Full Length Research Paper

ABSTRACT
In the present study, plant growth promoting rhizobacterial (PGPR) strains Pseudomonas fluorescence strain P2, Pseudomonas jessenii R62, Pseudomonas synxantha R81, Bacillus cereus BSB 38 (14B), Arthrobacter nitroguajacolicus strainYB3 and strain YB5 were tested for their role in enhancing plant growth and induction of stress related enzymes in Sahbhagi (drought tolerance) and IR-64 (drought sensitive) cultivars of rice (Oryza sativa L.) under different level of drought stress. PGPRs, P. jessenii, R62, P. synxantha, R81 were used as one consortium similarly A. nitroguajacolicus strainYB3 and strain YB5 were used as other consortia. Most of the PGPR inoculated plants showed enhanced growth as compared to uninoculated plants under all the level of drought stress. Quantitative analyses of antioxidant enzymes indicated that majority of the PGPRs inoculated plants in both varieties showed higher proline content, higher activity of superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX) and lower level of hydrogen peroxide (H2O2), malondialdehyde (MDA) in leaves at all the level of drought stress. The study suggests that PGPRs alleviates oxidative damage in rice plants grown under drought by improving plant growth and activating antioxidant defense systems, thereby improving stability of membranes in plant cells. This study provides evidence for a beneficial effect of PGPRs application in enhancing drought tolerance of rice under water deficit conditions.
Key words: Plant growth promoting rhizobacterial (PGPR), plant growth promotion, drought stress, antioxidant, rice.
INTRODUCTION
Rice (Oryza sativa L.) is the staple food consumed by more than half of the world population and fulfills 23% of their caloric demands (Khush, 2003). Rice has semi-aquatic nature and grown under flooded condition conventionally to provide nutrient supply and bulky amounts of water. However due to insufficient water, half of the rice areas in the world do not maintained flooded condition and thus reduced yield, to some extent, as a result of drought (Bernier et al., 2008). Rice with little adaptation under water limited condition is remarkably sensitive to drought stress (Kamoshita et al., 2008). Under varieties of environmental stress including drought, plants showed increased level of reactive oxygen species (ROS) (Sgherri et al., 1996), which includes superoxide radical (O2._), hydroxyl free radical (OH), hydrogen peroxide (H2O2) and singlet oxygen resulting in peroxidation of lipids, denaturation of proteins, mutation of DNA and various types of cellular oxidative damage (Smirnoff, 1993).
Enhanced membrane lipid peroxidation takes place in both cellular and organelle membranes when ROS reaches above threshold level, which, in turn affect normal cellular functioning and act as an indicator of ROS mediated damage to cell membranes under stressful conditions (Mishra et al., 2011) and can be measured by malondialdehyde (MDA) content, one of the final products of peroxidation of unsaturated fatty acids in phospholipids of membrane (Halliwell and Gutteridge, 1989). Plant cells are protected against damaging effect of ROS by antioxidant defense system comprising enzymatic and non enzymatic component. In enzymatic component superoxide dismutase (SOD) catalyses the conversion of superoxide radical into H2O2 through their varietal isoforms; catalase (CAT) removes the bulk of H2O2 generated by photorespiration in peroxysomes; peroxidase (POD) acts on H2O2 for substrate oxidation in vacuole, cell wall and cytosol; ascorbate peroxidase (APX) is localized in cytosol and various organelles and catalyses the conversion of H2O2 into H2O and thus protected plants against detrimental effect of ROS (Noctor and Foyer, 1998). Usually, higher antioxidant activity in plants correlated with enhance resistant against stress (Sairam and Srivastava, 2001).
Plant growth promoting Rhizobacteria (PGPR) is well known for their growth-promoting properties like production of phytohormones, ability to solubilize mineral phosphate and to antagonize plant pathogens, etc. (Glick, 1995). PGPR like Pseudomonas fluorescens and Bacillus subtilis, recently have obtained attention as inoculants to withstand plants under varied biotic and abiotic stress conditions because of their excellent root colonizing ability, versatility in their catabolic activity, and their capacity to produce a wide range of metabolites and enzymes (Mayak et al., 2004; Saravanakumar and Samiyappan, 2007). Several authors have suggested the possible role of PGPRs to alleviate the oxidative damage elicited by abiotic stress through the manipulation of antioxidant enzymes in different crops (Kohler et al., 2008; Sandhya et al., 2010; Saravanakumar et al., 2011).
In present study we selected the Pseudomonas strain R62 and R81 because of their importance as a biofertilizer under field condition (Mader et al., 2011; Roesti et al., 2006). The Arthrobacter nitroguajacolicus were used as the ability of Arthrobacter species to showed resistance against desiccation, starvation and other stresses (Mongodin et al., 2006); and Bacillus and Pseudomonas has their importance in previous study to withstand plants under varied biotic and abiotic stress conditions (Mayak et al., 2004; Saravanakumar and Samiyappan, 2007). All the selected PGPRs were tested for their plant-growth promoting ability as well as their role in stress related enzymatic adaptive mechanisms in terms of antioxidant enzymes in tolerant and susceptible varieties of rice under drought stress.
