed in a medium for callus induction (MCI) (Zhao et al., 2010), which consisted of MS basal medium with sucrose 30 g L-1, phytagel 2 g L-1, myo-inositol 100 mg L-1, adenine 0.5 mg L-1, ascorbic acid 3 mg L-1, and different plant growth regulators. Three different combinations of phytoregulators were added: 1 mg L-1 kinetin (KIN) and 1 mg L-1 2,4-dichlorophenoxyacetic acid (2,4-D; MCI-1); 0.75 mg L-1 indole-3-acetic acid (IAA) and 1 mg L-1 2,4-D (MCI-2); and 1 mg L-1 6-benzylaminopurine (BA) and 0.5 mg L-1 2,4-D (MCI-3). Assembled six plates, each containing ten explants, were inoculated and placed in a growth chamber at 25 ± 2°C for 22 days in the dark, passed this time, and were transferred to white light for an additional 7 days.
Betacyanin induction and light sources
All explants from the MCI media described earlier were transferred to a medium for betacyanin induction (MBI) (Zhao et al., 2010), composed of MS medium supplemented with 3 g L-1 phytagel, 30 g L-1 sucrose, 100 mg L-1 myo-inositol, 0.5 mg L-1 adenine, 3 mg L-1 ascorbic acid, thidiazuron (TDZ) 0.5 mg L-1, and 1 mg L-1 of 1-naphthaleneacetic acid (NAA). Over a 40-day period, the plates containing the explants were placed under different light regimes: white light (full-spectrum-fluorescent tube Sylvania®; 40 W), blue light (peak emission at 470 nm-14 W, compact fluorescent lamp Taschibra®), and red light (peak emission at 660 nm-15 W, compact fluorescent lamp G-light®). The photon flow density for the white, blue, and red-light levels were 25, 12, and 22 mmol m-2 s-1, respectively, as measured using a light meter (Hansatech® Quantum Sensor QSRED).
Evaluation of callus pigment appearance and callus pigment intensity
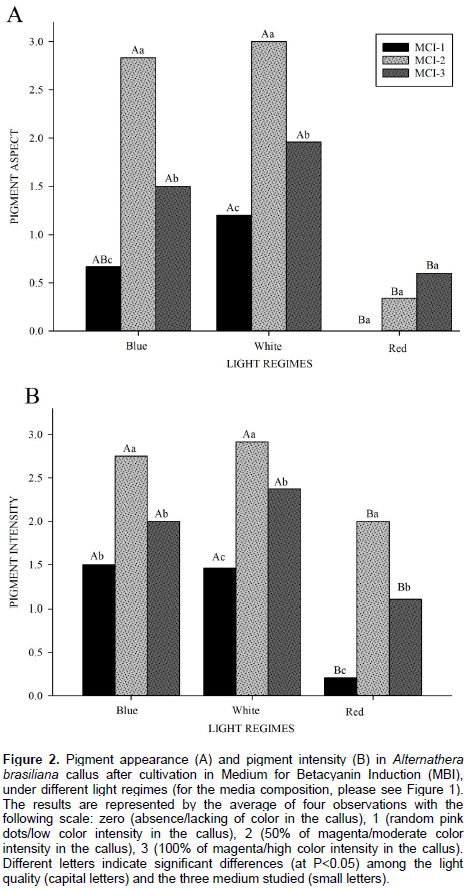
This evaluation was developed seeking a visual quantification of the appearance and intensity of the magenta pigmentation produced in the callus. Numerical scores were given to pigment appearance and intensity in the following manner: 0 (zero), for callus without pigmentation (absence of magenta color); (1) for random magenta dots in callus/low pigment intensity; (2) for approximately 50% of magenta in callus/moderate color intensity, and (3) for nearly 100% magenta in callus/high intensity of coloration visually detected. The scores of 0 to 3 were assigned by four different observers realized in the end of the experiment, after almost 70 days in culture, and the averages of the analysis were used.
Relative callus growth rate (RCGR) and total calli weight (Wc)
RCGR was calculated using the fresh callus weight (FC), as described by Silva and Dobránszki (2015), according to the following equation:
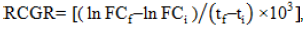
Where, FCf=final fresh callus weight, FCi=initial fresh callus weight, ti=first day of treatment, and tf=last day of treatment. The total calli weight (Wc) was calculated at the end of the experiment based on the fresh callus weight (FC) from a Petri dish containing 10 calli.
