Full Length Research Paper

ABSTRACT
The importance of extracellular enzyme producing endophytic bacteria and their ability to elicit ‘induced systemic tolerance’ against abiotic stress (sodic soil) has been documented. However, the performance of these microbes under various abiotic stresses, especially saline-sodic conditions is less understood and formed the focus of this investigation. The diversity of endophytic bacteria associated with leaves, stems and roots of sodicity tolerant polyembryonic mango root stock (GPL-1 and ML-2), grown at the sodic soil experiment farm (shivery farm), Central Soil Salinity Research Institute, Regional Research Station, Lucknow, India was investigated. In this study, we isolated 16 bacterial endophytes through natural selection from saline sodic soils, analysed extracellular enzyme activity, performed molecular profiling and phylogenetic analysis based on 16S rDNA sequences. Results indicate that the isolates belonged to four major phylogenetic groups: low G+C Gram positive bacteria, Firmicutes, Proteobacteria and Bacteroidetes. Endophytic bacteria from the phylum Firmicutes were predominant in the root portion 60.0% (Na+ - 7.72 ± 0.05; K+- 2.08 ± 0.85) and stem portion 75.0% (Na+ - 5.79 ± 0.05; K+- 13.58 ± 0.53), respectively. Most isolates that exhibited extracellular enzymatic activity in 3.0 M NaCl concentration belonged to the genus Bacillus sp., Bacillus clausii, Bacillus pumilus and Bacillus licheniformis (CSR-M-06, CSR-M-08, CSR-M-09 and CSR-M-16) exhibited the stronger activities in extracellular enzymes such as amylase, protease, cellulase and lipase than other isolates.
Key words: Endophytic bacteria, sodicity tolerance, polyembryonic mango, extracellular enzyme activity.
INTRODUCTION
Mango (Mangifera indica L.), one of the 73 genera of the family Anacardiaceae in order Sapindales, is amongst the most important tropical fruits of the world. It is more sensitive to salinity at early growth stages which led to identification of salt tolerant true to type polyembryony rootstocks ML-2, GPL-1 and ML-6 (Damodaran et al., 2013). Tolerance to sodicity in salt sensitive crops like gladioulus and banana may be epigenetic or may be through a response of the plant to the endophytic population that reside in the root zone of the crop and produce stress tolerant enzymes like superoxy dismutase (SOD), catalase, peroxidise, polyphenol oxidase and other phenol compounds (Damodaran et al., 2014). Bacterial endophytic species are common inhabitants of a wide range of plant species and reside within cells (Jacobs et al., 1985), in the intercellular space (Patriquin and Dobereiner, 1978), or in vascular systems (Bell et al., 1995) of a plant. Bacterial endophytes seem to be ubiquitous in plant tissues, having been isolated from flowers, fruits, leaves, stems, roots and seeds of various plant species (Kobayashi and Palumbo, 2000). Signi-ficant variations in the populations of both indigenous and introduced endophytes have been reported. These variations are attributed to plant source, plant age, tissue type and environment (Zinniel et al., 2002). The potential to use salt tolerant endophytes to remediate salt polluted soils has attracted considerable interest (Idris et al., 2004). Bacterial endophytes can stimulate contaminant disappearance by the accumulation and transformation of sodium (Na+).
Moore et al. (2006) describes the diversity of endophytes found in poplar trees, growing at a phytoremediation field site contaminated with toluene. Germaine et al. (2004) demonstrated that within the diverse bacterial communities found in poplar trees several endophytic strains were present that had the potential to enhance phyto-remediation of volatile organics and herbicides. For this, several bacterial species possess genes responsible for resistance to salt and have evolved a variety of mechanisms to reduce heavy metals stress (Alonso et al., 2000; Van Houdt et al., 2009; Khan et al., 2009).
Recently, production of enzymes such as amylase, proteases, cellulose and lipase by salt tolerant endophytic bacteria under adverse environments has become a major field of investigation (Alqueres et al., 2007). This interest is due to the possibility of these microorganism producing enzymes with biotechnologically potential antifungal agents (Cryptosporiopsis cryptocandina, quercine), besides producing factors of plant growth, toxins and enzymes (Araújo et al., 2002; Strobel, 2003; Azevedo, 1998; Stierle et al., 1993). Several reports confirmed that the activity of salt tolerant bacterial enzymes remained stable at high salt concentrations (Prakash and Jaiswal, 2009). However, extracellular enzymes like amylase, lipase, cellulose and protease are produced by different microorganisms via Bacillus subtilis, Bacillus stearothermophilus, Bacillus licheniformis, and Bacillus amyloliquefaciens, Pseudomonas (Devi et al., 2010; Bassabrani et al., 2012), Halobacillus sp. (Amoozegar et al., 2003) and Bacillus dipsosauri (Deutch, 2002) but production of this enzyme by salt tolerant plant bacterial endophytic strains. Previous studies on the occurrence of endophytic bacteria in sodicity tolerant polyembryonic mango root stock or other parts of the plants, on sodium uptake pattern and PGPR traits of microorganisms have been conducted (Damodaran et al., 2013; Kannan et al., 2014). However, there are no reports on the occurrence and diversity of extracellular enzyme producing endophytic bacteria in sodicity tolerant polyembryonic mango root stock grown in salt affected sodic soils.