MATERIALS AND METHODS
Bacterial inoculants
For the study, plant growth promoting bacterial strains Pseudomonas jessenii (R62), Pseudomonas synxantha (R81) (Mader et al., 2011; Roesti et al., 2006), two strains of Arthrobacter nitroguajacolicus, strainYB3 and YB5 (Gusain et al., 2015), Bacillus cereus BSB 38 (14B), and Pseudomonas fluorescence strain P2 were kindly provided by Rhizosphere biology lab of the Department of Biological Sciences of G. B. Pant University of Agriculture and Technology Pantnagar. In this study R62 and R81 were used as consortium (R62+R81), similarly both the strains of A. nitroguajacolicus YB3 and YB5 were used as consortium (A3+A5). All these strains grow separately in nutrient broth medium (Himedia, India) in flasks incubated at 28°C at 120 rpm until the late exponential phase. The final culture cfu was maintained at 107 to 108 cfu ml−1 level.
Rice varieties
Seeds of two genotype of rice, drought tolerant Sahbhagi (Mackill et al., 2010) and drought susceptible IR-64 (Singh and Ghosh, 2013) were kindly provided by the IRRI, Pusa New Delhi, India.
Pot experiment
Rice growth promotion by these bacterial strains under drought stress was performed in greenhouse condition (temperature: 27±2°C, photo period: 16/8 h day/night cycle, light intensity: 400 Em-2s-1, (400-700 nm), and relative humidity: 60%, respectively). Rice seeds were surface disinfected by immersion in 70% ethanol and 3% (v/v) sodium hypochlorite for 1 and 5 min. Seeds were washed thoroughly many times with sterile distilled water then germinated on sterilized Petri dish. The soil used for the experiment had pH 8.31, organic carbon of 1.2%, nitrogen of 186.7 kg/h, phosphorus of 34.91 kg/h, and potassium of 145.6 kg/h. Five hundred grams of the sterilized soil was filled in pots and watered to field capacity before sowing the seeds. At the time of sowing seeds in pots the bacterial inocula were given to 1 ml/pot. Two seedlings per pot were maintained. After 30 days of sowing, 10 ml of nutrient solution (Hoagland and Arnon, 1950) were given weekly to each pot to fulfill the nutrients requirements of the plants. The nine replicate of each treatment (including control) were arranged according to the complete randomized design. After 55 days of sowing, the pots were irrigated up to water holding capacity of soil and left for drought stress by withholding the irrigation. Leaf rolling in drought stress was determined based on rice standard evaluation system developed by International Rice Research Institute (IRRI). A visual score was taken of the degree of leaf rolling using a 0 to 9 scale (leaf rolling at vegetative stage) with 0 leaves for healthy while 9 for leaves tightly rolled V-shape. The plants were subjected to harvest at 0, 5 and 9 stages of leaf rolling (0, 8 and 10 days of drought) with three replicate, out of nine replicate, at each stage randomly selected for the measurement of growth promoting trait and antioxidant enzyme activities. After harvesting, fresh weight were taken immediately and sample were placed in -80°C for determination of antioxidant activity. Soil water content (SWC) for each harvesting was calculated using the weight fraction: SWC (%) = [(FW-DW)/DW] × 100, where FW was the fresh weight of a soil portion of the middle part of each pot and DW was the dry weight of the soil portion after drying in a hot air oven at 80°C for 48 h or till the complete drying of soil (Cha-um et al., 2012).
Estimation of chlorophyll and carotenoid content
Concentration of total chlorophyll and carotenoid was analysed following the method of Arnon (1949).
Estimation of H2O2, MDA and proline
For estimation of H2O2 and MDA content, leaf material (0.3 g) was homogenized in 4 ml of 0.1% trichloroacetic acid (TCA). Homogenate was centrifuged at 10,000 x g for 10 min at 4°C. MDA content was determined according to procedure of Heath and Packer (1968). The concentration of MDA was calculated by using an extinction coefficient of 155 mM-1 cm-1. Hydrogen peroxide was measured according to Alexieva et al. (2001). The amount of H2O2 was calculated using standard curve prepared with different dilutions of a working standard of 100 mM of H2O2. Free proline was determined by the method of Bates et al. (1973).
