Full Length Research Paper

ABSTRACT
One of the primary objectives in agriculture is providing high-quality crops to consumers. Multiple techniques and methods are utilized to achieve this objective, including nanotechnology that depends on the use of very small materials, which will help in decreasing the amounts usually used with similar effects. Nanomaterials are used as fertilizers and also as component of nano-pesticides for plants. Despite their benefits, however, studies have noted their potential for cytotoxicity and genotoxicity. In this study, five nanoparticles (NPs) were tested to assess their effects on plants. The chromosomal aberration assay was used. The results showed that some NPs decreased the mitotic index (MI) significantly, which indicates the NPs’ potential cytotoxicity. In addition, different NPs’ treatments caused different types of chromosomal abnormalities e.g., chromosomes stickiness and disturbance of the metaphase and anaphase, lagging chromosomes, bridges, disturbed poles, micronuclei, s-metaphase, s-telophase, c- metaphase and bi-nucleus cells. All treatments had significant effects at p≤005. Treatments with NPs concentrations for 24 h affected the DNA content, AlO2 and Fe3O4 NPs’ increased the DNA content, while CeO2, TiO2 and Ag NPs’ decreased it. High concentrations of the tested NPs decreased the DNA content. The study results showed that CeO2 was the most harmful NP compared to the control and other NPs. Some types of chromosome abnormalities such as lagging chromosomes, bridge, and micronuclei indicate potential genotoxicity for these NPs. Despite of the positive effects, they also had negative side effects such as decreasing the MI and increasing the occurrence of different types of chromosomal abnormalities.
Key words: Cytotoxicity, genotoxicity, nanoparticles, mitotic chromosomes abnormalities.
INTRODUCTION
MATERIALS AND METHODS
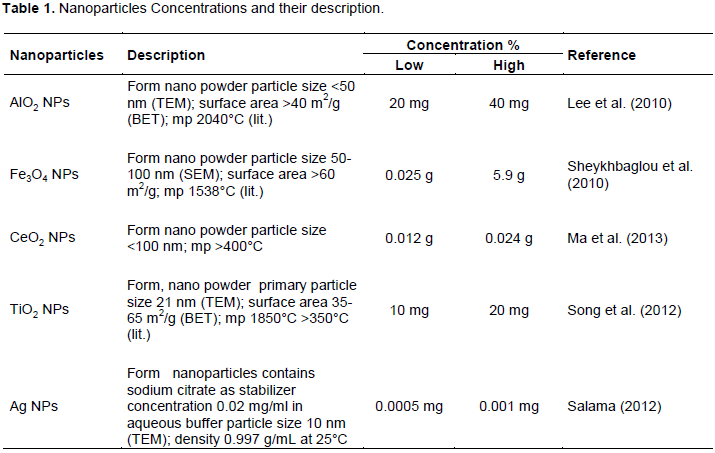
