Full Length Research Paper

ABSTRACT
This study aimed to characterize and evaluate possible modifications in the leaf anatomy of crambe during the process of shoot induction based on micropropagation protocol. The anatomic characteristics of the leaves, and also the morphological characteristics of crambe plantlets, were evaluated during the shoot induction phase of the micropropagation. The shoots were induced by the cytokinins, 6-benzylaminopurine (BAP), kinetin (KIN) and thidiazuron (TDZ), at distinct concentrations on Murashige and Skoog (MS) medium during 120 days of culture. Comparing the leaf anatomy, it was observed that, at day 30, only the adaxial epidermis and the palisade parenchyma presented significant differences in relation to the tested concentrations, independently of the type of cytokinin employed. At 120 days, the anatomic analysis of the mesophyll demonstrated no marked difference among the cytokinins at 5 µM. Therefore, the various sources and concentrations of cytokinins applied in this work did not promote marked changes in the sense of altering the organization and/or thickness compared to the control.
Key words: Oilseeds, micropropagation, mesophyll, histology, plant morphology.
INTRODUCTION
Crambe abyssinica (Brassicaceae) is a species of this genus, which is most economically important, with 34 representatives (Wang et al., 2004). Its seeds contain 45 to 50% of crude protein and up to 35% of oil, which consists of up to 55 to 60% of erucic acid (Palmer and Keller, 2011). Such characteristics make it highly valuable for animal feed, industrial and pharmaceutical applications, biodiesel production, as well as fabrication of coatings, detergents, gliding products, cosmetics, lubricants and nylon (Vargas-Lopez et al., 1999; Capelle and Tittonel, 1999; Palmer and Keller, 2011).
The genetic breeding of species like C. abyssinica, with the aim of increasing their oil production or altering any compound of interest, has been often employed (Wang and Peng, 1998; Murphy, 1999; Wang et al., 2003; Wang et al., 2004; Li et al., 2010). However, in order for the genetic breeding to be successful, biotechnological steps beforehand must be efficiently concluded. For crambe, the lack of protocols suitable for micropropagation (in vitro culture) and the need for large amount of plant material have hampered the development of transgenic plants (Gao et al., 1998; Li et al., 2010).
In previously published protocols, the in vitro regeneration of crambe was achieved by using hypocotyls as explants to enable studies related to genetic breeding of the species, such as transgenesis and somatic hybridization (Wang et al., 2004, 2003; Wang and Peng, 1998). However, studies related to the induction of shoots and regeneration of whole plants from the apical segment have not been described yet. The in vitro environment offers low light density, high humidity, presence of sugar and growth regulators in the substrate, and low gas exchange, with the lack of CO2 and excess of ethylene (Kadlecek et al., 2001). These factors induce disruptions in the growth, development and morphology, involving anatomic and physiological alterations in the plant (Kozai, 1991).
The anatomic abnormalities may result from excess of growth regulators present in the culture medium (Hronkova et al., 2003). Only few studies exist regarding these characteristics during the process of shoot induction in the successive phases of micropropagation (Appezzato-Da-Glória et al., 1999; Apóstolo et al., 2005; Hazarika, 2006). There are no anatomic studies of C. abyssinica available in the specific literature yet. Hence, the present study aimed to characterize and evaluate possible modifications in the leaf anatomy of C. abyssinica and morphology of the plantlets during the process of induction of shoots obtained through micropropagation protocol.
MATERIALS AND METHODS
The experiment was carried out at the Laboratory of Plant Tissue Culture at the Center of Agronomic Sciences, and at the Laboratory of Plant Anatomy at the Department of Biological Sciences, both at the Federal University of Espírito Santo, ES – Brazil. Seeds of C. abyssinica Hochst cv. ‘FMS brilliant’, harvest of 2012, were used, supplied by the Fundação MS, located in Maracaju, Mato Grosso do Sul – Brazil.
