Full Length Research Paper

ABSTRACT
Antibacterial activity of extracts of Zanthoxylum gilletii, Markhamia lutea and their fungal endophytes were evaluated against bacterial pathogens of common bean: Xanthomonus axonopodis pv. phaseoli and Pseudomonus syringae pv. phaseolicola. The leaves of both plants were dried under shade, ground to fine powder and extracted using methanol. The methanol extracts were fractionated sequentially using ethylacetate and hexane to produce various fractions. Endophytic fungi were isolated from fresh leaves and identified by ITS-rDNA sequence analysis. Antibacterial screening of the fungal endophytes was done by dual culture assay. The most active endophytic fungi were fermented on rice media and extracted using methanol. Pure compounds were analyzed by a combination of mass spectrometry and spectroscopic techniques which included 1D and 2D NMR. Antibacterial activity of all the extracts was determined by disc agar diffusion assay against the test organisms. Twenty-four (24) fungal endophytes were isolated which included: Fusarium, Chaetomium, Scopulariopsis and Trametes. Endophytic Fusarium solani was the most active against X. axonopodis pv. phaseoli (20.3 mm inhibition zone) and P. syringae pv. phaseolicola (18.6 mm inhibition zone). The plant extracts were active against X. axonopodis pv. phaseoli with an inhibition zone ranging between 8-12 mm except the methanol extract from Z. gilletii which did not show any activity. The endophytic extracts were active against both test organisms with a zone of inhibition ranging from 9.3-14 mm. Phenolic compounds present in Fusarium species may have contributed to the antibacterial activity of this strain against the test organisms.