RESULTS
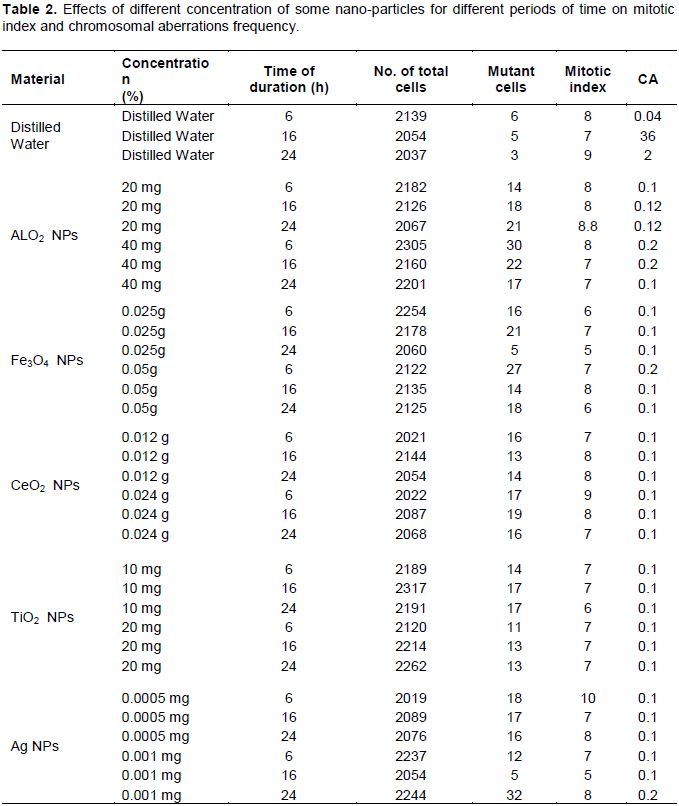
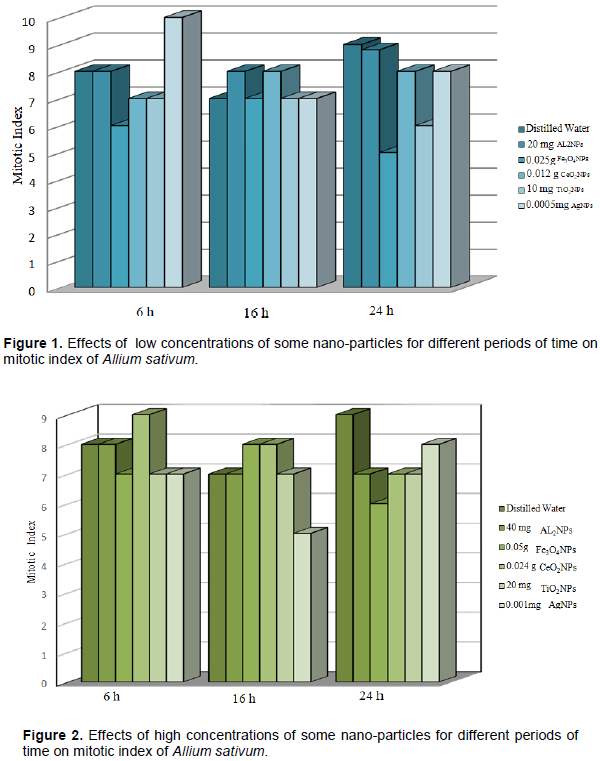
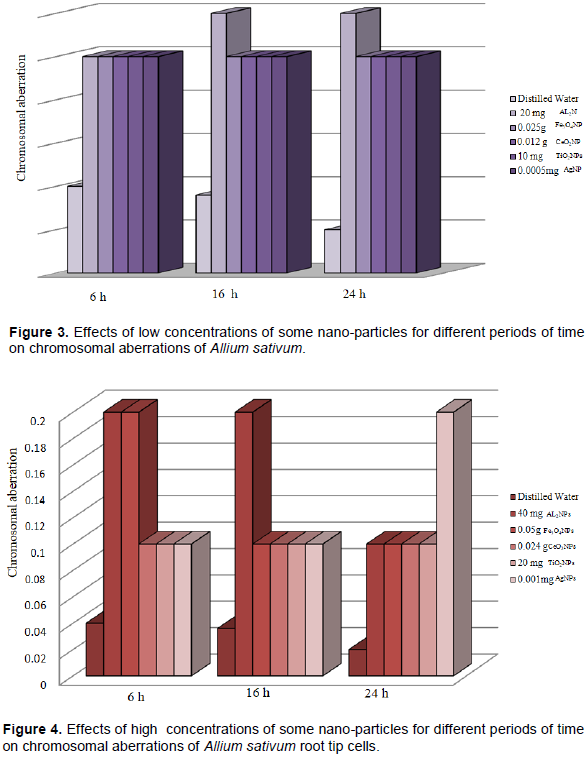
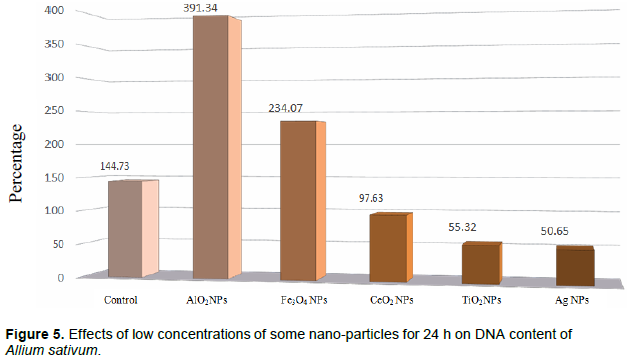
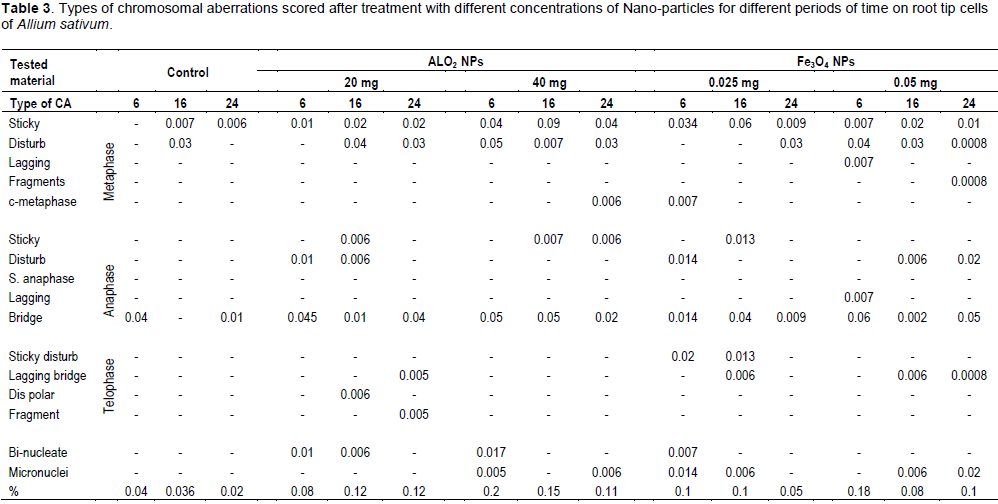
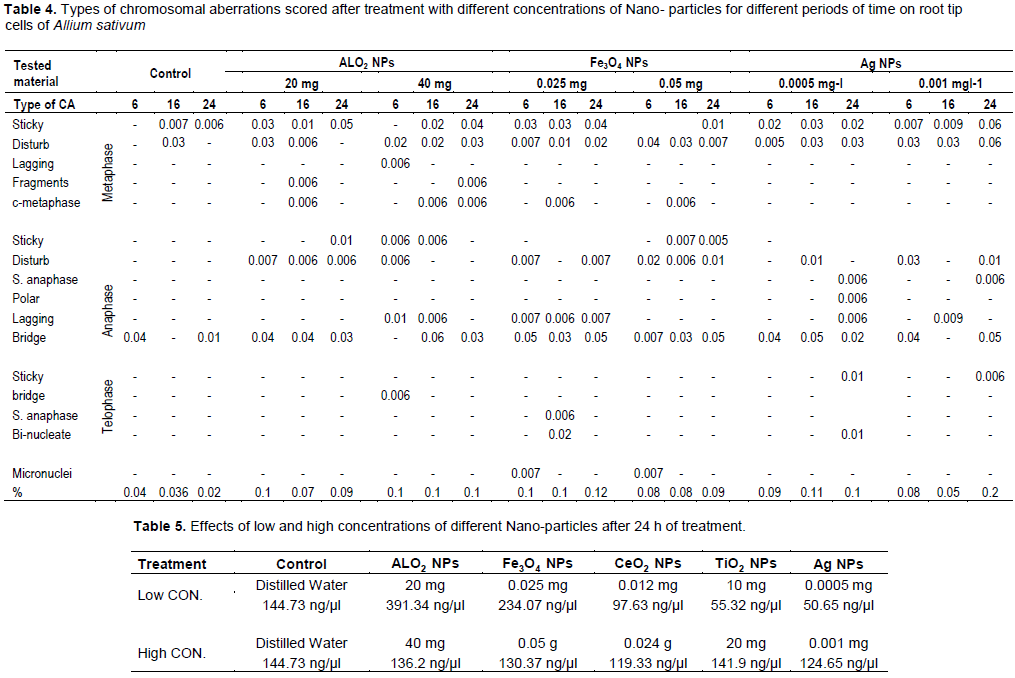
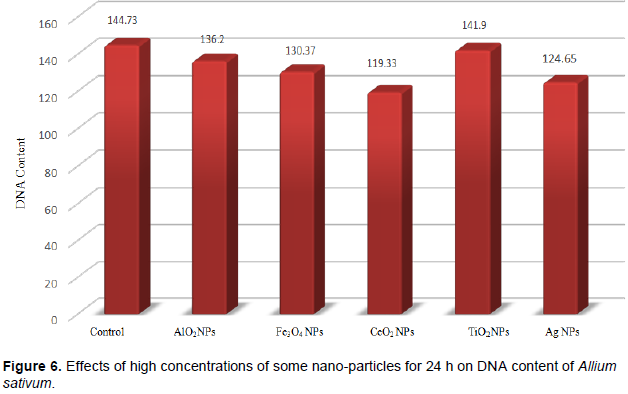
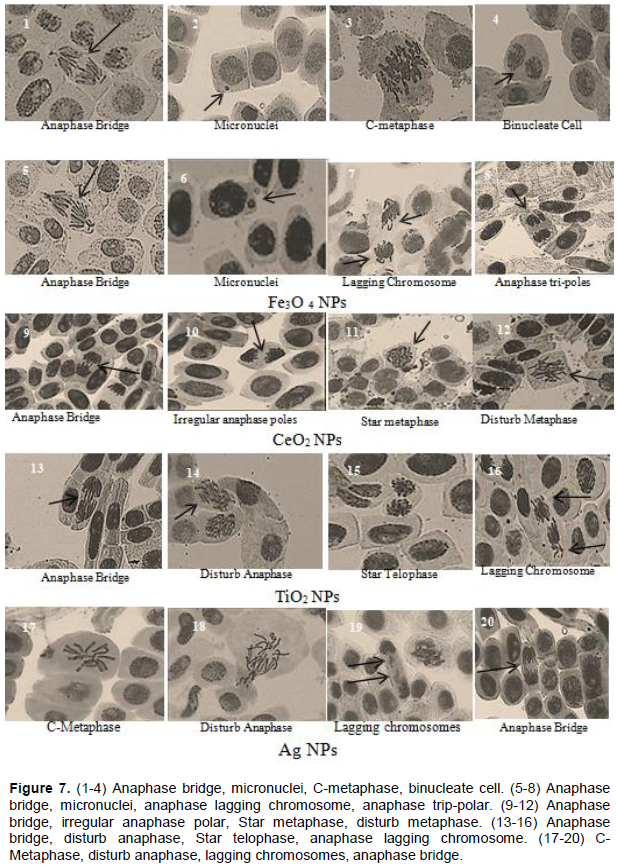
DISCUSSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Afaq F, Abidi P, Matin R, Rahman Q (1998). Cytotoxicity, pro-oxidant effects and antioxidant depletion in rat lung alveolar macrophages exposed ultrafine titanium dioxide. Journal of Applied Toxicology 18(5):307-312. |
|
|
Alarifi S, Ali D, ALkahtani S, Alhader MS (2014). Iron oxide nanoparticles induce oxidative stress, DNA damage, and caspase activation in the human breast cancer cell line. Biological Trace Element Research 159(1-3):416-424. |
|
|
Auffan M, Rose J, Orsiere T, De Meo M, Thill A, Zeyons O, Proux O, Masion A, Chaurand P, Spalla O, Botta A, Wiesner MR, Bottero JY (2009). CeO2 nanoparticles induced DNA damage towards human dermal fibroblasts in vitro. Nanotoxicology 3(2):161-171. |
|
|
Babu K, Deepa MA, Shankar SG, Rai S (2008). Effect of nano-Silver on Division and mitotic chromosomes: A prefatory Siren. Internet Journal of Nanotechnology 2(2):1-7. |
|
|
Benameur L, Affuan M, Cassien M, Liu W, Culcasi M, Rahmouni H, Stocker P, Tassistro V,Bottero J, Rose J, Botta A, Pietri S (2015). DNA damage and oxidative stress induced by CeO2 nanoparticles in human dermal fibroblasts: Evidence of a clastogenic effect as a mechanism of genotoxicity. Nanotoxicology 9(6):696-705. |
|
|
Brunner TJ, Wick P, Manser P, Spohn P, Grass RN, Limbach LK, Bruinink A, Stark WJ (2006). In vitro cytotoxicity of oxide nanoparticles: Comparison to asbestos, silica, and the effect of particle solubility. Environmental Science & Technology 40(14):4374-4381. |
|
|
Buzea C, Pacheco BI, Robbie K (2007). Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases 2(4):17-72. |