The seeds of crambe were initially washed under running water with neutral detergent. Subsequently, they were transferred into aseptic conditions under laminar flow hood. Next, the seeds were disinfested by immersion in antibiotic solution containing penicillin (10 mg L-1) and rifampicin (10 mg L-1) for 30 min, followed by 70% alcohol for 1 min, commercially available 50% sodium hypochlorite (active chlorine: 2 to 2.5%) for 30 min, then washed three times in autoclaved distilled water. The used culture medium was half-strength MS medium (Murashige and Skoog, 1962), supplemented with 15 g L-1 sucrose and 7 g L-1 agar. pH of the media was adjusted to 5.8 prior to autoclaving under 1.1 atm. and 121°C, for 20 min. The seeds were inoculated into 25 × 150 mm test tubes containing 10 ml of medium, and incubated in growth room with photoperiod of 16/8 h (light/dark), under fluorescent lamps providing flow of 25.2 µmol m-2 s-1 photosynthetic photons flux’s, and temperature of 25 ± 1°C.
After 30 days, the apical segments of the obtained suitable plantlets were removed and used as explants in the process of induction and subculture of shoots. The shoots were induced by the cytokinins 6-benzylaminopurine (BAP), kinetin (KIN) and thidiazuron (TDZ) during 120 days of culture. The experiment followed a factorial scheme 3 × 3, having three levels in the factor cytokinin (BAP, KIN and TDZ), and three levels in the factor concentration (0, 5 and 10 µM), in a completely randomized design with five repetitions, each consisting of ten test tubes of 25 × 150 mm with one explant each and 10 ml of culture medium.
The adopted culture medium was MS, supplemented with 30 g L-1 sucrose, 0.5 mg L-1 silver nitrate (AgNO3) (LI et al., 2010) and 7.0 g L-1 agar, and pH fixed to 5.8. The tubes were autoclaved and stored under the growth room conditions already described. The number of shoots per explant (NSE) and percentage of survival (%SUR) were evaluated after 30, 60, 90 and 120 days. The removal of leaves per treatment for the anatomic analyses was done at 30 and 120 days.
Anatomic analysis
The anatomic analyses were performed in five more developed leaves for each treatment, in the midrib region and internervural area. The samples were fixed in formalin-acetic acid-alcohol (FAA50; Johansen, 1940) for 48 h, stored in ethyl alcohol 70%, later subjected to dehydration in increasing ethanol series (70, 90, 95 and 100%), and embedded in hydroxyethyl methacrylate (Leica®, Germany). The blocks were cross sectioned on a rotating microtome, with thickness varying between 8 and 10 µm. The sections were stained with 0.05% toluidine blue in acetate buffer, pH 4.3 (O'brien et al., 1964), and mounted in synthetic resin Entellan®. Measurements of the thickness of the blade, mesophyll, adaxial and abaxial epidermis, palisade parenchyma, spongy parenchyma and midrib region were performed. Also, the vascular tissue area of the midrib was measured. The observations and measurements were carried out under microscope Nikon E200 with coupled digital camera connected to a computer containing the software Tsview® (China). The photo documentation was done in a photomicroscope Nikon 50i (Nikon, Japan). The design for anatomic evaluations was completely randomized, with five repetitions per treatment, each repetition corresponding to one leaf and comprising the mean value of three slides with eight sections each.
Statistical analysis
All obtained data was subjected to analysis of variance, after verification of normality and homogeneity. After confirming the significance of the treatments, the Tukey test of averages was applied, adopting 1 and 5% probability, with the software Assistat (Silva and Azevedo, 2009). The regression analyses in function of the cytokinins and the evaluation times were performed with the software SigmaPlot 10.0 (2006). However, none of the evaluated variables adjusted to the linear model based on the significance of the regression coefficients and determination coefficient (R2), assuming 5% probability by the F-test and also considering the biological phenomenon under study. For this reason, the Tukey test of averages was also adopted in the quantitative factor to support the discussion of this work.
RESULTS AND DISCUSSION
General anatomic description
The leaf of C. abyssinica is amphistomatic, with uniseriate epidermis constituted by cells of varying shapes and sizes, covered by a thin cuticle. Glandular and tector trichomes occur sparsely on both surfaces of the epidermis (data not shown). The mesophyll is dorsiventral, formed by one or two cell layers of palisade parenchyma, and spongy parenchyma with three to four cell layers. Vascular bundles of collateral type and of small size occur across the mesophyll. The midrib, in cross section, appears concave-convex, with epidermis formed by round cells. The vascular system is predominantly formed by a single collateral vascular bundle (Figures 1 and 2).