Betaxanthin quantification
Homogenates using 0.25 g of fresh calli, produced by induction, were obtained by maceration in a porcelain mortar using phosphate buffer at 10 mM (pH 6.0) and 10 mM sodium ascorbate. The homogenates were filtered through cheesecloth and centrifuged at 10000 g, for 20 min at 4°C (Gandía-Herrero et al., 2005). The spectrophotometric analyses were performed on a T80 UV/VIS Spectrometer (PG Instruments) kept at 25°C (PTC-2 Peltier Temperature Controller). The betaxanthin concentration calculated using the molar extinction coefficient of miraxanthin (e=48000 M-1cm-1) after measurements was taken at 480 nm (Schliemann et al., 1999).
Total betacyanins quantification
The total amount of betacyanins corresponds to the sum of betanidin and betanin amounts, being the first aglycone and the second glycosylated. To extract betanidin, an acetate buffer at 10 mM and methanol (70:30; v/ v) at a pH 5.0, plus sodium ascorbate of 10 mM were used. To obtain betanin, a phosphate buffer of 10 mM at pH 6.0, plus sodium ascorbate (10 mM) and organic solvents was used. For both extractions, 0.25 g of fresh calli was weighed, and the homogenates were produced as described for the betaxanthin quantification. The molar extinction coefficients e=54000 M-1cm-1 and e=65000 M-1cm-1 were employed to calculate the amount of betanidin and betanin, respectively. A wavelength of 536 nm was used as described previously (Gandía-Herrero et al., 2005).
Total flavonoids quantification
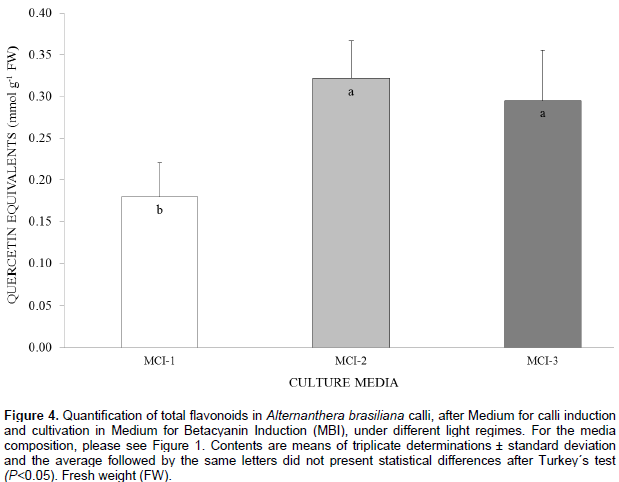
The quantification of total flavonoids was done as described by Salvador et al. (2006), using an acetate/methanol buffer as the extraction solvent and the measurements were done at wavelength of 330 nm. The results were expressed as the amount of quercetin (mmol) per g of calli, using the linear equation: y = 0.1548x - 0.005 (r2 = 0.9952).
Normalized index of variation (NIV)
NIV was calculated using the equation (Tattini et al., 2006):
NIV = (X Light - X Control) (X Light + X Control),
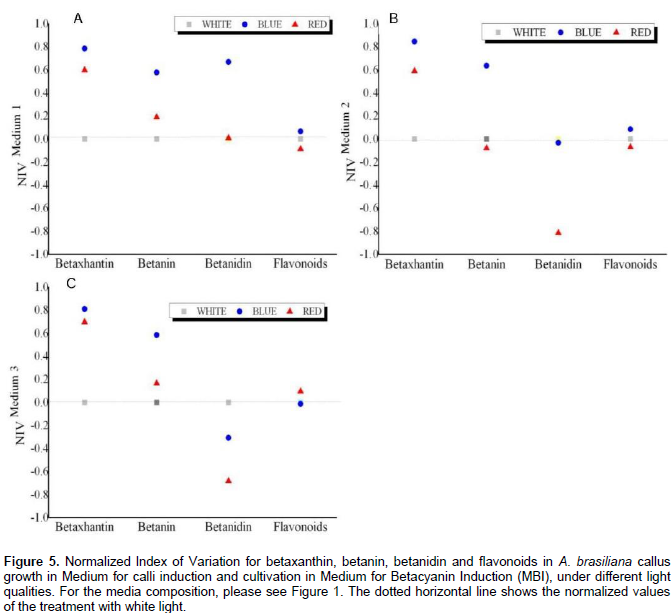
Where, Xlight is the two tested light colors (blue and red) and XControl is the white light. The calculations were performed separately for each type of medium used. The NIV depicts the actual treatment effect of the analyzed calli and it may be positive or negative as compared to the control (white light).