|
|
Castiglione MR, Giorgetti L, Geri C, Cermonini R (2011). The effects of nano-TiO2 on seed germination, development and mitosis of root tip cells of Vicia narbonensis L. and Zea mays L. Journal of Nanoparticle Research 13(6):2443-2449. |
|
|
ÒªekiÒ« FÖ, Ekinci S, Inal MS, Ünal D (2017). Silver nanoparticles induced genotoxicity and oxidative stress in tomato plants. Turkish Journal of Biology 41(5):700-707. |
|
|
Chen M, von Mikecz A (2005). Formation on nucleoplasmic protein aggregates impairs nuclear function in response to SiO2 Nanoparticles. Experimental Cell Research 305(1):51-62. |
|
|
Debnath P, Mondal A, Hajra A, Das C, Mondal K (2018). Cytogenetic effects of silver and gold nanoparticles on Allium cepa roots. Journal of Genetic Engineering and Biotechnology 16(2):519-526. |
|
|
Donaldson K, Beswick PH, Gilmour PS (1996). Free radical activity associated with the surface of particles: a unifying factor in determining biological activity. Toxicology Letters 88(1-3):293-298. |
|
|
EL-Khodar S, Habib A, Haliem A (1990). Effect of the herbicides tribunnil on root mitosis of Allium cepa. Cytologia 55:209-215. |
|
|
Faisa M, Saquib Q, Alatar A, Han MAM (2016). Cobalt oxide nanoparticles aggravate DNA damage and cell death in eggplant via mitochondrial swelling and NO signaling pathway. Biolgical Research 49:20. |
|
|
Ghormade V, Deshpande MV, Pakmikar KM (2011). Perspectives for nano-biotechnology enabled protection and nutrition of plants. Biotechnology Advances 29(6):792-803. |
|
|
Ghosh M, Bandyopadhyay M, Mukherjee A (2010). Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophies levels: Plant and human lymphocytes. Chemosphere 81(10):1253-62 |
|
|
Ghosh M, Manivannan J, Silnha S, Chakraborty A, Mallic SK, Bandyopadhyay M, Mukherjee A (2012). In vitro and in vivo genotoxicity of silver nanoparticles. Mutation Research 749(1-2):60-69. |
|
|