Estimation of anti-oxidative enzyme
For assays of SOD, CAT and POD, 0.5 g leaf samples (fresh weight) was homogenized with a pestle in an ice-cold mortar in 5 ml cold buffer containing: 50 mM potassium phosphate buffer (pH 7.0), 1 mM ethylene diamine tetra acetic acid (EDTA) and 1% (w/v) polyvinylpyrolidone (PVP). Whole extraction procedure was carried out at 4°C. The homogenate was centrifuged at 10,000 x g for 30 min at 4°C and the supernatant collected was used to assay enzymatic activity. For determination of APX activity 0.4 g leaf sample were homogenised in 4 ml of of ice-cold 25 mM phosphate buffer (pH 7-8) containing 1% PVP and 0.2 mM EDTA. The homogenates were filtered, and then centrifuged at 4°C for 15 min at 18000 g. All the antioxidant enzymes were determined as described by Zhang and Kirkham (1996). Protein concentration in the enzyme extract was determined by the method of Bradford (1976) using bovine serum albumin as a standard.
Statistical analysis
The data presented here are mean values ± SD. The data of individual stress level has three replicate (n=3) for each treatments of individual variety. The data were subjected to factorial analysis of variance (ANOVA), with varieties, stress level and treatments used for analysis and the differences between the means were compared using least significant differences at p<0.05. Different letters denote significant differences among treatments (including control) in two varieties.
RESULTS AND DISCUSSION
The plants were harvested at 0 days, 8 days and 10 days of drought and all these stages the soil moisture content of pot were measured as 640 ± 17.83, 62 ± 03.09 and 37 ± 03.30% respectively.
Plant growth parameters
The treatments showed varied effect on the shoot length, root length, shoot fresh weight and root fresh weight under all the level of drought stress. However, in majority of the inoculated plants showed higher effect on growth parameters as compare to uninoculated plants, although most of the differences are not significant. Irrespective of bacterial inoculation and stress level Sahbhagi showed enhanced effect on growth parameters as compared to IR-64 (Table 1). The PGPR can show the various kind of the plant growth promoting (PGP) activities which may be the mechanism through which they influence the plant growth promotion (Glick, 1995). The inoculation effect of our bacterial isolates had remarkable positive effect on plant fresh weight under non stress and stress condition. The higher growth enhancement effect of Sahbhagi as compared to IR-64 under drought stress might be related with better stress tolerance characteristics of the variety. Various study indicated that PGPRs inoculated plants can take up a higher volume of water and nutrients from rhizosphere soil; the attributes could be useful for the growth of plants under drought stress (Alami et al., 2000). However, the highest benefits of the PGPRs as bioinoculants can occur when crops faced prolonged stressful condition (Egamberdiyeva and Hoflich, 2004).
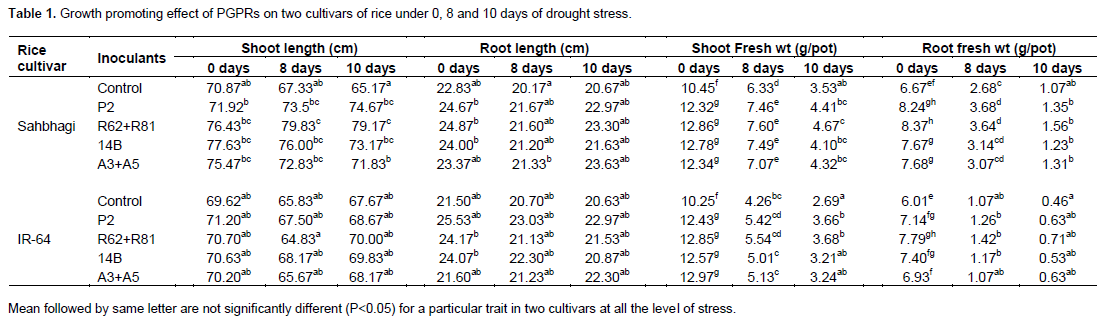
Chlorophyll and carotenoid content
In the present study, a gradual decrease in chlorophyll and caretenoid content was found with stress in both varieties. Similar to the growth parameters, majority of the treatments showed enhanced chlorophyll and carotenoid contents over control under all the level of stress (Table 2). The enhanced chlorophyll content may increase the photosynthetic efficiency of inoculated plants, and thus may be a reason for the tolerance of abiotic stress. Increased chlorophyll and caretenoid content in plants inoculated with PGPR is supported by previous study (Gururani et al., 2013) where authors found improved photosynthetic performance in Solanumtuberosum when inoculated with Bacillus sp. under abiotic stress.