Key words: Common bean, medicinal plants, fungal endophytes, antibacterial activity, Xanthomonus axonopodis pv phaseoli, Pseudomonus syringae pv phaseolicola.
INTRODUCTION
MATERIALS AND METHODS
RESULTS
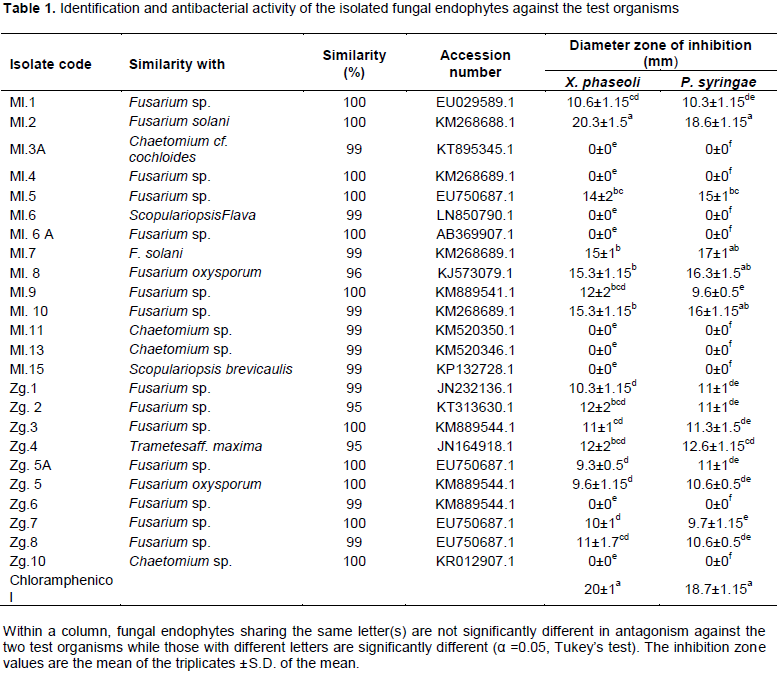
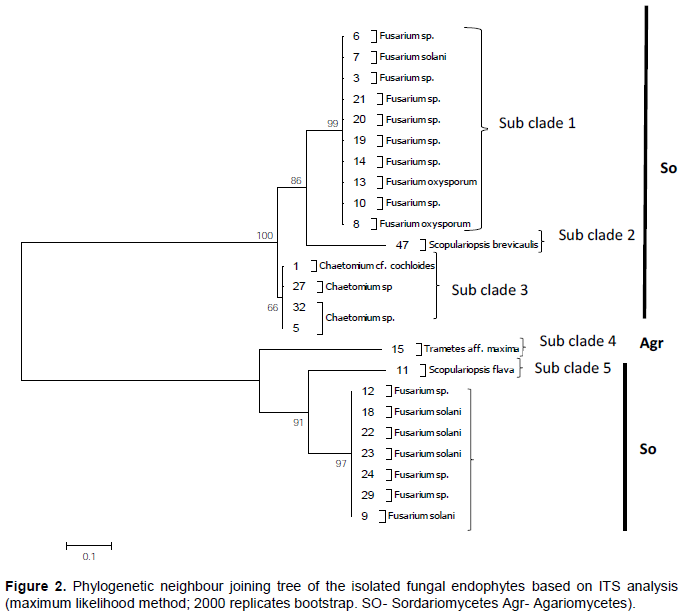
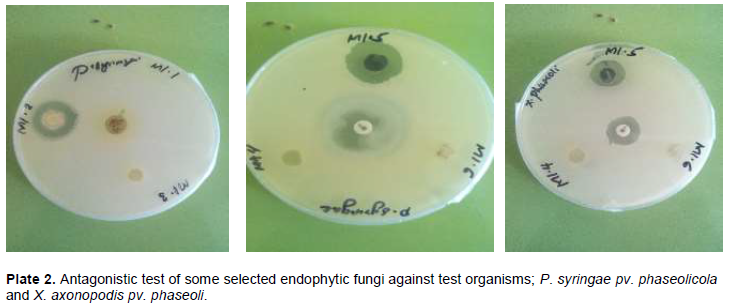
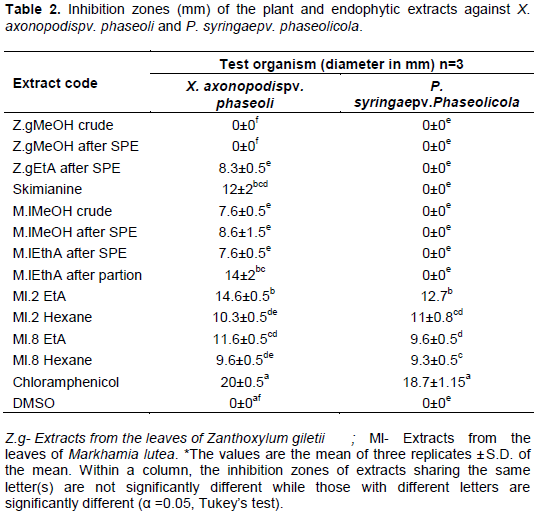
DISCUSSION
CONCLUSION
This study demonstrated that the leaves of Z. gilletii and M. lutea are inhabited by different strains of endophytic fungi with promising benefits in controlling X. axonopodis pv. phaseoli and P. syringae pv. phaseolicola. The results indicated that Fusarium species contains secondary metabolites that can be used as antibacterial agents against these two bacterial pathogens. The leaf extracts of both plants also contain secondary metabolites that can be used directly or incorporated in other available pesticides to control or manage these infections in common bean.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the financial support provided by ERAfrica ASAFEM Project (ERAFRICA_RE-70). They thank the Helmholtz Zentrum FürInfektionforschung (HZI) in Braunschweig, Germany (especially Prof. Dr. Marc Stadler and Ms. Clara Chepkirui) for availing the laboratory space and guidance in conducting the molecular aspects of this study.