The leaf anatomy of C. abyssinica is similar to that described for other representatives of Brassicaceae, such as Brassica napus L. (colza), Brassica gravinae Ten., Brassica rapa L. (turnip), Raphanus sativus L. (radish), Diplotaxis tenuifolia (Ueno, 2011) and Arabidopsis thaliana (L.) Heynh (Boeger and Poulson, 2006).
Survival and morphological evaluations
The survival percentage of plantlets originated from the four subcultures did not have any significant difference among the treatments, varying from 96.66% (at 90 days) to 100% (at 30, 60 and 120 days). These results disagree with those obtained by Pennell (1987), who relates that the higher the number of subcultures in the micropropagation, the lower the rate of survival. With this result, it can be highlighted that the utilization of apical segments obtained from plantlets germinated in vitro, in active growth, is preferable for the establishment of a micropropagation process, since the explants present very high rates of survival with the increasing number of subcultures. The oxidation, contamination and eventual death of the explant are problems for the in vitro establishment of some species owing to their peculiar characteristics (Sato et al., 2001).
By analyzing the summary of the analysis of variance, the induction of shoots in Crambe was more significantly influenced by the tested concentrations than by the types of cytokinins applied. The analysis of variance for the number of shoots per explant (NSE) after 30, 60, 90 and 120 days of cultivation in vitro are shown in Table 1. This analysis did not demonstrate significance in the interactions between the factors cytokinins and concentration in any of the evaluation times. In the factor cytokinin, significance was only found at 90 days of cultivation (Table 1). For the factor concentration, significance was found at 60, 90 and 120 days (Table 1).

For the morphologic variables evaluated in crambe, it is noted that there was difference among the cytokinins BAP, KIN and TDZ, influencing the development of new shoots and their growth. The differences in the factor concentration showed the necessity of cytokinin in the culture medium at the tested concentrations to stimulate the shoot in apical segments of crambe (Table 1).
For NSE at 90 days (Table 2) in the factor cytokinin, BAP (10.46) and TDZ (10.03) did not differ statistically. Thus, the application of BAP or TDZ at the tested concentrations, promoted significant increase in the induction of shoots in comparison with KIN. The production of nodal segments is an important variable in the in vitro propagation of Crambe, as it reflects the production of new plants at every subculture. According to Flores et al. (2009), BAP is the most efficacious cytokinin for the multiplication of aerial parts of most species. Furtado et al. (2007) described that TDZ and BAP induced greater formation of shoots on the cotyledonary bud explant in all treatments of peanut (Arachis hypogaea L.– Fabaceae), showing higher mean values than for KIN. Marino and Bertazza (1990) obtained greater proliferation of shoots when using BAP than KIN or zeatin for micropropagation of kiwi (Actinidia deliciosa A. Chev. - Actinidiaceae). Grattapaglia and Machado (1998) indicated that the higher efficiency of BAP in comparison with KIN might rely on the capacity of the plant tissues of metabolizing synthetic growth regulators more rapidly than plant hormones. According to the authors, this may vary depending on the used plant species. Krikorian (1991) comments that, at present, BAP has been more used than the cytokinins KIN and TDZ due to being a more active compound which can be easily found and at reasonable price.

In the tested concentrations of cytokinins, the highest values of NSE (10.43) were found at 5 µM (Table 3), being statistically superior to 10 µM at 60 days. After 90 days, NSE value at the concentration of 5 µM (11.26) did not differ from 10 µM (10.93), which did not change at 120 days, with NSE values of 10.96 for 5 µM and 10.20 for 10 µM. In all time points, the lowest values were observed in the control, in the absence of the growth regulator, demonstrating the importance of an exogenous cytokinin to stimulate the shoot induction in the process of micropropagation in crambe. This way, according to Grattapaglia and Machado (1998), the choice of cytokinin type and its concentration are factors that influence the success of multiplication in vitro, indispensibly, during the process of overcoming the apical dominance and induction of proliferation of axillary buds. This way, the concentration of 5 µM for the tested cytokinins was sufficient to induce efficient shooting in apical explants of crambe.