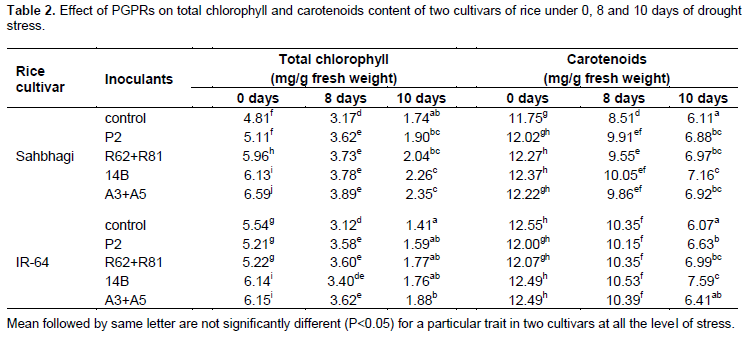
Hydrogen peroxide content
Among all the treatments, P2 treated plants in Sahbhagi and IR-64 showed maximum reduction in H2O2 level with 1.17 and 1.59 fold respectively, over control in 8th days of drought. In 10th days of drought, only R62+R81 treated plants in Sahbhagi with 1.15 fold reduction showed significant effect on H2O2 level, while in IR-64, all the PGPRs treated plants significantly reduced the H2O2 level over control plants (Figure 1a). Under drought, increased level of H2O2 was well documented which may be due to the formation of superoxide ion by electron transport chains which dismutase to form H2O2 in chloroplast and mitochondria (Elstner, 1991). However H2O2 may also be involved for the formation of highly reactive hydroxyl radicals by reacting with superoxide radicals in plants cells (Prousek, 2007) which can initiate self- replicating reactions leading to peroxidation of membrane lipids and destruction of proteins, and ultimately cell death (Jaw and Ching, 1998). Here the level of H2O2 in plants gradually increased with increase in stress level, however the results indicated that treated plants of both varieties showed the reduced level of H2O2 as compared to their respective control, although the difference were not significant in all the treatments. In this study, irrespective of treatments and stress level, Sahbhagi showed the reduced (35.27%) level of H2O2 as compared to IR-64, which might be an attribute of a tolerance variety to cope with drought stress. It is believed that under drought stress, PGPRs treated plants showed the high amount of reactive oxygen species (ROS) scavenging agent which may help plants to maintain the reduced level of H2O2 (Moslemi et al., 2011).
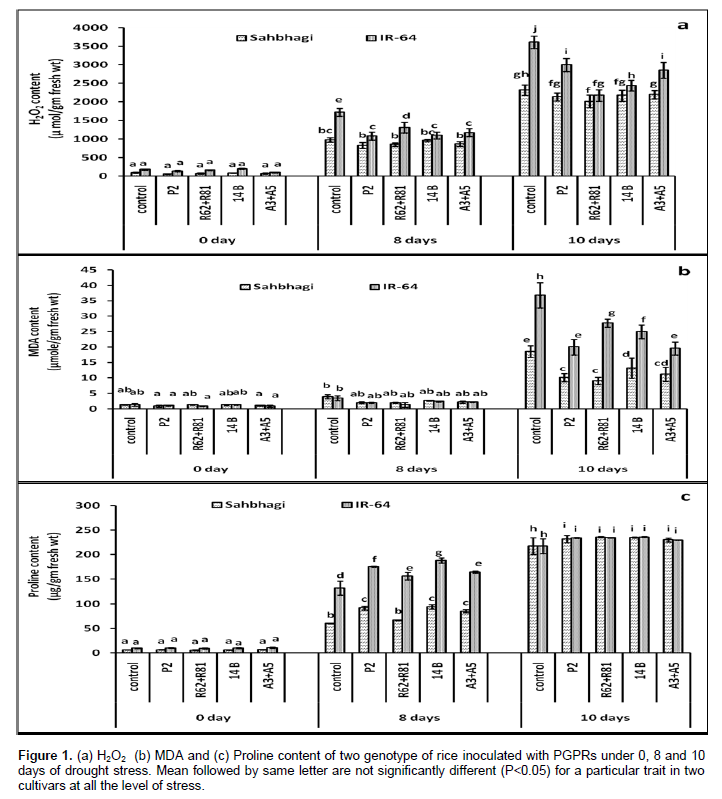
Malondialdehyde content
In 0 and 8th days of stress there was non-significant effect of the treatments over the control while at 10th days of stress all the bacterial treatment significantly reduced the MDA content when compared with control in both varieties (Figure 1b). MDA is one of the byproducts of lipid peroxidation, which is one of the consequences of higher accumulation of ROS such as H2O2, superoxide radical and hydroxyl radical in plant cell and could reflect the degree of peroxidation of membrane lipids (Gill and Tuteja, 2010). Here under severe drought both varieties showed higher MDA content in leaves which may be associated with higher accumulation of H2O2 in stressed plants. However the PGPRs treated plants in both varieties showed remarkably lesser amount of MDA content as compared to their respective control, suggesting the involvement of PGPRs in ROS metabolism in rice plants. In overall, irrespective of treatments and stress level sahbhagi performed better with 81.53% lesser MDA content as compared to IR-64. This might be an attribute of tolerant variety that helps to sustain it under prolonged environmental stress condition. Less MDA content in drought tolerance Phaseolus acutifolius as compared to sensitive ones have also been obtained by El-Tayeb (2006).