REFERENCES
|
Bacon CW, Yates IE (2006). Endophytic root colonization by Fusarium species: histology, plant interactions, and toxicity. In. Microbial root endophytes. Springer, Berlin, Heidelberg pp. 133-152. |
|
|
Bai WD, Kai HL, Wen QC, Xiao WD, Xiu CX (2009). Fusarium solani, Tax-3, a new endophytic taxol-producing fungus from Taxus chinensis. World Journal of Microbiology and Biotechnology 25:139-143. |
|
|
Banhos EFD, Souza AQLD, Andrade JCD, Souza ADLD, Koolen HHF, Albuquerque PM (2014). Endophytic fungi from Myrcia guianensis at the Brazilian Amazon: distribution and bioactivity. Brazilian Journal of Microbiology 45(1):153-162. |
|
|
Bashyal BP, Wijeratne EK, Faeth SH, Gunatilaka AL (2005). Globosumones A− C, Cytotoxic Orsellinic Acid Esters from the Sonoran Desert Endophytic Fungus Chaetomium globosum. Journal of Natural Products 68(5):724-728. |
|
|
Chen XM, Dong HL, Hu KX, Sun ZR, Chen J, Guo SX (2010). Diversity and antimicrobial and plant-growth-promoting activities of endophytic fungi in Dendrobium loddigesii Rolfe. Journal of Plant Growth Regulation 29:328-337. |
|
|
Clay K (1990). Fungal endophytes of grasses. Annual Review of Ecology and Systematics 21(1):275-297. |
|
|
Crozier J, Thomas SE, Aime MC, Evans HC, Holmes KA (2006). Molecular characterization of fungal endophytic morphospecies isolated from stems and pods of Theobroma cacao. Plant Pathology 55(6):783-791. |
|
|
Deng BW, Liu KH, Chen WQ, Ding XW, Xie XC (2009). Fusariumá solani, Tax-3, a new endophytic taxol-producing fungus from Taxus áchinensis. World Journal of Microbiology and Biotechnology 25:139. |
|
|
Devaraju R, Satish S (2011). Endophytic Mycoflora of Mirabilis jalapa L. and Studies on Antimicrobial Activity of its Endophytic sp. Society of Applied Sciences 2(1):75-79. |
|
|
Gichangi A, Maobe SN, Karanja D, Getabu A, Macharia CN, Ogecha JO, Nyang'au MK, Basweti E, Kitonga L (2012). Assessment of production and marketing of climbing beans by smallholder farmers in Nyanza region, Kenya. World Journal of Agricultural Sciences 8:293-302. |
|
|
Gong L, Guo S (2009). Endophytic fungi from Dracaena cambodiana and Aquilaria sinensis and their antimicrobial activity. African Journal of Biotechnology 8(5):731-736. |
|
|
Hassan Eman O. El-Meneisy Afaf ZA (2014). Biocontrol of halo blight of bean caused by Pseudomonas phaseolicola. International Journal of Virology 10:235-242. |
|
|
Hidayat I, Radiastuti N, Rahayu G, Achmadi S, Okane I (2016). Three Quinine and Cinchonidine producing Fusarium species from Indonesia. Current Research in Environmental and Applied Mycology 6:20-34. |
|
|
Ibrahim M, Kaushik N, Sowemimo A, Chhipa H, Koekemoer T, van de Venter M, Odukoya OA (2017). Antifungal and anti-proliferative activities of endophytic fungi isolated from the leaves of Markhamia tomentosa. Pharmaceutical Biology 55(1):590-595. |
|
|
Ihrmark K, Bödeker IT, Cruz-Martinez K, Friberg H, Kubartova A, Schenck J, Lindahl BD (2012). New primers to amplify the fungal ITS2 region–evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiology Ecology 82(3):666-677. |
|
|
Kajaria D, Gangwar M, Kumar D, Sharma AK, Nath G, Tilak R, Tiwari SK (2012). Qualitative phytochemical characterization and antimicrobial evaluation of a polyherbal compound-Bharangyadi. Journal of Pharmacy Research 5:416-419. |