Gilmour P, Brown DM, Beswick PH, Benton E, MacNee W, Donaldson K (1997). Surface free radical activity of PM10 and ultrafine titanium dioxide: a unifying factor in their toxicity? The Annals of Occupational Hygiene 41(1):32-38. |
|
|
Golbamaki N, Rasulev B, Cassano A, Robinson RIM, Benfenati E, Leszczynski J, Cronin MTD (2015). Genotoxicity of metal oxide nanomaterials: Review of recent data and discussion of possible mechanisms. Nanoscale 7(6):2154-98. |
|
|
Grant WF (1978). Chromosome Aberrations in Plants as a Monitoring System. Environmental Health Perspectives 27:37-43 |
|
|
Grant WF (1982). Chromosome aberration assay in Allium. A report of the US Environmental Protection Agency Gene-Tox Program. Mutation Research 99(3):273-291. |
|
|
Grant WF (1999). Higher plant assays for the detection of chromosomal aberrations and gene mutations-a brief historical background on their use for screening and monitoring environmental chemicals. Mutation Research /Fundamental and Molecular Mechanisms of Mutagenesis 426(2):107-112. |
|
|
Huang S, Chueh PJ, Lin YW, Shih TS, Chuang SM (2009). Disturbed mitotic progression and genome segregation are involved in cell transformation mediated by nano-Tio2 long-term exposure. Toxicology and Applied Pharmacology 241(2):182-94. |
|
|
Huijing B, Xiaoxu Y, Xhen X, Zhaoyang L, Dianjun W, Yunde L (2015). New Toxicity Mechanism of Nanoparticles. Promoting Apoptosis and Inhibiting Proliferation. PLoS One 10(3):e0122535. |
|
|
Hunt PR, Marquis BJ, Tyner KM, Conklin S, Olejnik N, Nelson BC, Sprand RL (2013). Nano silver suppresses growth and induces oxidative damage to DNA in Caenorhabditis elegans. Journal of Applied Toxicology 33(10):1131-1142. |
|
|
Jaskowiak J, Tkaczyk O, Slota M, Kwasniewska J, Szarejko L (2018). Analysis of Aluminum toxicity in Hordeum vulgare roots with an emphasis on DNA integrity and cell cycle. PLoS One 13(2):e0193156 |
|
|