Statistical analysis
The experiment was completely randomized in a factorial 3 × 3 design, represented by the three different media cultures and the three light levels. A total of six replications (6 Petri dishes) were performed, with experimental unit being a Petri dish with ten explants. The assays were performed three times and the statistical analyses, ANOVA and Tukey’s tests were performed at P <0.05 to calculate significant differences in treatments, using the statistical software SAS v.9.3 (SAS Institute Inc., Cary, NC) (SAS, 2003).
RESULTS
Evaluation of callus pigment appearance and callus pigment intensity
The first changes in coloration of the calli were observed at the beginning of the experiment. Coloration intensified when the samples were placed under the different lights. The calli cultivated in the darkness were white in color and after exposure to different lights turned green (Figure 1G and H), pink (Figure 1E), or magenta (Figure 1B), or de mixture of this colors, green with small magenta dots on some calluses (Figure 1I), green with the center becoming magenta (Figure 1A, C and D) and the inverse, and most magenta/pink with some clear green parts (Figure 1F). Pigmentation changes were not found along the entire epidermal surface, but only in specific regions, thus forming these mosaics of colors. The calli grown for 40 days in the MBI composed of white, green, and magenta spherical-cell agglomerates, and an opaque color (clear green) was also observed in the calli surface (Figure 1F). Under red light, all the experimental plates, with the different combination of phytoregulators showed oxidation of the calli tissue. As the only source of photosynthesis and a vital environmental factor, light plays an important role in directing photosynthetic biosynthesis and photo-morphogenesis, which are correlated with the phytochemical biosynthesis and accumulation (Bian et al., 2014). Although, in the experiment, all plants were clones, same growth age and initial culture medium, in the first phase of callogenesis induction, different growth regulators were used, which caused the formation of tissues with their inherent characteristics and, allowing different photomorphogenesis and demonstrating these characteristics phenotypically and metabolically by the diverse colorations presented in the callus.

The light quality (spectrum) has the potential to be involved in the shades of betalains seen in A. brasiliana calli, where it promoted a great impact on the morphogenesis and metabolism of these structures. The apparent colors of the calli depended on the level of light exposure for the three media combinations used. The explants cultivated in blue and white lights provided a higher number of calli with pigmented dots, partially and fully magenta. These qualities of light were found as the most visually pigmented treatment, in MCI-2 (Figure 1B and E, for blue and white light, respectively), where the calli were 100% magenta and associated with the presence of betalains. In the red light, however, there was no formation of apparent pigments, barely small spots in some callus (less than 5% magenta) in the MCI-3 (Figure 1I). The evaluation of color intensity showed the same pattern for color appearance: the best lights for production of pigments are moderate and high intensity white and blue lights. The calli grown in MCI-2 had the highest intensity of magenta color for all light regimes, with moderate to intense coloration being displayed (Figure 2B). The statistical analysis corresponds to the score values as shown in Figure 2.