|
|
Katungi E, Farrow A, Mutuoki T, Gebeyehu S, Karanja D, Alamayehu F, Sperling L, Beebe S, Rubyogo JC, Buruchara R (2010). Improving common bean productivity: An Analysis of socioeconomic factors in Ethiopia and Eastern Kenya. Baseline Report Tropical Legumes II. Centro Internacional de Agricultura Tropical-CIAT. Cali, Colombia. |
|
|
Kusari S, Hertweck C, Spiteller M (2012). Chemical ecology of endophytic fungi: origins of secondary metabolites. Chemistry and Biology 19(7):792-798. |
|
|
Lamorde M, Tabuti JRS, Obua C, Kukunda-Byobona C, Lanyero H, Byakika-Kibwika P, Bbosa GS, Lubega A, Ogwal-Okeng J, Ryan M. (2010). Medicinal plants used by traditional medicine practitioners for the treatment of HIV/AIDS and related conditions in Uganda. Journal of Ethnopharmacology 130:43-53. |
|
|
Li W, Zhou J, Guo S, Guo L (2007). Endophytic fungi associated with lichens in Baihua mountain of Beijing, China. Fungal Diversity 25:69-80. |
|
|
Momesso LDS, Kawano CY, Ribeiro PH, Nomizo A, Goldman GH, Pupo MT (2008). Chaetoglobosins produced by Chaetomiumglobosum, endophytic fungus found in association with Viguierarobusta Gardn (Asteraceae). Química Nova 31(7):1680-1685. |
|
|
Nascimento AMD, Conti R, Turatti IC, Cavalcanti BC, Costa-Lotufo LV, Pessoa C, Pupo MT (2012). Bioactive extracts and chemical constituents of two endophytic strains of Fusarium oxysporum. Revista Brasileira de Farmacognosia 22(6):1276-1281. |
|
|
Negi JS, Bisht VK, Bh AK, Singh P, Sundriyal RC (2011). Chemical constituents and biological activities of the genus Zanthoxylum: a review. African Journal of Pure and Applied Chemistry 5(12):412-416. |
|
|
Nyunja ARO, Onyango JC, Erwin B (2009). The Kakamega forest medicinal plant resources and their utilization by the adjacent Luhya community. International Journal of Tropical Medicine 4:82-90. |
|
|
Orwa C, Mutua A, Kindt R, Jamnadass R, Simons A (2009). Agroforestree database: a tree species reference and selection guide version 4.0. World Agroforestry Centre ICRAF, Nairobi, KE. |
|
|
Paterson RR (2004). The isoepoxydon dehydrogenase gene of patulin biosynthesis in cultures and secondary metabolites as candidate PCR inhibitors. Mycological Research 108(12):1431-1437. |
|
|
Puri SC, Nazir A, Chawla R, Arora R, Riyaz-ul-Hasan S, Amna T, Ahmed B, Verma V, Singh S, Sagar R (2006). The endophytic fungus Trametes hirsuta as a novel alternative source of podophyllotoxin and related aryl tetralin lignans. Journal of Biotechnology 122:494-510. |
|
|
Qin JC, Zhang YM, Gao JM Bai MS, Yang SX, Laatsch H, Zhang AL (2009). Bioactive metabolites produced by Chaetomiumglobosum, an endophytic fungus isolated from Ginkgo biloba. Bioorganic & Medicinal Chemistry Letters 19(6):1572-1574. |
|
|
Rani R, Sharma D, Chaturvedi M, Yadav JP (2017). Antibacterial Activity of Twenty Different Endophytic Fungi Isolated from Calotropis procera and Time Kill Assay. Clinical Microbiology 6:3. |
|
|
Rivera-Ordu-a FN, Suarez-Sanchez RA, Flores-Bustamante ZR, Gracida-Rodriguez JN, Flores-Cotera LB (2011). Diversity of endophytic fungi of Taxus globosa (Mexican yew). Fungal Diversity 47:65-74. |
|
|