MATERIALS AND METHODS
Sites and isolation of endophytic bacteria
Endophytic bacteria were isolated from surface-sterilized sodicity tolerant polyembyonic mango root stock ML-2 and GPL-1 (the plants can grow normally at the level of pH-9.53 and electrical conductivity EC- < 4.0). The rhizosphere soil total Na+ and K+ concentrations of GPL-1 and ML-2 were 21.20 meq / L; and 0.126 meq / L respectively. Two tolerant polyembryonic mango plants were collected from the experimental farm of Central Soil Salinity Research Institute Regional Research Station, Lucknow, India, located at 260 47?45?? to 260 48?13?? N latitude and 800 46?7?? to 800 46?32?? E longitude, lying in the Central part of Indo-Gangetic plain, in June 2012. Roots, stems and leaves from each plant were separated and washed extensively first in several changes of 0.01 M EDTA and then in distilled water to remove any non-specifically bound Na+ / K+ and dried to constant weight. Dried plant tissues were ground into a fine power, and digested with HNO3:H2SO4:HClO4 (10:1:4). Digested material was diluted to 10 ml and the sodium (Na+) / potassium (K+) content was analyzed with the help of a flame photometer (Furman, 1975). Plant samples were washed with tap water followed by three rinses with deionized water and then separated into roots, stems and leaves. Healthy plant tissues were sterilized by sequential immersion in 75% (v/v) ethanol for 2 min, and 1% mercuric chloride for 1 min and washed three times with sterile water to remove surface sterilization agents. To confirm the surface disinfection process was successful, water from the final rinse was plated on Nutrient agar. Plant tissue materials (0.2 g) were ground by a mortar and pestle in the presence of 5 ml of sterile distilled water. Serial dilutions were spread on plates containing 0.1 to 3.0 M range NaCl3 supplement of nutrient agar medium with or without NaCl3. The medium was found to be most suitable for the isolation of sodium tolerant bacteria in preliminary experiments. NaCl3 resistant colonies were picked randomly and purified by streaking three to four times on the same media. Sixteen endophytic bacterial isolates growing well on sub culturing were finally selected and stored on slants for further study.
Analysis of extracellular enzyme activity
Endophytic bacterial isolates were analysed for production of four enzymes under (0.1, 0.5, 1.0, 1.5, 2.0, 2.5 and 3.0 M) NaCl stress, that is, protease, amylase, cellulase and lipase by plate methods. Proteolytic activities of the cultures were studied in a medium containing skimmed milk (HiMedia, Mumbai). Zones of precipitation of paracasein around the colonies in the next 48 h were taken as evidence of proteolytic activity. The presence of amylolytic activity was determined using starch agar medium (HiMedia, Mumbai). After inoculation of endophytic bacteria and incubation at 28°C for 5 days, the plates were flooded with 0.3% I2 and 0.6% KI solution and a clear zone around the growth indicated hydrolysis of starch. For cellulase activity, mineral-salt agar plate 5 containing 0.4% (NH4)2SO4, 0.6% NaCl, 0.1% K2HPO4, 0.01% MgSO4, 0.01% CaCl2 with 0.5% carboxymethylcellulose and 2% agar (HiMedia, Mumbai) were surface inoculated. Iodine solution was used to detect cellulase activity as described.
Growth pattern of effective isolates at different NaCl concentrations
Growth of salt tolerant bacteria was measured at different NaCl concentration (0.1, 0.5, 1.0, 1.5, 2.0, 2.5, and 3.0 M). L-Broth having different NaCl concentrations was prepared and autoclaved. For preparation of inoculum, bacterial cells harvested from overnight culture were washed and resuspended in sterile distilled water to adjust the OD at 106 cells per ml at 600 nm. 100 μl of inoculum was given in all media having different NaCl concentrations and incubated at 37°C on shaker with 150 rpm shaking for 24 h. Bacterial growth was monitored at 600 nm (Spectrophotometer S200, R & M Marketing, England). Three replicates per strain were taken for each salt concentration.
Vigour index studies
Plant growth promoting (PGP) activities of the current strains were assessed using seedling vigour index by pot culture studies in saline sodic soils of pH 9.40, electrical conductivity (ECe) 6.72 dSm-2; sodium (Na+) 21.20 meq / L. The vigor index was calculated by using the formula as described by Abdul-Baki and Anderson (1973).
Vigor index= Percent germination x Seedling length (shoot length + root length)
Molecular profile of endophytic bacterial
Bacterial DNA from the elite strains showing higher extra cellular activities were extracted according to the method given by Araujo et al. (2002), with amplification of 16S rDNA being performed in a 25 μl reaction mixture, which consisted of 25 ng genomic DNA, 10× reaction buffer with 15 mM MgCl2, 2.5 mM each of dNTPs, 0.2 mM bacterial universal primer (27F-AGAGTTTGATCMTGGCTCAG) (1492R-GGYTACCTTGTTACGACTT), and 1 unit of Taq DNA polymerase (Bangalore Genei Pvt Ltd, India). The reaction was carried out in a thermal cycler (PTC-200, MJ Research Inc, USA). The DNA amplification program was set to a 5 min initial denaturation at 94°C followed by 30 cycles of 1 min at 94°C for denaturation, 1 min at 54°C for annealing, 2 min at 72°C for extension, and ended with a final 10 min extension at 72°C. The reaction products were separated by running 5 μl of the polymerase chain reaction mixture in 1.2% (w/v) agarose gel and staining the bands with ethidium bromide (Sambrook et al., 1989). The Restriction Fragment Length Polymorphism (RFLP) assay involved restriction enzyme digests and the 16S rRNA gene PCR products using five different enzymes. Reactions were performed in a 30 μl reaction mixture, which consisted of 10 μl of each PCR product, 2 μl of 10× Fast digest green buffer, 17 μl nuclease free water and 1 μl (1FDU) for restriction enzyme cleavage by either AluI, Hae III, EcoRI, MspI or BamHI (Fermentas, USA). Digestions were performed for 5 min at 37°C to ensure that complete fragmentation was achieved. Digested DNA fragments were analysed in a 1.5% (w/v) agarose gel with 10 mg / ml of ethidium bromide and photographed on a gel documentation system. For cluster analysis, the data were converted to a binary matrix, where the digit 1 or 0 represented the presence of or absence of DNA band in the gel. NTSYSpc 2.20 (Exeter software, New York) was used to analyze the fingerprinting. The data were analysed using the neighbor-joining tree construction method to generate the Dice similarity coefficient. Similarity coefficients were used to construct the dendrogram using the Dice coefficient in clustering (Damodaran et al., 2012). Statistical stability of the branches in the cluster was estimated by bootstrap analysis using Winboot software program (Yap and Nelson, 1996). The principle co-ordinate analysis was performed with NTSYS software with the Dice (1945) coefficient.