RCGR and Wc
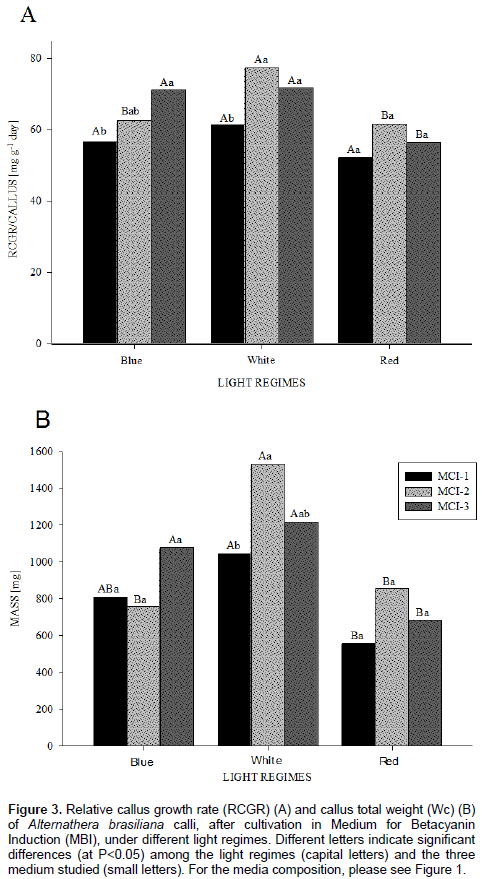
The effects of light regime are felt throughout the process of plant morphogenesis. Thus, through the analysis of RCGR and the Wc, the differences in the growth pattern were clearly visualized in the different light qualities tested. RCGR in blue and white light exposure the outcomes stood out from the results in red light (Figure 3A). For calli grown on MCI-1, their RCGR did not differ among the three light levels. When grown on MCI-2, the cultivation under white light promoted better growth results in these cells formations. The RCGR in MCI-3 was better influenced by white and blue lights, with similar results, found for a daily increase in calli cell mass in these two light qualities. About the different medium in the same light, under white light, the RCGR had better results for MIC-1 and 2; MIC-3 presented the best results in blue light, although, did not differ statistically from the MCI-2 medium cultured in this quality of light. Under red light, however, significant differences were not detected among the different media for calli cultivation. Corresponding to the Wc, the greatest mass accumulation was visualized in white light in all combinations of growth regulators tested (Figure 3B). The callus cultivated in MCI-1 showed noteworthy results in white light, but did not differ statistically from the blue light. In the cultures from MCI-2, the white light provided a greater increase in the total cellular mass and those cultured in the MCI-3 medium, had a rise of the mass in the white and blue lights, not statistically different in both. Concerning the correlation between the formation of cell agglomerates and the light quality, under blue and red lights, the final mass is not different among the media, but for the media cultivated in white light, both MCI-2 and 3 yielded calli with a higher final biomass accumulation.
Betaxanthins, betacyanins and total flavonoids quantification
To investigate the additional effects of the light qualities of white, blue, and red levels in the accumulation of secondary metabolites in the A. brasiliana calli, the samples were subjected to a quantification analysis of their betalain pigments and of their total flavonoids (both after 40 days of cultivation in MBI). The spectrophotometric analysis revealed that the blue light was more effective in the induction of betaxanthin biosynthesis (Table 1 and Figure 5A, B, and C) in all the media assayed, with the concentration of this pigment peaking in MCI-2. Under irradiation by red light, however, both MCI-2 and MCI-3 induced a higher accumulation of betaxanthins, whereas the white light did not show any significant differences among the treatments. It shows the results for betanin, which had the highest yield in the calli extracts grown under blue light and in both MCI-2 and MCI-3 (Table 1). The MCI-1 was an exception for blue light and did not show significant results in the production of this glycosylated betacyanins, similar to the treatment using white light, which was not statistically significant in the media culture used. The results of the quantification of betanidin in calli are presented in Table 1. The blue light was inductor for this type of pigment production in the MCI-2 and MCI-3. Under blue light, pigment production was higher in the calli cultivated in MCI-2, medium composed by two auxins. For the calli subjected to white light, calli inducted in MCI-2 and MCI-3 had higher biosynthetic production of aglycones. MCI-1 and red-light treatments did not present statistical differences in the treatments.
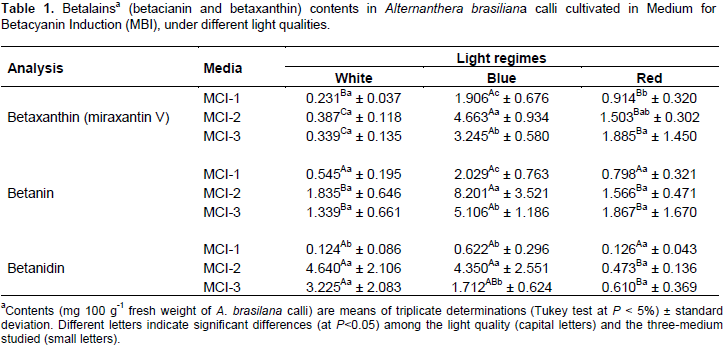
In the quantification of total flavonoids (Figure 4), all the culture media treatments produced a significant effect independently of the light quality to which it was submitted. The media containing only auxins (MCI-2) and cytokinin combined to auxin (MCI-3), presented the best results as inductors for the biosynthesis of flavonoids in A. brasiliana calli.The estimated Normalized Index of Variation (NIV) helps to visualize the relative effects of the red and blue qualities of light in different culture media. In MCI-1, betaxanthin, betanin, and betanidin indices were higher (0.78, 0.58 and 0.66, respectively) when treated with the blue light rather than that by red light (the latter had values closer to the control, white light). The NIV values for flavonoids in different light qualities did not change compared to those in the control (Figure 5A). In MCI-2, there is an increase in NIV values for the content of betaxanthin (0.84) and betanin (0.63) under the treatment of blue light. However, neither the blue or red lights increased the content of flavonoids and betanidin (Figure 5B). In MCI-3, the NIV-related content of betaxanthin and betanin enhance under blue light(0.81 and 0.58, respectively) but not so for contents of flavonoid (-0.013) and betanidin (-0.30). For the last variable, however, both light qualities of blue and red had a negative effect compared to the control (Figure 5B and C).