Li et al. (2010), working with hypocotyls of C. abyssinica cv. Galactica, described a protocol for direct induction of shoots in MS medium supplemented with 10 µM TDZ and 2.7 µM α-naphthaleneacetic acid (NAA), promoting a regeneration frequency of up to 60%. In a new work, Li et al. (2011) obtained 95% of regeneration of shoots from hypocotyl segments, using the medium Lepiovre (Quoirin and Lepoivre, 1977) again supple-mented with 10 µM TDZ and 2.7 µM NAA. Chhikara et al. (2012) demonstrated that the regeneration of shoots from calli obtained from hypocotyls of crambe presented greater efficiency in the medium, with 8.8 µM BAP and 0.10 µM NAA, with a frequency of 70%. Although a different explant was used in the present work than in the aforementioned works with crambe, it was noticed that the concentrations of the used cytokinins were close in both works, and efficiently promoted the expected morphogenetic response.
Anatomic evaluations
Regarding the leaf anatomy characteristics evaluated after 30 days (Table 4) in the plantlets cultivated in MS medium with different cytokinins at distinct concentrations, the analysis of variance has revealed no significant interaction between the employed factors. The various sources of cytokinins × concentrations used in this work did not exhibit significant effects on the thickness of the blade, mesophyll, abaxial epidermis, spongy parenchyma, midrib and area of the vascular bundle of crambe leaves at 30 days of cultivation. Only the adaxial epidermis and the palisade parenchyma presented significant differences in relation to the tested concentrations independently of the employed cytokinin type.

Table 5 shows the summary of the analysis of variance for the anatomic characteristics of crambe leaves, cultivated in vitro with different cytokinins at distinct concentrations, after 120 days. There was significant interaction between the tested factors for the thickness of the mesophyll and palisade parenchyma. Analyzing the factor concentration, only the thickness of the adaxial epidermis demonstrated statistical difference. Just as observed in the analysis carried out at 30 days, variations among the treatments were only observed for the adaxial epidermis and the palisade parenchyma. It is suggested that, at 30 days, owing to the short time of contact of the cytokinin with the explant, this factor may thus far not have influenced in such way as to alter effectively the thickness or structure of the evaluated characteristics. However, in analysis at 120 days, it was noted that few alterations remained. This result corroborates the fact that the in vitro multiplication of Crambe shoots under effect of the tested cytokinins does not alter the morphoanatomy of the leaves until day 120 of cultivation.
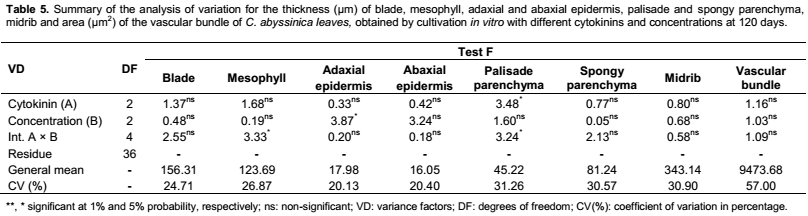
The thickness of the adaxial epidermis after 30 days and 120 days, and of the palisade parenchyma after 30 days of crambe leaves cultivated in vitro with cytokinins at different concentrations is represented in Table 6. The thickness of the adaxial epidermis (30 days) and of the palisade parenchyma presented similar behavior, in that their highest values were recorded for the concentration of 0 µM (21.69 and 47.74 µm, respectively); this differed statistically from the values obtained for 5 µM (16.16 and 29.68 µm, respectively), which were lowest. The treatments with 0 µM and 5 µM yielded the highest values of the palisade parenchyma after 120 days (20.10 and 17.10 µm, respectively), though not differing between themselves, and the latter not differing from the thickness at 10 µM (16.75 µm).

The use of cytokinins in the culture medium may induce the production of cells with decreased size during cell division (Pereira et al., 2000), explaining the reduction in the thickness of the adaxial epidermis and of the palisade parenchyma in the culture media that contained exogenous cytokinin, in comparison with the control at 30 days. The increase in thickness of the leaf epidermis seen in the control is one of the protection mechanisms through reducing the penetration of radiation into the sensitive tissues of the leaf mesophyll (Sullivan et al., 1996).