Proline accumulation
In plants with 0 days of stress, bacterial inoculation was non-significant over the control while in stressed, all the PGPRs treated plants except R62+R81 treated plants of Sahbhagi in 8th days of drought, showed the significant increase in proline contents in both of the varieties of rice at both the level of stress. In 10th days of stress, a maximum increase of 1.08 fold in proline content was observed in R62+R81 treated plants of Sahbhagi while in IR-64, a maximum increase in 1.09 fold was observed in 14B treated plants over control (Figure 1c). High proline contents in plant cells may be due to higher induction of proline biosynthesis which may help cells to maintain their water status and protects their vital function against the consequences of drought stress (Yoshiba et al., 1997). Higher proline accumulation in inoculated plants may indicate higher plant tolerance to water stress. Irrespective of treatments, proline accumulation was higher (105.43%) in IR-64 as compare to Sahbhagi in early drought stress (in 8th days of drought), while in severe drought stress both the varieties accumulate proline in almost similar ways, the attribute might differentiate the tolerance and sensitive varieties of rice and suggesting the vital role of proline as an osmoregulatory solute in plants (Kumar et al., 2011). Similarly in various studies on abiotic stresses, plants showed the varietal differences in proline accumulation (Yadav et al., 2004; Liu et al., 2011).
Superoxide dismutase activity
The SOD activity were continued to increase till the last day of drought (10th days of drought). On the 8th day of drought, P2 showed 1.76 fold increased SOD over control in Sahbhagi, followed by A3+A5 with 1.36 fold increase. In IR-64, Arthrobacter A3+A5 increased the 1.78 fold SOD activity over control followed by R62+R81 with 1.65 fold. On the 10th days of drought, A3+A5 with 1.93 fold followed by P2 with 1.65 fold maximally increased the SOD activity over control in Sahbhagi. In IR-64 all the treated plants showed significantly higher activity of SOD as compared to control plants in 10th days of drought. Among the treatments, R62+R81 with 1.75 fold followed by Arthrobacter A3+A5 with 1.72 fold maximally increased the SOD activity over control (Figure 2a). It is suppose that SOD catalyses the dismutation of superoxide radical into H2O2, which is further obliterate by CAT and POD activity (Scandalios, 1993). Here the increased SOD activity in stressed plants may be either due to increased production of ROS or could be a protective mechanism adopted by rice plants against ROS and oxidative damage. The higher level of SOD activity in inoculated plants could be related with the enhanced protective mechanism induced by PGPRs in plants to reduce the level of H2O2. An earlier study demonstrated that mechanisms that reduce oxidative stress indirectly play an important role in drought tolerance (Bowler et al., 1992).
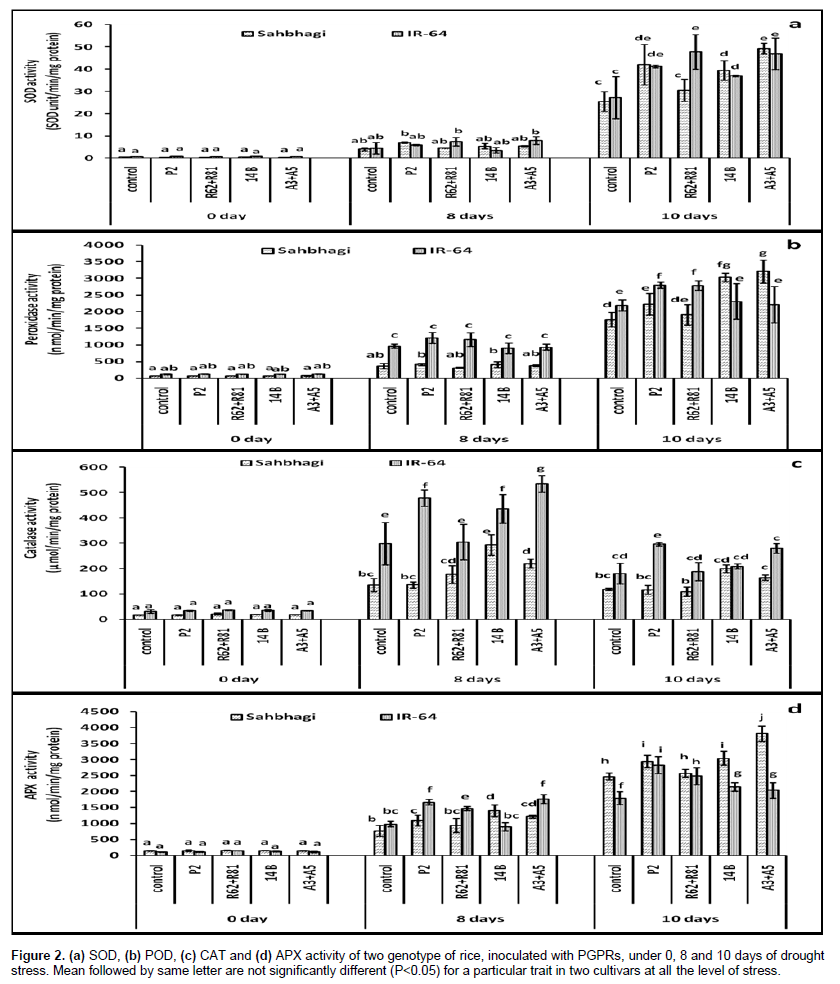
Peroxidase activity
Sahbhagi strain P2 with 1.39 fold maximally increased the POD activity followed by 14B with 1.34 fold which increased activity under 8th days of drought. Similarly in IR-64 with 1.25 fold, Pseudomonas strain P2 maximally increased the POD activity followed by R62+R81 with 1.2 fold. In 10th days of drought Arthrobacter (with 1.81 fold), 14B (with 1.71 fold) and P2 (with 1.26 fold) significantly increased the POD activity over control in Sahbhagi, while IR-64 Pseudomonas strain P2 (with 1.27 fold) and R62+R81(with 1.27 fold) showed significant effect on POD activity over control (Figure 2b). Irrespective of the treatments, IR-64 showed 25.72% higher POD activity as compared to Sahbhagi. Under stress condition, increased level of peroxidase in plants can be correlated with an increased level of ROS in plants cells (Radotic et al., 2000). Similar to the present study, PGPRs mediated increased POD activity under drought stress has also been reported in green gram plants by Saravanakumar et al. (2011).