DISCUSSION
The quality of the light spectrum and the culture medium can influence the morphogenesis process from the entire plant to those grown in vitro. Species like Camptotheca acuminata had their seedlings studied with different qualities of light demonstrating that the development, leaf area, chloroplast development and photosynthetic efficiency were modified by the use of red light rather than blue or yellow light (Yu et al., 2017). In strawberry (Fragaria × ananassa), the total content of anthocyanins increased by the use of red and yellow plastic films, suggesting the activation of enzymes or transcription factors related to the flavonoid pathway (Miao et al., 2016). As reported in the methods, the period of darkness at the beginning of calli induction were required for the formation of the cell masses and the reduction of phenolic compounds, which can affect the explants; this was specified by Tan et al. (2010). The changes in the calli pigments and its intensity were visualized in the first week after exposing the plates in an environment treated by different kinds of light (visual observations, data not shown), firstly in white light. Reports have demonstrated that betalains biosynthesis occurs between the sixth and fifteenth days, but is most intense towards the ninth day (Radfar et al., 2012). The development of different lineages with colored cells in plant tissue cultures depends on specific gene sequences in the calli induction and these individual phenotypes, once established, can be perpetuated and maintained in the culture medium (Girod and Zyrd, 1991). A determining factor for the establishment and stability of these lineages is the composition of the culture medium for A. brasiliana calli. The MS medium containing IAA (0.75 mg L-1) and 2,4-D (1 mg L-1) exposed to blue and white lights have been highlighted as the strongest inductors for production and intensification of the pigments. In studies that used Zaleya decandra calli grown under white light (Radfar et al., 2012), the highest pigments intensity was seen when the samples were cultured on the MS-plus-TDZ (2 mg L-1) and 2,4-D (1 mg L-1). However, it is not entirely clear whether the intense pigmentation is due to an increase of metabolic activity in individual cells, or simply an increase in the number of cells able to produce betalains (Kishima et al., 1991).
Changes in the combinations of the plant growth regulators in a culture medium can be used to modulate the frequency and direction of the inter-conversion events that result in chimeric phenotypes, which are calli with only one-color pattern and after to transferring them to a medium with low concentration of 2,4-D, sectors with another coloration arise (Leathers et al., 1992). The speed of this response, which may be 1-2 cell generations, indicates that the transformation of the phenotype, induced by hormones, it is associated with cellular DNA replication and therefore susceptible to a cell proliferation process (Girod and Zryd, 1991). In other words, depending on the effect induced by the phytohormones, a phenotype can change the color and shape of the cells made of it. The RCGR and Wc of A. brasiliana callus showed the best results under white and blue lights, agreeing with results reported for another species as Suaeda salsa (Zhao et al., 2010), which had a RCGR markedly higher in calli grown under white light. The calli grown on medium containing IAA and 2,4-D (MCI- 2) and medium with BA and 2,4-D (MCI-3) had higher rates of daily growth, as well as higher final mass accumulations. Studies done with Portulaca species calli (Noda and Adachi, 2000)showed the highest growth rate in calli cultured on a MS medium supplemented with 5 to 10 mM of 2,4-D, with identical growth rates in different concentrations of auxin. Also, calli growth is higher when auxins are incorporated in the medium than when compared with that supplemented by cytokinins (Lee et al., 2011; Abu-romman and Suwwan, 2013). The accumulation of betacyanins when the plates were submitted to cultivation under darkness indicates that the light exposure is not a prerequisite for the formation of betacyanins in some plant species, but rather a powerful stimulant for their biosynthesis (Leathers et al., 1992).