Sequencing and phylogenetic analysis
Purified PCR products from the 16S rRNA genes of pure culture isolates representing all major RFLP patterns were sequenced on an ABI3730 automated sequencer (Applied Biosystems, USA) with primers 27 f and 1492 r. All reference sequences were obtained from the National Center for Biotechnology Information (NCBI) and the 16S rDNA similarity sequences searches were performed using the BLASTN tool in the NCBI website. These sequences were aligned using the multiple sequence alignment program, CLUSTAL X (Thompson, 1996). The method of Jukes and Cantor (1969) was used to calculate evolutionary distances. A phylogenetic dendogram was constructed by the neighbor-joining method and tree topologies were evaluated by performing bootstrap analysis of 1,000 data sets using MEGA 3.1 (Molecular Evolutionary Genetic Analysis).
Nucleotide sequence accession numbers
The sequence obtained in this study was deposited in the GeneBank nucleotide sequence database under the accession numbers (KC433665 and KC433667).
RESULTS
Na+, K+ partitions and bacterial endophytes in different parts of sodicity tolerant mango root stock
Na+ and K+ concentration in plant tissues are shown in Table 1. Na+ concentration in the roots were significantly higher than that of stem and leaves and K+ concentration of root parts were significantly lower than that of other parts. There were significant differences between the Na+ and K+ concentrations in the roots of salt tolerant polyembryonic mango root stock (GP-1 and ML-2) and salt sensitive polyembryonic mango root stock (GTP-2). It is worth noting that GPL-1 and ML-2 polyembryonic root stock accumulated significantly higher Na+ and lower accumulation K+ in its (7.72 ± 0.05 meq / l 2.08 ± 0.85 meq / l) dry weight. A total of forty NaCl-resistant-endophytic isolates were obtained, from GPL-1 and ML-2 root stock. Endophytic bacterial counts of the two sodicity tolerant polyembryonic mango root stock (GPL-1 and ML-2) ranged from 4.4 × 104 to 9.6 × 104 fresh weight (Tables 1 and 2). It was very noticeable that NaCl-resistant-endophytic bacterial from GPL-1 root stock attained 104 cfu g-1 with fresh weight of 2.6 and ML-2 root stock with fresh weight of 2.31 ± 0.20 × 104 cfu g-1, respectively. It was also observed that endophytic NaCl tolerant bacterial were nil or 0.00 in stem and leaves of the GPL-1, ML-2 root stock.

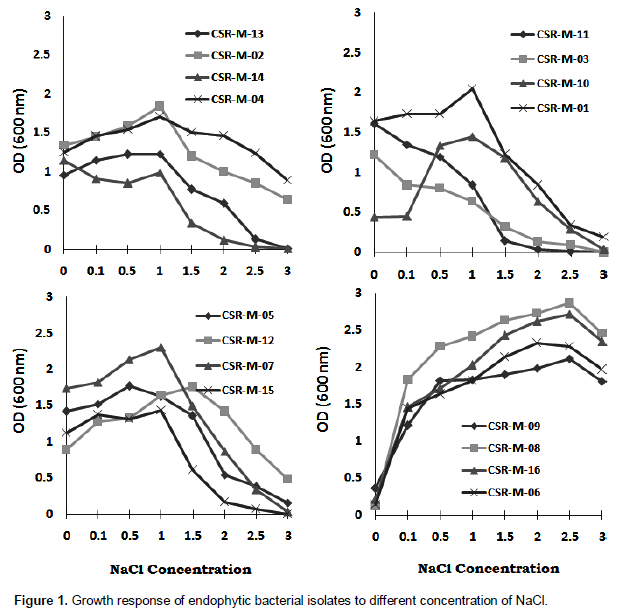
In-vitro growth response of endophytes with effect on different concentration of NaCl
Endophyte isolates were grown in the presence of different concentrations (0.1 to 3.0 M NaCl in L-Broth) of NaCl is presented in Figure 1. All the isolates were able to grow in medium containing 1.0 M and more NaCl. They could tolerate 2.0 to 3.0 M NaCl in solid medium and 2.5 to 3.0 M NaCl in the liquid medium (L-Broth). However, growth was less at 2.0 and 2.5 M NaCl. In liquid medium most of the isolates could respond up to 3.0 M NaCl while some could grow up to the range of 2.5 (CSR-M-08, CSR-M-09, CSR-M-06, CSR-M-16) and 3.0 M (CSR-M-06, CSR-M-08, CSRM-09, CSR-M-16).