Catalase activity
The catalase activity increased in the 8th day of drought and then decreased under severe drought stress in 10th day. A non significant effect was observed in all the treatments with control plants under 0 days of stress. In 8th days of drought 14B and A3+A5 significantly increased the CAT activity in Sahbhagi, while IR-64, P2, 14B and A3+A5 significantly increased the catalase activity over control. In the 10th days of drought, 14B with 1.67 fold maximally increased the catalase activity over control in Sahbhagi. In IR-64, P2 with 1.64 fold significantly increased the CAT activity as compared to uninoculated plants (Figure 2c). Irrespective of treat-ments and stress level 91.25% increased CAT activity was reported in IR-64 as compared to Sahbhagi. Here the catalase activity increased at the 8th day of drought and decreased under severe stress condition after 10th days of drought which either suggest catalase poor affinity for H2O2 or it may have undergone subsequent degradation of H2O2 due to photoinactivation in the presence of light (Hertwig et al., 1992). Similar decline in catalase activity in plants has also been observed in varieties under stressful condition (Hertwig et al., 1992; Radotic et al., 2000). However, all the inoculated plants showed higher catalase activity as compared to their respective control which suggests the PGPR mediated reduction of oxidative stress in plants (Saravanakumar et al., 2011).
Ascorbate peroxidase activity
In the 8th day of drought 14B (with 1.8 fold), A3+A5 (with 1.58 fold) and pseudomonas strain P2 (with1.41 fold) significantly increased the APX activity over control in Sahbhagi. In IR-64, Arthrobacter (with 1.79 fold) and pseudomonas strain P2 (with 1.69 fold) showed significant effect on APX activity over control. Similarly in the 10th day of drought 14B (with 1.23 fold) Arthrobacter (with 1.54 fold) and pseudomonas strain P2 (with1.19 fold) significantly increased the APX activity over control in Sahbhagi. In IR-64 all the treated plants showed significantly increased level of APX activity over control (Figure 2d). Overall (irrespective of treatments and stress level), Sahbhagi showed 12.25% higher APX activity as compared to IR-64. APX acted on H2O2 and prevents its accumulation in cells via ascorbate-glutathione pathway (Foyer and Halliwell, 1976). An increased activity of APX in PGPRs treated plants as observed here could be related with the decreased concentration of H2O2 in rice under drought stress suggesting a key role of APX in detoxification of H2O2 under drought stress and appear to constitute a basic antioxidative defense mechanism in plants (Madhusudhan et al., 2003). However, in the overall antioxidant study, higher activity of CAT, POD and APX could interrelate with lower activities of H2O2 in all the treated plants as compared to their respective control, as all these antioxidant acted as an scavenger on H2O2 (Gill and Tuteza, 2010).
CONCLUSION
It was observed from the present study that PGPRs inoculation induced plants to produce the higher amount of antioxidant under drought stress which might be a basis for the lower accumulation of H2O2 in inoculated plants as compared to their respective control. Both the genotype differs in their response to different growth and biochemical parameters under drought stress condition, however, lower accumulation of H2O2 in Sahbhagi indicated that this cultivar might have an efficient ROS quenching system at cellular level, which might help it to withstand prolonged drought. Overall, the study shows the significance of PGPRs as the alleviation of drought stress in rice and suggest the further utilization of selected PGPRs as biofertilizer under drought prone environment.