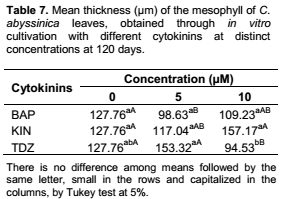
For the mean thickness of the mesophyll after 120 days of cultivation (Table 7), only TDZ demonstrated difference among its tested concentrations, with the highest value (153.32 µm) being observed at 5 µM, though not differing from the control (127.76 µm) or KIN (117.04 µm). In turn, at the concentration of 10 µM, the highest value (157.17 µm) was observed for KIN, not differing from BAP (109.23 µm). According to Araujo et al. (2009), the greater the thickness of the mesophyll, the higher the photosynthetic efficiency. Since one of the main problems of acclimatization is related to the low photosynthetic rates, the excess of cytokinins may hamper this step. In accordance with Oliveira et al. (2008), in their work with pond-apple (Annona glabra – Annonaceae) using different cytokinins in the concentration of 5 µM, the thickness of the mesophyll was affected by the presence of BAP and KIN, and not by TDZ and ZEA (zeatin). This result diverges from that obtained in the present work at 120 days, in which only the presence of TDZ differed from the control.
In the thickness of the palisade parenchyma after 120 days (Table 8) no significant difference was observed among the tested cytokinin concentrations. However, among the cytokinins, difference occurred at 10 µM KIN (63.49 µM), which differed significantly from the others. Overall, the highest thickness values were found at the concentration of 0 µM, repeating the results observed at 30 days. Prolonged use of cytokinins may exert phytotoxic effect, which is mainly characterized by the lack of elongation and reduction in the size of the leaves (Grataplaglia and Machado, 1998). The palisade parenchyma is responsible for the photosynthesis, with its thickening being essential for allowing higher fixation of CO2, contributing to the autotrophy of the plant after transfer into ex vitro environment (Silva et al., 2008; Castro et al., 2009).

Cross sections of the mesophyll of crambe leaves collected at the beginning of the in vitro multiplication (30 days), under effect of the cytokinins × concentrations, are shown in Figure 1. Although very discrete, it is still possible to notice the mesophyll limit, which consists of palisade parenchyma, with one or two layers of juxtaposed cells showing little elongation, and of spongy parenchyma, with three to four layers of weakly organized cells.
In Figure 1, it is remarkable that the appearance of the mesophyll in vitro treated with 0 µM cytokinin presented greater length compared to other concentrations. It is also possible to observe that, in the treatment with 10 µM cytokinin, a higher concentration of cells exists (variable not evaluated) (Figure 1C). The cytokinins are associated with the stimulation of cell division (Howell et al., 2003), which explains the smaller length and higher number of cells observed in the treatments with cytokinins.
The anatomic observations of mesophyll at 120 days demonstrated that there is no marked difference among the effects of the cytokinins at 5 µM (Figure 2A to C). However, at 10 µM KIN is superior to other cytokinins (10 µM BAP and TDZ) (Figure 2D to F), and the palisade parenchyma presents more elongated cells, characterizing more markedly the mesophyll limit of the mesophyll and demonstrating greater differentiation. Oliveira et al. (2008) also verified that leaves of plants subjected to treatment with BAP and KIN presented greater differentiation of the mesophyll compared to the treatments with TDZ and ZEA, with reduced proportion of intercellular spaces; this suggests higher efficaciousness of these two sources of cytokinin on the development of the assimilatory system of the plants during cultivation in vitro. The data from the aforementioned author corroborates that described for the present work at 120 days, in which the treatments with BAP and KIN at 5 and 10 µM demonstrated greater differentiation between the parenchyma.