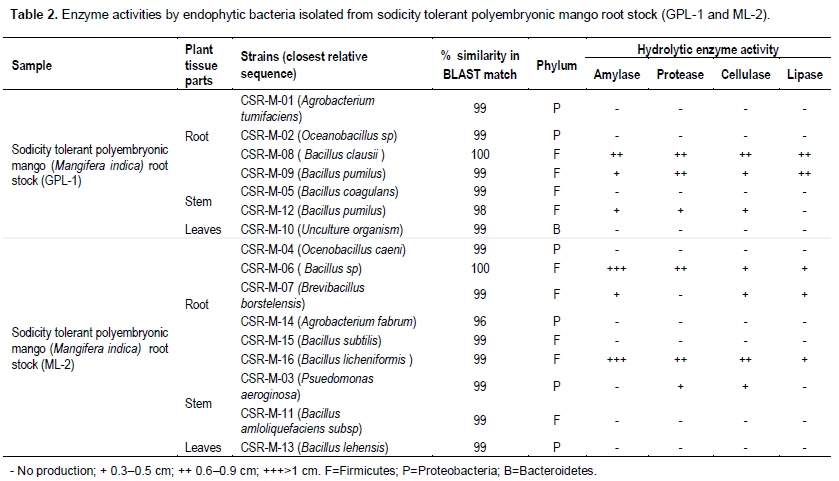
Extracellular enzyme production by bacterial endophytes under different concentration of NaCl
Sixteen (16) newly isolated endophytic bacterial were screened for production of extracellular enzymes. The method of radial diffusion in solid media with supplement of 0.1 to 3.0 M of sodium chloride concentration indicated that actively hydrolysing enzymes with higher salt concentration could be detected in qualitative form by direct correlation of the diameter of the zone of substrate hydrolysis and colony growth. This practical tool facilitates and speeds the selection and comparison of the enzymatic production of different isolates. The enzyme biosynthesis capacity of isolated bacterial endophytes was shown in Table 2. Four isolates (CSR-M-08, CSR-M-09, CSR-M-06 and CSR-M-16) showed positive hydrolytic activity from sodicity tolerant polyem-bryonic mango root stock (GPL-1 and ML-2), two isolates from root of GPL-1 root stock (CSR-M-08 and CSR-M-09) and the other two isolates (CSR-M-06, CSR-M-16) from root of ML-2 mango root stock. The highest amylase, protease, cellulase and lipase activity was recorded from bacterial endophytes isolated from the root portion of both GPL-1 and ML-2. The zones of substrate hydrolysis ranged from 0.3 to >1.0 cm, respectively, and among the four most hydrolytic isolates, amylase production was found to be superior.
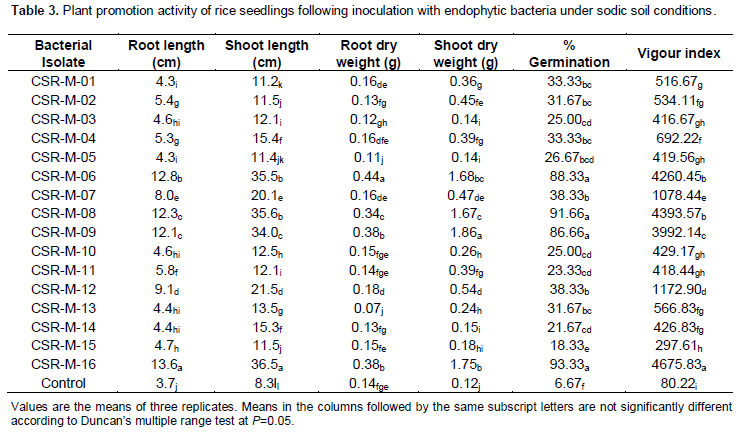
Screening of bacterial isolates for rice seedling vigour under sodic soil
Among the 16 isolates tested for their efficacy to improve the vigour index of rice seedlings under sodic soils (Table 3), interestingly, CSR-M-16 isolate showed the highest germination of 93.33% with a vigour index of 4675.83 while the vigor index of CSR-M-8, CSR-M-06 and CSR-M-09 isolates were on par (4393.57, 4260.45 and 3992.14, respectively) and these also showed range of 86.33 to 91.66% germination. Apart from this the isolates CSR-M-8, CSR-M-06 and CSR-M-09 also showed higher germination percentage of >86% and vigor index. The root length of 30 days old seedling for all treatment ranged from 4.3 to 13.6 cm, while higher value (13.6, 12.8, 12.3 and 12.1 cm) was recorded in seedling of CSR-M-16, CSR-M-06, CSR-M-08 and CSR-M-09. The lowest was found in control seedling (3.7 cm). Shoot length significantly higher (36.5, 35.6, 35.5 and 34.0 cm) was observed in seedling of CSR-M-16, CSR-M-08, CSR-M-06 and CSR-M-09. The smallest length (8.3 cm) was found in control seedling. Root dry weight ranged from length 0.07 to 0.44 g where higher (0.44, 0.38, 0.38, 0.34 g) root weight was found in seedling of CSR-M-06, CSR-M-09, CSR-M-16 and CSR-M-08 treatment. The lowest was observed in seedling of CSR-M-13 (0.07 g) treatment. Similarly significantly higher shoot dry weight (1.86, 1.75, 1.68 and 1.67 g) was found in seedling of CSR-M-09, CSR-M-16, CSR-M-06 and CSR-M-08 treatment.