CONFLICT OF INTEREST
The authors declared they have no conflict of interest.
ACKNOWLEDGEMENTS
The first author is very thankful to the International Rice Research Institute (IRRI), Philippine for providing financial support through the STRASA project.
REFERENCES
|
Alami Y, Achouak W, Marol C, Heulin T (2000). Rhizosphere soil aggregation and plant growth promotion of sunflowers by exopolysaccharide producing Rhizobium sp. strain isolated from sunflower roots. Appl. Environ. Microbiol.66:3393-3398. |
|
|
Alexieva V, Sergiev I, Mapelli S, Karanov E (2001). The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 24:1337-1344. |
|
|
Arnon DI (1949). Copper enzymes in isolated chloroplast polyphenol oxidase in Beta vulgaris. Plant physiol. 24:1-15. |
|
|
Bates LS, Waldren RP, Teare ID (1973). Rapid determination of free proline for water–stress studies. Plant Soil.39:205-207. |
|
|
Bernier J, Atlin GN, Serraj R, Kumar A, Spaner D (2008). Breeding upland rice for drought resistance. J. Sci. Food Agric.88: 927-39. |
|
|
Bowler C, Vanmontagu M, Inze D (1992). Superoxide-dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43:83-116. |
|
|
Bradford MM (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:246–254. |
|
|
Cha-um S, Yooyongwech S, Supaibulwatana K (2012). Water-deficit tolerant classification in mutant lines of indica rice. Sci. Agric. 69(2):135-141. |
|
|
Egamberdiyeva D, Hoflich G (2004). Effect of plant growthpromoting bacteria on growth and nutrient uptake of cotton and pea in a semi-arid region of Uzbekistan. J. Arid. Environ. 56:293-301. |
|
|
Elstner EF (1991). Mechanism of oxygen activation in different compartments. in: Pell EJ, Steffen KL (Eds.), Active Oxygen/Oxidative Stress and Plant Metabolism. American Society of Plant Physiologists, Roseville. pp. 13-25. |
|
|
El-Tayeb MA (2006). Differential responses of pigments, lipid peroxidation, organic solutes, catalase and per-oxidase activity in the leaves of two Vicia faba L. cultivars to drought. J. Agri. Biol. 8(1): 116-122. |
|
|
Foyer CH, Halliwell B (1976). The presence of glutathione and glutathione reductase in chloroplasts: a proposed role in ascorbic acid metabolism. Planta. 133:21-25. |
|
|
Gill SS, Tuteja N (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48:909-930. |
|
|
Glick BR (1995). The enhancement of plant growth by free living bacteria. Can. J. Microbiol. 41:109-114. |
|
|
Gururani MA, Upadhyaya CP, Venkatesh J, Baskar V, Park SW (2013). Plant growth promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS scavenging enzymes and improved photosynthetic performance. J. Plant Growth Regul. 32(2): 245-258. |
|
|
Gusain YS, Kamal R, Mehta CM, Singh US, Sharma AK (2015). Phosphate solubilizing and indole-3-acetic acid producing bacteria from the soil of Garhwal Himalaya aimed to improve the growth of rice. J. Environ. Biol. 36:301-307. |
|
|
Halliwell B, Gutteridge JMC (1989). Free Radicals in Biology and Medicine (2nd ed) Clarendon Press, Oxford. |
|
|
Heath RL, Packer L (1968). Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophysic. 125(1):189-198. |
|
|
Hertwig B, Streb P, Feierabend J (1992). Light dependency of catalase synthesis and degradation in leaves and the influence of interfering stress conditions. Plant Physiol. 100:1547-1553. |
|
|
Hoagland DR, Arnon DI (1950). The water-culture method for growing plants without soil. Calif. Agr. Expt. Sta. Circ. 347:1-32. |
|
|
Jaw NL, Ching HK (1998). Effect of oxidative stress caused by Hydrogen peroxide on senescence of rice leaves. Bot. Bull. Acad. Sin. 39:161-165. |
|
|
Kamoshita A, Babu RC, Boopathi NM, Fukai S (2008). Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res. 109:1-23. |
|
|
Khush G (2003). Productivity improvements in rice. Nutr. Rev. 61:S114-S116. |
|
|
Kohler J, Hernandez JA, Caravaca F, Roldàn A (2008). Plant-growth-promoting rhizobacteria and abuscular mycorrhizal fungi modify alleviation biochemical mechanisms in water-stressed plants. Funct. Plant Biol. 35:141-151. |
|
|
Kumar RR, Karajol K, Naik GR (2011). Effect of polyethylene glycol induced water stress on physiological and biochemical responses in pigeonpea (Cajanus cajan L. Millsp.). Recent Res. Sci. Tech. 3:148-152. |
|
|