This light-induced mechanism of biosynthesis begins when the light signal from the phytochrome or cryptochrome passes through multiple signaling intermediates, which then regulates a transcription factor that can control the expression of genes encoding key enzymes such as tyrosinase, DOPA oxidase, and glycosyl transferases, thereby triggering their post-translational modification, a process of fundamental importance in the formation of this pigment (Zhao et al., 2010). Exposure of Amaranthus tricolor (Elliott, 1979)and Celosia plumose (Nicola et al., 1974)seedlings to red-and white-light lamps increased their betacyanins and betaxanthins production. In A. brasiliana, the highest biosynthesis of betaxanthins and glycosylated betacyanins was visualized in MCI-2 medium after exposure to blue light. These results corresponded to an approximately 12- and 4.5-fold increase in productivity when compared with the white light that is commonly used in tissue culture. Similar results were reported for cell suspensions of Chenopodium rubrum, for which an increase of 30% in amaranthin levels and 10% of betanin when cultivated under blue light (Berlin et al., 1986). Studies about the synergistic effect between kinetin and light in A. tricolor seedlings (Bianco-Colomas and Hugues, 1990)showed extremely positive results, with a high accumulation of betacyanins. By contrast, the results showed that the highest betacyanin concentration arose from betacyanins aglycones in a medium composed of auxin; these results showed that exposure to blue and white lights induced approximately 10 times more aglycones than under red light. It seems, therefore, that the pigmentation response generated by the luminous environment is an intrinsic feature of the cells themselves and related to the species to which they belong (Kishima et al., 1991).
The quantification of total flavonoids revealed higher amounts in the MCI-2 and 3, agreeing with results obtained using Morus alba in which the auxins strongly induced the production of flavonoids in calli from adventitious roots cultured on MS medium supplemented with 5 mg L-1 of IAA (Lee et al., 2011). Interestingly, in studies using cell cultures of Centella asiatica, adding 2,4-D and kinetin to the culture media stimulated the production of flavonoids as quercetin, kaempferol, luteolin and rutin (Tan et al., 2010). It is evident then, in a general picture, that the MCI-2 promoted the highest biosynthesis of metabolites, regardless of the level in light quality, when composed of a natural and a synthetic auxin (IAA, 0.75 mg L-1 and 2,4-D, 1 mg L-1), respectively. Both auxins play an important role in the induction of calli and various auxins can have different effects, although the synthetic auxins in many cases are more effective than the natural ones (Baskaran et al., 2014). According to the results, the best medium for calli with a greater amount of mass and RCGR is the MCI-2, supplemented with both IAA and 2,4-D before inoculation on the MBI and cultivation in white light was ideal for A. brasiliana callus. The MCI-2 was the best also for the production of the metabolites of interest in this study; so, for production of quercetin-type flavonoids, white light has an essential importance. In the production of betacyanin aglycone- types, the white and blue qualities of lights are both inductive of these pigments. However, for the biosynthesis of a greater amount of betacyanin glycosylated-types and for betaxanthins in A. brasiliana calli, it is indicated that the induction phase of the betalains is carried out under blue light, reaching a greater quantity of pigments in this quality of light. In these experiments, the red light was ineffective at inducing pigmentation in the calli of this plant species.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This study was supported by the Brazilian research funding agencies: Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação de Amparo à Pesquisa do Rio Grande do Sul (FAPERGS).