Sartori AF, Reis EM, Casa RT (2004). Quantificação da transmissão de Fusariummoniliforme de sementesparaplântulas de milho. Fitopatologia Brasileira 29(4):456-458. |
|
|
Sati SC, Joshi S (2011). Antibacterial activities of Ginkgo biloba L. leaf extracts. The Scientific World Journal 11:2237-2242. |
|
|
Shiono Y, Tsuchinari M, Shimanuki K, Miyajima T, Murayama T, Koseki T, Laatsch H, Funakoshi T, Takanami K, Suzuki K (2007). Fusaristatins A and B, two new cyclic lipopeptides from an endophytic Fusarium sp. Journal of Antibiotics 60:309. |
|
|
Specian V, Sarragiotto MH, Pamphile JA, Clemente E (2012). Chemical characterization of bioactive compounds from the endophytic fungus Diaporthe helianthi isolated from Luehea divaricata. Brazilian Journal of Microbiology 43(3):1174-1182. |
|
|
Srivastava A, Anandrao RK (2015). Antimicrobial potential of fungal endophytes isolated from leaves of Prosopis juliflora (sw.) Dc. An important weed. International Journal of Pharmacy and Pharmaceutical Sciences 7:128-136. |
|
|
Stangarlin JR, Kuhn OJ, Assi L, Schwan-Estrada KRF (2011). Control of plant diseases using extracts from medicinal plants and fungi. Science against microbial pathogens: communicating current research and technological advances. Badajoz: Formatex 2:1033-1042. |
|
|
Suryanarayanan TS (2011). Diversity of fungal endophytes in tropical trees. In. Endophytes of forest trees. Springer Netherlands pp. 67-80. |
|
|
Suryanarayanan TS, Thirunavukkarasu N, Govindarajulu MB, Sasse F, Jansen R, Murali TS (2009). Fungal endophytes and bioprospecting. Fungal Biology Reviews 23(1):9-19. |
|
|
Tavares LDC, Zanon G, Weber AD, Neto AT, Mostardeiro CP, Da Cruz IB, Morel AF (2014). Structure-activity relationship of benzophen anthridine alkaloids from Zanthoxylum rhoifolium having antimicrobial Activity. PloS one 9(5):e97000. |
|
|
Tayung K, Barik BP, Jha DK, Deka DC (2011). Identification and characterization of antimicrobial metabolite from an endophytic fungus, Fusarium solani isolated from bark of Himalayan yew. Mycosphere 2:203-213. |
|
|
Vaz AB, Mota RC, Bomfim MRQ, Vieira ML, Zani CL, Rosa CA, Rosa LH (2009). Antimicrobial activity of endophytic fungi associated with Orchidaceae in Brazil. Canadian Journal of Microbiology 55(12):1381-1391. |
|
|
Waithaka PN, Gathuru EM, Githaiga BM, Onkoba KM (2017). Antimicrobial Activity of Mushroom (Agaricus bisporus) and Fungal (Trametes gibbosa) Extracts from Mushrooms and Fungi of Egerton Main Campus, Njoro Kenya. Journal of Biomedical Sciences, 6:3. |
|
|
Wiese J, Ohlendorf B, Blümel M, Schmaljohann R, Imhoff JF (2011). Phylogenetic identification of fungi isolated from the marine sponge Tethya aurantium and identification of their secondary metabolites. Marine Drugs 9(4):561-585. |
|
|
Xing YM, Chen J, Cui JL, Chen XM, Guo SX (2011). Antimicrobial Activity and Biodiversity of Endophytic Fungi in Dendrobium devonianum and Dendrobium thyrsiflorum from Vietman. Current Microbiology 62:1218-1224. |
|
|
Zinniel DK, Lambrecht P, Harris NB, Feng Z, Kuczmarski D, Higley P, Vidaver AK (2002). Isolation and characterization of endophytic colonizing bacteria from agronomic crops and prairie plants. Applied and Environmental Microbiology 68(5):2198-2208. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