The various sources of cytokinins at different concentrations used did not promote changes in the sense of modifying the organization and/or thickness of the leaves compared to the control. Consequently, the treatments employed in this experiment did not alter the leaves morphologically.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
ACKNOWLEDGMENT
The present research was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
REFERENCES
|
Apóstolo NM, Brutti CB, Llorente BE (2005). Leaf anatomy of Cynara scolymus L. in successive micropropagation stages. In Vitro Cell. Dev. Biol. Plant 41:307-313. |
|
|
Appezzato-Da-Glória B, Vieira MLC, Dornelas MC (1999). Anatomical studies of in vitro organogenesis induced in leaf-derived explants of passionfruit. Pesqui. Agropecu. Bras. 34(11):2007-2013. |
|
|
Araujo AGDe, Pasqual M, Miyata LY, Castro EMDe, Rocha HS (2009). Qualidade de luz na biometria e anatomia foliar de plântulas de Cattleya loddigesii L. (Orchidaceae) micropropagadas. Ciênc. Rural 39(9):2506-2511. |
|
|
Boeger MRT, Poulson ME (2006). Efeitos da radiação ultravioleta-B sobre a morfologia foliar de Arabidopsis thaliana (L.) Heynh. (Brassicaceae). Acta Bot. Brasilica 20(2):329-338. |
|
|
Capelle A, Tittonel ED (1999). Crambe, a potential non-food oil crop: production. Agro Food Industry Hi-Tech 10:22-27. |
|
|
Castro EM, Pereira FJ, Paiva R (2009). Histologia vegetal: estrutura e função de órgãos vegetativos. 1st Ed, Editora UFLA, Lavras-MG, v. 1, 234p. |
|
|
Chhikara S, Dutta I, Paulose B, Jaiwal PK, Dhankher OP (2012). Development of an Agrobacterium-mediated stable transformation method for industrial oilseed crop Crambe abyssinica 'BelAnn'. Ind. Crops Prod. 37:457-465. |
|
|
Flores R, Nicoloso FT, Maldaner J, Garlet TMB (2009). Benzilaminopurina (BAP) e thidiazuron (TDZ) na propagação in vitro de Pfaffia glomerata (Spreng.) Pedersen. Rev. Bras. Plantas Med. 11(3):292-299. |
|
|
Furtado CM, Carvalho JMFC, Castro JPDe, Silva H (2007). Comparação da frequência de regeneração in vitro do amendoim (Arachis hipogaea), utilizando diferentes citocininas. Rev. Biol. Ciênc. Terra 7(1):51-58. |
|
|
Gao HB, Wang Y, Gao F, Luo P (1998). Studies on the Plant regeneration from single cell culture of Crambe abyssinica. Hereditas 20:50-52. |
|
|
Grattapaglia D, Machado MA. Micropropagação (1998). In: Torres AC, Caldas LS, Buso JA. Cultura de tecidos e transformação genética de plantas. Embrapa-SPI/Embrapa-CNPH, Brasília, v.1, p. 183-260. |
|
|
Hazarika BN (2006). Morpho-physiological disorders in in vitro culture of plants. Sci. Hortic. 108:105-120. |
|
|
Howell SH, Lall S, Che P (2003). Cytokinins and shoot development. Trends Plant Sci. 8(9):453-459. |
|
|
Hronkova M, Zahradnickova H, Simkova M, Simek P, Heydová A (2003). The role of abscisic acid in acclimation of plants cultivated in vitro to ex vitro conditions. Biol. Plant. 46:535-541. |
|
|
Johansen DA (1940). Plant microtechnique. 2. ed. Mc-Graw-Hill, New York. 523p. |
|
|
Kadlecek P, Tichá I, Haisel D, Capková V, Schafer C (2001). Importance of in vitro pretreatment for ex vitro acclimatization and growth. Plant Sci. 161:695-701. |
|
|
Kozai T (1991). Micropropagation under photoautotrophic conditions. In: Debergh PC, Zimmerman RH (eds.). Micropropagation: Technology and Application, Kluwer, Dordrecht. pp. 447-469. |
|
|
Krikorian AD (1991). Medios de cultivo: generalidades, composición y preparación. In: Roca WR, Mroginski LA. Cultivo de tejidos en la agricultura: fundamentos y aplicaciones. Cali: Centro Internacional de Agricultura Tropical. pp.41-78. |
|
|