Liu C, Liu Y, Guo K, Dayong Fan D, Li G, Zheng Y, Yu L, Yang R (2011). Effect of drought on pigments, osmotic djustment and antioxidant enzymes in six woody plant species in karst habitats of south western China. Environ. Exp. Bot. 71:174-183. |
|
|
Mackill DJ, Ismail AM, Pamplona AM, Sanchez DL, Carandang JJ, Septiningsih EM (2010). Stress Tolerant Rice Varieties for Adaptation to a Changing Climate. Crop Environment & Bioinformatics.7:250-259. |
|
|
Mader P, Kaiser F, Adholeya A, Singh R, Uppal HS, Sharma AK, Srivastava R, Sahai V, Aragno M, Wiemken A, Johri BN, Fried PM (2011). Inoculation of root microorganisms for sustainable wheat-rice and wheat black gram rotations in India. Soil Biol. Biochem. 43:609-619. |
|
|
Madhusudhan R, Ishikawa T, Sawa Y, Shiqeoka S, Shibata H (2003). Characterization of an ascorbate peroxidase in plastids of tobacco BY-2 cells. Physiol. Plant. 117:550-557. |
|
|
Mayak S, Tirosh T, Glick BR (2004). Plant growth promoting bacteria that confer resistance to water stress in tomato and pepper. Plant Sci. 166:525-530. |
|
|
Mishra S, Jha AB, Dubey RS (2011). Arsenite treatment induces oxidative stress, upregulates antioxidant system, and causes phytochelatin synthesis in rice seedlings. Protoplasma 248(3): 565–577. |
|
|
Mongodin EF, Shapir N, Daugherty SC, De Boy RT, Emerson JB, Shvartzbeyn A, Raduneet D, Vamathevan J, Riggs F, Grinberg V, Khouri H, Wackett LP, Nelson KE, Sadowsky MJ (2006). Secrets of soil survival revealed by the genome sequence of Arthrobacter aurescens TC1. PLoS Genetics 2(12):2094-2106. |
|
|
Moslemi Z, habibi D, Asgharezadeh A, Ardakani MR, Mohammadi A, Mohammadi M (2011). Response of phytohormones and biochemical markers of maize to super absorbent polymer and plant growth promoting rhizobacteria under drought stress. American Eurasian J. Agric.& Environ. Sci. 10 (5):787-796. |
|
|
Noctor G, Foyer CH (1998). Ascorbate and glutathione: keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49:249-279. |
|
|
Prousek J (2007). Fenton chemistry in biology and medicine. Pure Appl. Chem. 79(12):2325-2338. |
|
|
Radotic K, Ducic T, Mutavdzic D (2000). Changes in peroxidase activity and isozymes in spruce needles after exposure to different concentrations of cadmium. Environ. Exp. Bot. 44:105-113. |
|
|
Roesti D, Gaur R, Johri BN, Imfeld G, Sharma S, Kawaljeet K, Aragno M(2006). Plant growth stage, fertiliser management and bio-inoculation of arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria affect the rhizobacterial community structure in rain-fed wheat fields. . Soil Biol. Biochem. 38:1111-1120. |
|
|
Sairam RK, Srivastava GS (2001). Water stress tolerance of wheat (Triticum aestivum L.): Variations in hydrogen peroxide accumulation and antioxidant activity in tolerant and susceptible genotypes. J. Agron. Crop Sci. 186:63-70. |
|
|
Sandhya V, Ali SKZ, Grover M, Reddy G, Venkateswarlu B (2010). Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth. Regul. 62:21-30. |
|
|
Saravanakumar D, Kavino M, Raguchander T, Subbian P, Samiyappan R (2011). Plant growth promoting bacteria enhance water stress resistance in green gram plants. Acta Physiol. Plant. 33:203-209. |
|
|
Saravanakumar D, Samiyappan R (2007). ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 102:1283-1292. |
|
|
Scandalios JG (1993). Oxygen stress and superoxide dismutases. Plant Physiol. 101:7-12. |
|
|
Sgherri CLM, Pinzino C, Navari-Izzo F (1996). Sunflowers seedlings subjected to stress by water deficit: changes in O2 production related to the composition of thylakoid membranes. Physiol. Plant. 96:446-452. |
|
|
Singh KK, Ghosh S (2013). Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L.) cultivars under water deficit conditions. Plant Cell Reports 32(2):183-193. |
|
|
Smirnoff N (1993). The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 125:27-58. |
|
|
Yadav RS, Sharma RK, Yadav RM (2004). Effect of simulated drought on free proline accumulation in some wheat (Triticum aestivum L.) genotypes. Indian J. Agric. Biochem. 17:43-44. |
|
|
Yoshiba Y, Kiyosue T, Nakashima K, Yamaguchi-Shinozaki K, Shinozaki K(1997). Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 38:1095-1102. |
|
|
Zhang J, Kirkham MB (1996). Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytologist 132:361-373. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