Jezek P, Hlavata I (2005). Mitochondria in homeostasis of reactive oxygen species in cell, tissues and organism. The International Journal of Biochemistry and Cell Biology 37(12):2478-503. |
|
|
Khodakovskaya MV, Silva K, Biris AS, Dervishi E, Villagarcia H (2012). Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 6(3):2128-2135. |
|
|
Klasterska I, Natarajan AT, Ramel C (1976). An interpretation of the origin of sub chromatid aberration and chromosome stickiness as a category of chromatid aberrations. Hereditas 83(2):153-162. |
|
|
Klien K, Godnic J (2012). Genotoxicity of Metal Nanoparticles: Focus on in vivo Studies. Arh Hig Rada Toksikol 63(2):133-45. |
|
|
Kovacic P, Somanathan R (2010). Biomechanisms of nanoparticles (toxicants, antioxidants and therapeutics): Electron transfer and reactive oxygen species. Journal of Nanoscience and Nanotechnology 10(12):7919-7930. |
|
|
Kruszewski M, Brzoska K, brunborg G, Asare N, Dobrzynska M, Dusinska M, Marie Fjellsbo L, Georgantzopoulou A, Gromadzka-Ostrowska J, Gutleb AC, Lankoff A, Magdolenova Z, Pran ER, Rinna A, Instanes Ch Sandberg WJ, Schwarze PE, Maciej Stepkowski T, Wojewodzka M, Refsnes M (2011). Toxicity of silver nanomaterials in higher eukaryotes. Advances in Molecular Toxicology 5:179-218. |
|
|
Kumari M, Kumari SI, Grover P (2014). Genotoxicity analysis of cerium oxide micro and nanoparticles in Wistar rats after 28 days of reported oral administration. Mutagenesis 29(6):467-479. |
|
|
Kumari M, Mukherjee A, Chandrasekaran N (2009). Genotoxicity of Silver Nanoparticles on Allium cepa. Science of The Total Environment 407(19):5243-5246. |
|
|
Kuriyama R, Sakai H (1974). Role of tubulin-SH group in polymerization to microtubules. Journal of Biochemistry 76(3):651-654. |
|
|
Kwon JY, Koedrith P, Seo YR (2014). Current investigations into the genotoxicity of Zinc oxide and silica nanoparticles in mammalian models in vitro and in vivo. Carcinogenic/ genotoxic potential relevant mechanisms and biomarkers, artifacts, and limitations. International Journal of Nanomedicine 9(Suppl 2):271-86. |