PCR-RFLP finger printing
Endophytic bacterial isolates obtained from the rhizosphere of the salt tolerant mango rootstocks were found to be gram-negative predominantly. Of the 40 isolates discovered, 16 were used to assess genetic diversity by PCR-RFLP using 16S rRNA gene universal primer 27F and 1492R. The primers amplified a single band of 1.5 Kb in all the endophytic isolates and no inter or intraspecific variation was observed among the 16 endophyte strains. A set of five different restriction enzymes (AluI, HaeIII, EcoRI, MspI and BamHI) were used for RFLP analysis. The banding pattern generated with enzyme AluI, HaeIII and MspI showed greater polymorphism compared to the other enzymes and were chosen to construct the dendrogram and principle coordinate.
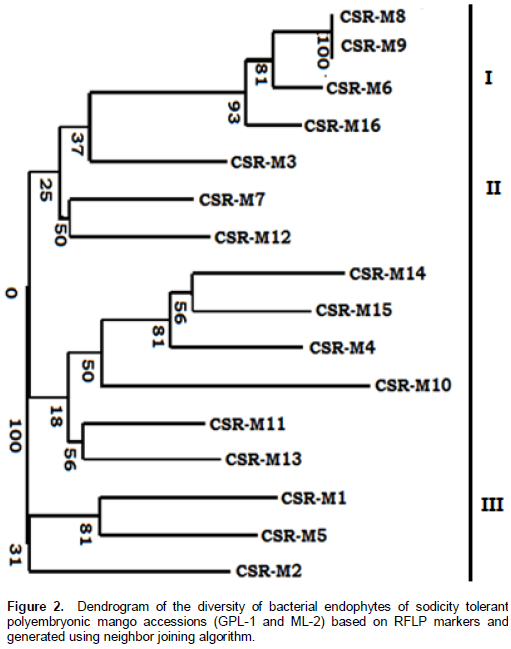
Cluster-based dendrogram
The dendrogram showing the genetic relation, extracellular enzyme activity under higher salt concentration among 16 endophytes based on RFLP markers is presented in Figure 2. The dendrogram showed three major clusters. The list of clusters along with the isolate name included in each cluster is also shown in Figure 2. Cluster I (primary cluster) consisted of four salt tolerant endophytes (CSR-M-08, CSR-M-09, CSR-M-06, CSR-M-16) growing above 2.5 M NaCl and positive for extracellular enzyme activity under higher salt concentration while the moderately salt tolerant isolates CSR-M-03, CSR-M-07, CSR-M-12 up to 1.5 M NaCl showed limited presence of extracellular enzymes like protease and cellulose. Cluster II and cluster III consisted of 12 endophytic bacterial isolates that are negative for extracellular enzyme activity and growth in salt concentration below 0.1 M NaCl. All the isolates of cluster I and II were isolated from roots of the salt tolerant mango rootstocks while the isolates showing limited response to NaCl toxicity are isolated from stem and leaves of the rootstocks.
Principle coordinate analysis
The positioning of 16 bacterial endophytes based on principle coordinate 1 and 2 is given in Figure 3. The scatter plot of Dice similarity showed three major groups of bacterial endophytes belonging to salt tolerant endophytes (group I) and those below 1.0 M NaCl concentration and negative for hydrolysis enzyme activity (group II, group III). Three bacterial endophytes (CSR-M-03, CSR-M-07, CSR-M-12) separated from the three groups. The principle coordinates 1 and 2 encompassed 59.42 and 38.96% of total variation, respectively.
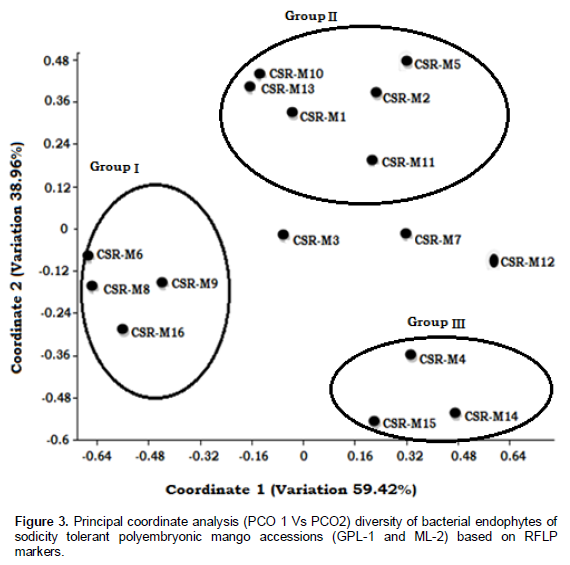
Confirmation of enzyme producing salt tolerant endophytes
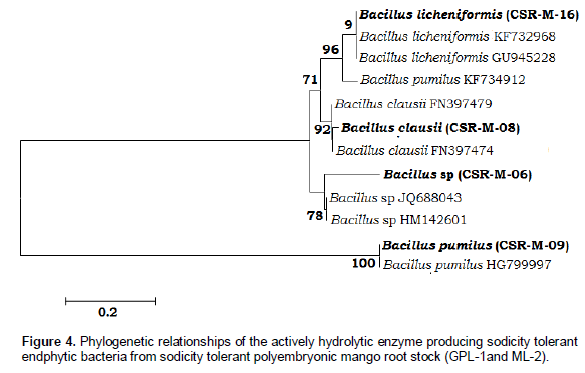
Identification of salt tolerant enzyme producing strains clustered in group I were verified by sequencing the 16S rRNA genes and validated with BLAST (ncbi.nlm.gov.blast/Blast.cgi) database. This method was carried out as follows: DNA was extracted from the isolates by a standard kit (Xcelgen-Germany). Then amplification of 16S rRNA gene (27F-AGAGTTTGATCCTGGCTCAG; 1492R-ACGGCTACCTTGTTACGACTT) was performed by PCR method and eventually the products were sequenced on an ABI 3730xl Genetic analyzer (Applied Biosystems, USA). The results (Figure 4) show that all of the potential strains belonged to the genus Bacillus, with the strain CSR-M-06 identified as Bacillus sp., CSR-M-08 as Bacillus clausii, CSR-M-09 as Bacillus pumilus and CSR-M-16 as B. licheniformis.