Li X, Ahlman A, Lindgren H, Zhu LH (2011). Highly efï¬cient in vitro regeneration of the industrial oilseed crop Crambe abyssinica. Ind. Crops Prod. 33:170-175. |
|
|
Li X, Ahlman A, Yan X, Lindgren H, Zhu L-H (2010). Genetic transformation of the oilseed crop Crambe abyssinica. Plant Cell Tissue Organ Cult. 100:149-156. |
|
|
Marino G, Bertazza G (1990). Micropropagation of Actinidia deliciosa Cvs Hayward and Tomuri. Sci. Hortic. 45:65-74. |
|
|
Murashige T, Skoog FA (1962). A revised medium for rapid growth and bioassays with Tabaco tissue cultures. Physiol. Plant. 15(3):473-497. |
|
|
Murphy DJ (1999). Production of novel oils in plants. Curr. Opin. Biotechnol. 10:175-180. |
|
|
O'brien TP, Feder N, Mccully ME (1964). Polychromatic staining of plant cell walls by toluidine blue. Protoplasma 59(2):368-373. |
|
|
Oliveira LMDe, Paiva R, Aloufa MAI, Castro EMDe, Santana JRFDe, Nogueira RC (2008). Efeitos de citocininas sobre a anatomia foliar e o crescimento de Annona glabra L. durante o cultivo in vitro e ex vitro. Ciênc. Rural 38(5):1447-1451. |
|
|
Palmer CD, Keller WA (2011). Somatic embryogenesis in Crambe abyssinica Hochst. Ex R.E. Fries using seedling explants. Plant Cell Tissur Organ Cult. 104:91-100. |
|
|
Pennell D (1987). Strawberry micropropagation within the UK. In: Boxus P, Larvor P (eds.). In vitro culture of strawberry plants. Bruseels: Commission of the European Communities. pp. 27-34. |
|
|
Pereira FD, Pinto JEBP, Cardoso MG, Lameira OA (2000). Propagação in vitro de chapéu-de-couro (Echinodorus cf. scaber Rataj), uma planta medicinal. Ciência e Agrotecnologia 24(Edição especial):74-80. |
|
|
Quoirin M, Lepoivre P (1977). Étude de milieux adaptés aux cultures in vitro de Prunus. Acta Hortic. 78:437-442. |
|
|
Sato AY, Teixeira HCD, Andrade LAde, Souza VCde (2001). Micropropagação de Celtis sp: controle da contaminação e oxidação. Cerne 7(2):117-123. |
|
|
SigmaPlot: Exact Graphs and Data Analysis (2006). Version 10.0. Systat Software, Alemanha. |
|
|
Silva AB, Pasqual M, Castro EMde, Miyata LY, Melo LA, Braga FT (2008). Luz natural na micropropagação do abacaxizeiro (Ananas comosus L. Merr). Interciência 33(11):839-843. |
|
|
Silva FASE, Azevedo CAV (2009). Principal Components Analysis in the Software Assistat-Statistical Attendance. In: World Congress On Computers In Agriculture, 7, Reno-NV-USA: American Society of Agricultural and Biological Engineers. |
|
|
Sullivan JH, Howells BW, Ruhland CT, Day TA (1996). Changes in leaf expansion and epidermal screening effectiveness in Liquidambar styraciflua and Pinus taeda in response to UV-B radiation. Physiol. Plant. 98:349-357. |
|
|
Ueno O (2011). Structural and biochemical characterization of the C3–C4 intermediate Brassica gravinae and relatives, with particular reference to cellular distribution of Rubisco. J. Exp. Bot. 62(15):5347-5355. |
|
|
Vargas-Lopez JM, Wiesenborn D, Tostenson K, Cihacek L (1999). Processing of Crambe oil and isolation of erucic acid. J. Am. Oil Chem. Soc. 76:801-809. |
|
|
Wang Y, Peng P (1998). Intergeneric hybridization between Brassica species and Crambe abyssinica. Euphytica 101:1-7. |
|
|
Wang YP, Snowdon RJ, Rudloff E, Wehling P, Friedt W, Sonntag K (2004). Cytogenetic characterization and fae1 gene variation in progenies from asymmetric somatic hybrids between Brassica napus and Crambe abyssinica. Genome 47:724-731. |
|
|
Wang YP, Sonntag K, Rudloff E (2003). Development of rapeseed with high erucic acid content by asymmetric somatic hybridization between Brassica napus and Crambe abyssinica. Theor. Appl. Genet. 106:1147-1155. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