REFERENCES
|
Abu-Romman S, Suwwan M (2013). The influence of plant growth regulators on callus induction from hypocotyls of cucumber (Cucumis sativus L.). Adv Environ Biol. 5:339-344. |
|
|
Andreazza LN, Lourenço C, Siqueira ATC, Sawaya AF, Lapinski T, Gasparetto A, Salvador JM (2013). Photodynamic inactivation of yeast and bacteria by extracts of Alternanthera brasiliana. Curr. Drug Targets. 15:1015-1022. |
|
|
Baskaran P, Moyo M, Van-Staden J (2014). In vitro plant regeneration, phenolic compound production and pharmacological activities of Coleonema pulchellum. South Afr. J. Bot. 90:74-79. |
|
|
Berlin J, Sieg S, Strack D, Bokern M, Harms H (1986). Production of betalains by suspension cultures of Chenopodium rubrum L. Plant Cell Tissue Organ Cult. 5:163-174. |
|
|
Bian ZH, Yang QC, Liu WK (2014). Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 95:869-877. |
|
|
Bianco-Colomas J, Hugues M (1990). Establishment and characterization of a betacyanin producing cell line of Amaranthus tricolor: inductive effects of light and cytokinin. Plant Physiol. 136:734-739. |
|
|
Biswas M, Das SS, Dey S (2013). Establishment of a stable Amaranthus tricolor callus line for production of food colorant. Food Sci. Biotechnol. 22:1-8. |
|
|
Chen JR, Wu L, Hu BW, Yi X, Liu R, Deng RN, Xiong XY (2014).The influence of plant growth regulators and light quality on somatic embryogenesis in China rose (Rosa chinensis Jacq.). J. Plant Growth Regul. 33:295-304. |
|
|
Elliott DC (1979). Ionic regulation for cytokinin-dependent betacyanin synthesis in Amaranthus seedlings. Plant physiol. 63:264-268. |
|
|
Facundo VA, Azevedo MS, Rodrigues RV, Nascimento LFD, Militão JS, da Silva GV, Braz-Filho R.(2012). Chemical constituents from three medicinal plants: Piper renitens, Siparuna guianensis and Alternanthera brasiliana. Braz. J. Pharmacogn. 22: 1134-1139. |
|
|
Gandía-Herrero F, Escribano J, García-Carmona F (2005). Betaxanthins as substrates for tyrosinase. An approach to the role of tyrosinase in the biosynthetic pathway of betalains. Plant physiol.138:421-432. |
|
|
Gandía-Herrero F, Escribano J, García-Carmona F (2010). Structural implications on color, fluorescence, and antiradical activity in betalains. Planta 232:449-460. |
|
|
Georgiev V. Ilieva M. Bley T. Pavlov A. (2008). Betalain production in plant in vitro systems. Acta Physiol Plant. 30:581-593. |
|
|
Georgiev VG, Weber J, Kneschke EM, Denev PN, Bley T, Pavlov AI (2010). Antioxidant activity and phenolic content of betalain extracts from intact plants and hairy root cultures of the red beetroot Beta vulgaris cv. Detroit dark red. Plant Foods Hum. Nutr. 65:105-111. |
|
|
Harris NN, Javellana J, Davies KM, Lewis DH, Jameson PE, Deroles SC, Schwinn KE (2012). Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA. BMC Plant Biol. 12:34. |
|
|
Herbach KM, Stintzing FC, Carle R (2006). Betalain stability and degradation—structural and chromatic aspects. J. Food Sci. 71:41-50. |
|
|
Karuppusamy SA (2009). Review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 3:1222-1239. |
|
|
Katerova Z, Todorova D, Sergiev I (2017). Plant secondary metabolites and some plant growth regulators elicited by UV irradiation, light and/or shade. Med. Plants Environ. Challenges. pp. 97-121. |
|
|
Kishima Y, Nozaki K, Akashi R, Adachi T (1991). Light-inducible pigmentation in Portulaca callus; selection of a high betalain producing cell line. Plant Cell Rep. 10:304-307. |
|
|
Kishima YSA, Adachi T (1995). Evidence that blue light induces betalain pigmentation in Portulaca callus. Plant Cell Tissue Organ Cult. 43:67-70. |
|
|
Lagrota MMC, Wigg MD, Santos MMG, Miranda MMFS, Camara FP, Conceiro JNSS, Costa SS (1994). Inhibitory activity of extracts of Alternanthera brasiliana (Amaranthaceae) against the Herpes Simplex Virus. Phytother. Res. 8:358-361. |
|
|
Leathers RR, Davin C, Zryd P (1992). Betalain producing cell cultures of Beta vulgaris L. var. bikores monogerm (red beet). In Vitro-Plant. 28:39-45. |
|
|
Lee Y, Lee DE, Lee HS, Kim SK, Lee WS, Kim SH, Kim MW (2011). Influence of auxins, cytokinins, and nitrogen on production of rutin from callus and adventitious roots of the white mulberry tree (Morus alba L.). Plant Cell Tissue Organ Cult. 105:9-19. |