DISCUSSION
Although, special attention has been paid to endophytic bacterial isolated from sodicity tolerant polyembryonic mango root stock (GPL-1 and ML-2) in special environments such as sodium and heavy metal contaminated soils (Idris et al., 2004; Barzanti et al., 2007), no information is available about the composition of endophytic bacterial communities present in the two dominant sodium tolerant and accumulating plants naturally growing on salt affected wasteland. The surface sterilization protocol is a critical prerequisite for isolation and identification of the endophytic bacterial that express plant growth promotion (PGP) traits at high salt concentrations (Kannan et al., 2014). The present study showed that there was a reduction in the bacterial diversity with increased sodium concentration in plant tissue parts. It has been reported earlier that Na+ concentration in tissue parts plays a prominent role in the microbial adaption process as environmental stress leads to reduced bacterial diversity (Borneman et al., 1996). In our study, we have cultured 40 isolates from different tissue portions of polyembryonic mango accessions grown in sodic soils and identified 16 pure cultures for further investigations. The majority of endophytes are identified as Bacillus sp. based on 16S rDNA sequencing and validated with BLAST (ncbi.nlm.gov.blast/Blast.cgi) database. A similar process was adopted in many previous characterization studies (Sturz et al., 2000; Kuklinsky-Sobral et al., 2004; Tank and Saraf, 2010).
In this study, 16 bacterial endophytic strains were isolated from both sodicity tolerant polyembryonic mango root stock GPL-1 and ML-2. Out of the 16 endophytes, four (CSR-M-06, CSR-M-08, CSR-M-09 and CSR-M-16) showed tolerance to high (3.0 M) NaCl concentration (Figure 1). It has also been reported previously that bacterial isolated from saline soil are more likely to withstand saline conditions (Upadhyay et al., 2009). Among the isolates, four strains possessed amylase, cellulase, protease and lipase enzyme activity. According to the 16S rRNA sequences the strains belonged to genus Bacillus with CSR-M-06 identified as Bacillus sp, CSR-M-08 as B. clausii, CSR-M-09 as B. pumilus and CSR-M-16 as B. licheniformis which were able to produce above four enzymes under high NaCl concentrations. The characterized strains showed optimum growth between 2.5 to 3.0 M of NaCl. Therefore, according to the salt tolerance (up to 3.0 M) these strains could be described as salt tolerant microorganisms (Fahimeh et al., 2013; Dave et al., 2006). In general, the hydrolysis enzymes of endophytes appear to be important for the colonization of plant roots with external stress condition (Quadt-Hallmann et al., 1997; Reinhold-Hurek and Hurek, 1998; Sakiyama et al., 2001). This hypothesis is supported by the endophytic bacterial possessing higher activity of amylase, cellulase, protease and lipase enzymes from young radish roots (Weon Taek Seo et al., 2010). Similarly, the presence of cellulolytic and pectinolytic enzymes produced by numerous endophytic bacterial such as Rhizobium sp. has been described (Al-mallah et al., 1987). In addition, bacterial enter the interior of the root by hydrolyzing wall-bound cellulose, auxin-induced tumors, water flow and wounds, or where the lateral roots branch (Al-Mallah et al., 1987).
Molecular characterization based on amplification of 16s rDNA revealed the presence of B. clausii, B. pumilus, B. licheniformis and Bacillus sp. Variation of 16S rDNA allow the inference of phylogenetic relationship among taxonomically related as well as distinct organisms (Espinosa-Victoria et al., 2009). Based on Amplified Ribosomal Restriction Analysis (ARDRA), different groups were obtained in the dendrogram and these results find support from the observations made by Chowdhury et al. (2007) who divided the bacterial into various groups based on ARDRA pattern and one representative from each of the 4 groups was selected for sequencing of a 16S rRNA gene fragment so as to compare sequence similarity. Kannan et al. (2014) revealed that apart from normally encountered endophytic microflora, Bacillus coagulans, Bacillus megatarium, B. pumilus and B. subtilis were encountered. Molecular characterized phyto-ameliorant grasses grown on the patches of undisturbed sodic soil with prominent rhizosphere encountered microflora B. pumilus and B. subtilis, respectively (Damodaran et al., 2014). In the present study, the bacterial endophyte identified in higher Na+/K+ in the root portion of sodicity tolerant polyembryonic mango root stock belonged to the same group of firmicutes reported previously (Bharathkumar et al., 2008; Islam et al., 2010).