|
|
Landa P, Vankova R, Andrlova I, Hodek J, Marsik P, Storchova H, White JC, Vanek T (2012). Nanoparticle-specific changes in Arabidopsis thaliana gene expression after expouser to ZnO, TiO2, and gulleren soot. Journal of Hazardous Materials 241:55-62 |
|
|
Landsiedel R, Kapp MD, Schulz M, Wienc, K, Oesch F (2009). Genotoxicity investigations on nanomaterials: Methods, preparation and characterization of test material, potential artifacts and limitations- Many questions, some answers. Mutation Research 681(2-3):241-58. |
|
|
Lee CW, Mahendra S, Zodrow K, Li DM, Tsai YC, Braam , lvarez PJJ (2010). Developmental phytotoxicity of metal oxide nanoparticles to Arabidopsis thaliana L. Environmental Toxicology and Chemistry 29(3):669-675. |
|
|
Liman R, Acikbas Y, Hakki I (2019). Cytotoxicity and genotoxicity of cerium oxide micro and nanoparticles by Allium and comet tests. Ecotoxicology and Environmental Safety 168:408-414. |
|
|
López-Moreno M, Guadalupe R, José ÁH, Hiram C, Cristina E, José RP, Jorge LG (2010). Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environmental Science and Technology 44(19):7315-7320. |
|
|
Ma C, Chhikara S, Xing B, Musante C, White JC, Dhankher OP (2013). Physiological and molecular response of Arabidopsis thaliana L. to nanoparticle cerium and indium oxide exposure. ACS Sustainable Chemistry and Engineering 1(7):768-778. |
|
|
Ma TH (1982). Vicia cytogenetic tests for environmental mutagen. A report of the US Environmental Protection Agency Gene-Tox Program. Mutation Research 99(3):257-271. |
|
|
Magdolenova Z, Collins A, Kumar A, Dhawan A, Stone V, Dusinska M (2014). Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 8(3):233-78. |
|
|
Mattiello A, Filippi A, Poscic F, Musetti R, Salvtici MC, Giordano C, Vischi M, Bertolini A, Marchiol L (2015). Evidence of Phytotoxicity and Genotoxicity in Horeum vulgare L. Exposed to Ceo2 and TiO2 Nanoparticles. Forntiers in Plant Science 6:1043. |