|
|
Macedo AF, Barbosa NC, Esquibel MA, Souza MM, Cechinel-Filho V (1999). Pharmacological and phytochemical studies of callus culture extracts from Alternanthera brasiliana. Pharmazie. 54:776-7. |
|
|
Macedo AF, Leal-Costa MV, Tavares ES, Lage CLS, Esquibel MA (2011). The effect of light quality on leaf production and development of in vitro-cultured plants of Alternanthera brasiliana Kuntze. Environ. Exp. Bot. 70:43-50. |
|
|
Martins N, Roriz CL, Morales P, Barros L, Ferreira IC (2017). Coloring attributes of betalains: a key emphasis on stability and future applications. Food Funct. 8:1357-1372. |
|
|
Miao L, Zhang Y, Yang X, Xiao J, Zhang H, Zhang Z, Jiang G (2016). Colored light-quality selective plastic films affect anthocyanin content, enzyme activities, and the expression of flavonoid genes in strawberry (Fragaria× ananassa) fruit. Food Chem. 207:93-100. |
|
|
Namdeo AG (2007). Plant cell elicitation for production of secondary metabolites, Reviews Pharmacognos. 1:69-79. |
|
|
Nicola MGDe, Amico V, Piattelli M (1974). Effect of white and far-red light on betalain formation. Phytochemistry 13:439-442. |
|
|
Noda N, Adachi T (2000). Isolation of stable, variously colored callus lines in Portulaca sp. "Jewel" and analysis of Betalain composition. Plant Biotechnol. 17:55-60. |
|
|
Osbourn A (2017). Painting with betalains. Nat. Plants. 3:852-853. |
|
|
Othman A, Ismail A, Hassan FA, Yusof BN, Khatib A (2016). Comparative evaluation of nutritional compositions, antioxidant capacities, and phenolic compounds of red and green sessile joyweed (Alternanthera sessilis). J. Funct. Foods. 21:263-271. |
|
|
Radfar M, Sudarshana MS, Niranjan MH (2012). Betalains from stem callus cultures of Zaleya decandra L. N. Burm. f. - A medicinal herb. Res.J. Med. Plant. 6: 2443-2447. |
|
|
Rao SR, Ravishankar GA (2002). Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 20:.101-153. |
|
|
Reis A, Kleinowski AM, Klein FRS, Telles RT, do Amarante L, Braga EJB (2015). Light quality on the in vitro growth and production of pigments in the genus Alternanthera. J. Crop Sci. Biotechnol. 18:349-357. |
|
|
Salvador MJ, Ferreira EO, Mertens-Talcott SU, De Castro WV, Dias DA (2006). Isolation And HPLC Quantitative Analysis Of Antioxidant Flavonoids from Alternanthera tenella Colla. Zeitschrift fur Naturforsch. 61:19-25. |
|
|
SAS- Statistical Analysis Software (2003). Statistical Analysis Software, Version 9.1.3. SAS, Institute Inc., Cary North Carolina, U.S.A. |
|
|
Schallenberger C, Vieira V, Krai JS, Suyenaga E, Tavares RG, Coitinho AS (2017). Anticonvulsant Effect of Alternanthera brasiliana Extract On Pentylenetetrazole-induced Seizures in Rats. J. Neurosci. Clin. Res.100:15-25. |
|
|
Silva NCB, Macedo AF, Lage CLS, Esquibel MA, Sato A (2005). Developmental effects of additional ultraviolet a radiation, growth regulators and tyrosine in Alternanthera brasiliana (L.) kuntze cultured in vitro. Braz. Arch. Biol. Technol. 48:779-786. |
|
|
Silva JAT, da Dobránszki J (2015). How do magnetic fields affect plants invitro? Vitr. Cell Dev. Biol-Plant.51:233-240. |
|
|
Tan SH, Musa R, Ariff A, Maziah M (2010). Effect of plant growth regulators on callus, cell suspension and cell line Selection for flavonoid production from pegaga (Centella asiatica L. Urban). Am. J. Biochem. Biotechnol. 6:284-299. |
|
|
Tariq U, Ali M, Abbasi BH (2014). Morphogenic and biochemical variations under different spectral lights in callus cultures of Artemisia absinthium L. J Photochem. Photobiol. B. Biol. 130:264-271. |
|
|
Tattini MD, Remorini D, Pinelli P, Agati G, Saracini E, Traversi ML, Massai R (2006). Morpho-anatomical, physiological and biochemical adjustments in response to root zone salinity stress and high solar radiation in two Mediterranean evergreen shrubs, Myrtus communis and Pistacia lentiscus. New Phytol. 170:779-794. |
|
|
Yu W, Liu Y, Song L, Jacobs DF, Du X, Ying Y, Wu J (2017). Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata seedlings. J. Plant Growth Regul. 36:148-160. |
|
|
Zhao SZ, Sun HZ, Chen M, Wang BS (2010). Light-regulated betacyanin accumulation in euhalophyte Suaeda salsa calli. Plant Cell Tissue Organ Cult. 102:99-107. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