Therefore, this salt tolerant stability and adaptation of salt tolerant strains with capability to produce amylase, cellulase, protease and lipase enzymes (higher than the other strains of other researcher isolates) suggest that these strains could be a good choice for commer-cialisation for cultivation of crops in normal and salinesodic soil applications. In conclusion, elucidation of the origin of the potential endophytes clarifies influence of these endophytes in reclamation of salt affected soils.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
REFERENCES
|
Al-Mallah MK, Davey MR, Cooking EC (1987). Enzymatic treatment of clover root hairs removes a barrier to rhizobium-host specificity. Biotechnol. 5:1319-1322. |
|
|
Alonso A, Sanchez P, Martinez JL (2000). Stenotrophomonas maltophilia D457R contains a cluster of genes from Gram-positive bacteria involved in antibiotic and heavy metal resistance. Antimicrob. Agents Chemother. 44:1778-1782. |
|
|
Alqueres SMC, Almeida RV, Clementino MM, Vieira RP, Almeida WI, Cardoso AM, Martins OB (2007). Exploring the biotechnological applications in the archaeal domain. Braz. J. Microbiol. 38:398-405. |
|
|
Amoozegar MA, Malekzadeh F, Malik KA (2003). Production of amylase by newly isolated moderate halophile, Halobacillus sp. strain MA-2. Microbiol. Methods 52:353-359. |
|
|
Araujo WL, Marcon J, Maccheroni JW, Van Elsas JD, Van Vuurde JWL, Azevedo JL (2002). Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl. Environ. Micobiol. 68:4906-4914. |
|
|
Barzanti R, Ozino F, Bazzicalupo M, Gabbrielli R, Galardi F, Gonnelli C, Mengoni A (2007). Isolation and characterization of endophytic bacteria from the nickel hyper accumulator plant Alyssum bertolonii. Microb. Ecol. 53:306-316. |
|
|
Bassabrani D, Pallavi D, BalaGolpalan U, SawlongBorsing W, IromManoj S, Ramakrishna S (2012). Characterization of selected alpha-amylase producing bacterial strains isolated from soil samples of the alluvial plains of South Brahmaputra corridor of Assam. Afr. J. Microbiol. Res. 6(25):5335-5343. |
|
|
Bell CR, Dickie GA, Harvey WLG, Chan JWYF (1995). Endophytic bacteria in grapevine. Can. J. Microbiol. 41:46-53. |
|
|
Bharathkumar S, Kumar R, Paul D, Prabavathy VR, Nair S (2008). Characterization of the predominant bacterial population of different mangrove rhizosphere soils using 16S rRNA gene-based single-strand conformation polymorphism (SSCP). W. J. Microbiol. Biotechnol. 24:387-394. |
|
|
Borneman J, Skroch PW, O"Sullivan KM, Paulus JA, Rumjanek NG, Jansen JL, Nienhuis J, Triplett EW (1996). Molecular microbial diversity of an agricultural soil in Wisconsin. Appl. Environ. Microbiol. 62:1935-1943. |
|
|
Chowdhury SP, Schmid M, Hartmann A, Tripathi K (2007). Identification of diazotrophs in the culturable bacterial community associated with roots of Lasiurus sindicus, a perential grass of Thur desert, India. Microb. Ecol. 54:82-90. |
|
|
Damodaran T, Kannan R, Ahmed I, Srivastava RC, Rai RB, Umamaheswari S (2012). Assessing genetic relationships among mango (Mangifera indica L.) accessions of Andaman Islands using inter simple sequence repeat markers. New Zealand J. of Crop and Horti. Science.1:1-12. |
|
|
Damodaran T, Rai RB, Jha SK, Kannan R, Pandey BK, Sah V, Misra VK, Sharma DK (2014). Rhizosphere and endophytic bacteria for induction of salt tolerance in gladioulus grown in sodic soils. J. Plant Interaction. 9(1):577-584. |
|
|
Damodaran T, Sah V, Rai RB, Sharma DK, Mishra VK, Jha SK, Kannan R (2013). Isolation of rhizospheric bacteria by natural selection and screening for promising plant growth-promoting rhizobacteria (PGPR) and salt tolerance traits. Afr. J. Microbiol. 7(44):5082-5089. |
|
|
Dave BP, Anshuman K, Hajela P (2006). Siderophores of halophilic archeae and their chemical characterization. Indian. J. exp. Biol. 44:340-344. |
|
|
Deutch CE (2002). Characterization of a salt-tolerant extracellular α-amylase from Bacillus dipsosauri. Lett. Appl. Microbiol. 35:78-84. |
|
|
Devi LS, Khaund P, Joshi SR (2010). Thermostable amylase from natural variants of Bacillus spp. prevalent in eastern Himalayan Range. African. J. Microbiol. Res. 4:2534-2542. |
|
|
Dice LR (1945). Measures of the amount of ecologic association between species. Ecology 26. |
|
|
Espinosa-Victoria D, Lopez-Reyes L, De A, Cruz-benitez L (2009). Use of 16S rRNA gene for characterization of phosphate-solubilizing bacteria associated with corn. Rev. Fitotec. Mex. 32:31-37. |
|
|
Fahimeh M, Bahador N, Baserisalehi M (2013). Isolation, characterization and identification of amylase producing halothermophilic isolates from Howz Soltan Lake, Iran. Afr. J. Microb. Res. 7(36):4483-4490. |
|
|
Germaine K, Keogh E, Garcia-Cabellos G, Borremans B, Van der Lelie D, Barac T, Oeyen L, Vangronsveld J, Porteous Moore F, Moore ERB, Campbell CD, Ryan D, Dowling DD (2004). Colonisation of poplar trees by gfp expressing bacterial endophytes. FEMS Microbiol. Ecol. 48:109-118. |
|
|
Idris R, Trifonova R, Puschenreiter M, Wenzel WW, Sessitsch A (2004). Bacterial communities associated with flowering plants of the Ni hyper accumulator Thlaspi geoesingense. Appl. Environ. Microbiol. 70:2667-2677. |
|
|
Islam R, Trivedi P, Madhaiyan M, Seshadri S, Lee G, Yang J, Kim Y, Kim M, Han G, hauhan PS, Sa T (2010). Isolation, enumeration and characterization of diazotrophic bacteria from paddy soil sample under long-term fertilizer management experiment. Biol. Fertil. Soils. 46:261-269. |
|
|
Jacobs MJ, Bugbee WM, Gabrielson DA (1985). Enumeration, location and characterization of endophytic bacteria within sugar-beet roots. Can. J. Bot. 63:1262-1265. |
|
|
Kannan R, Damodaran T, Rai RB, Pandey BK, Sharma DK, Mishra VK, Jha SK, Sah V (2014). Isolation and characterization of endophytic plant growth-promoting bacteria (PGPB) associated to the sodicity tolerant polyembryonic mango (Mangifera indica L.) root stock and growth vigour in rice under sodic environment. Afr. J. Biotechnol. 8(7):628-636. |
|
|
Khan AA, Jilani G, Akhtar MS, Naqvi SMS, Rasheed M (2009) Phosphorus solubilizing bacteria: occurrence, mechanisms and their role in crop production. J. Agric. Biol. Sci. 1:48-58. |
|
|
Kobayashi DY, Palumbo JD (2000). Bacterial endophytes and their effects on plants and use in agriculture. In: Bacon CW and White JF (eds) Microbial endophytes. Marcel Dekker, Inc, NY. pp. 199-233. |
|
|
Kuklinsky-Sobral J, Araujo WL, Mendes R, Geraldi IO, Pizzirani-Kleiner AA, Azevedo JL (2004). Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ. Microbiol. 6:1244-1251. |
|
|
Moore FP, Tanja B, Brigitte B, Licy O, Jaco V, Daniel VL, Colin DC, Edward RBM (2006). Endophytic bacterial diversity in poplar trees growing on a BTEX-contaminated site: The characterisation of isolates with potential to enhance phytoremediation. System & Appld. Microbio. 29:539-556. |
|
|
Patriquin DG, Dobereiner J (1978). Light-microscopy observation of tetrazolium-reducing bacteria in endorhizosphere of maize and other grasses in Brazil. Can. J. Microbiol. 24:734-742. |
|
|
Prakash O, Jaiswal N (2009). Alpha-amylase: An ideal representative of thermostable enzyme. Appl. Biochem. Biotechnol. 5:14-18. |
|
|
Quadt-Hallmann A, Benhamou AN, Kleopper JW (1997). Bacterial endophytes in cotton: Mechanisms of entering the plant. Can. J. Microbiol. 43:577-582. |
|
|
Reinhold-Hurek B, Hurek T (1998). Life in grasses: Diazotrophic endophytes. Trend. Microbiol. 6:139-144. |
|
|
Sakiyama CCH, Paula EM, Pereira PC, Borges AC, Silva DO (2001). Characterization of pectin lyase produced by an endophytic strain isolated from coffee cherries. Lett. Appl. Microbiol. 33:117-121. |
|
|
Stierle A, Strobel G, Stierle D (1993). Taxol and taxane production by Taxomyces andreanae an endophytic fungus of Pacific yew. Sci. 260:214-216. |
|
|
Strobel GA (2003). Endophytes as sources of bioactive products. Microb. Infect. 5:535-544. |
|
|
Sturz AV, Christie BR, Nowak J (2000). Bacterial endophytes: potential role in developing sustainable systems of crop production. Crit. Rev. Plant. Sci. 19:1-30. |
|
|
Tank N, Saraf M (2010). Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J. Plant. Interact. 5:51-58. |
|
|
Thompson DC (1996). Evaluation of bacterial antagonist for reduction of summer patch symptoms in Kentucky blue grass. Plant. Dis. 80:856-862. |
|
|
Upadhyay SK, Singh DP, Saikia R (2009). Genetic diversity of plant growth promoting rhizobacteria isolated from rhizospheric soil of wheat under saline condition. Curr. Microbiol. 59:489-496. |
|
|
Van Houdt R, Monchy S, Leys N, Mergeay M (2009). New mobile genetic elements in Cupriavidus metallidurans CH34, their possible roles and occurrence in other bacteria. Antonie. Van. Leeuwenhoek. 96:205-226. |
|
|
Weon Taek Seo, Woo Jin Lim, Eun Jin Kim, Han Dae Yun, Young Han Lee Kye Man Cho (2010). Endophytic bacterial diversity in the young radish and their antimicrobial activity against pathogens. J. Korean. Soc. Appl. Biol. Chem. 53(4):493-503. |
|
|
Yap IV, Nelson RJ (1996). Winboot: A program for performingbootstrap analysis for binary data to determine the confidence limits of UPGMA based dendrograms. International Rice Research Institute, Discussion Paper Series No 14. |
|
|
Zinniel KD, Lambrecht P, Harris NB, Feng Z, Kuczmarshki D, Higley P, Ishimaru CA, Arunakumari A, Barletta RG, Vidaver AK (2002). Isolation and characterization of endophytic bacteria from agronomic crops and Prairie plants. Appl. Environ. Microbiol. 68:2198-2208. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