|
|
McShan D, Ray PC, Yu H (2014). Molecular toxicity mechanism of nano silver. Journal of Food and Drug Analysis 22(1):116-127. |
|
|
Mehrian SK, De Lima R (2016). Nanoparticles cyto and genotoxicity in plants: Mechanisms and abnormalities. Environmental Nanotechnology, Monitoring and Management 6:184-193. |
|
|
Mohandas T, Grant WF (1972). Cytogenetic effect of 2,4-D and amitol in relation to nuclear volume DNA content in some higher plants. Canadian Journal of Genetics and Cytology 14(4):773-783. |
|
|
Mukherjee A, Peralta-Videa J, Gardea-Torresdey J (2016). Effects and Uptake of Nanoparticles in Plants. Engineered Nanoparticles and the Environment: Biophysicochemical Processes and Toxicity. |
|
|
Pakrashi S, Jerobin J, dalai S, Prathna TC (2014). In vivo Genotoxicity Assessment of Titanium Dioxide Nanoparticles by Allium cepa Root Tip Assay at High Exposure Concentration. PLoS One 9(2):e87789. |
|
|
Park EJ, Choi J, Park YK, Park K (2008). Oxidative stress induced by cerium oxide nanoparticles in cultured BEAS-2B cells. Toxicology 245(1-2):90-100. |
|
|
Patil BC, Bhat GI (1992). A comparative study of MH and EMS in the induction of chromosomal aberration on lateral root meristem in Clitoria ternate L. International Journal of Cytology 57(2):259-264. |
|
|
Patlolla AK, Berry A, May L, Tchounwou PB (2012). Genotoxicity of silver nanoparticles in Vicia faba: a pilot study on the environmental monitoring of nanoparticles. International Journal of Environmental Research and Public Health 9(5):1649-1662. |
|
|
Pesnya DS (2013). Cytogenetic effects of chitosan-capped silver nanoparticles in the Allium cepa test. International Journal of Cytology, Cytosystematics and Cytogenetics 66(3):275-281. |
|
|
Pulate PV, Ghurde MU, Deshmukh VR (2011). Cytological effects of the biological and chemical silver-nanoparticles in Allium cepa L. International Journal of Innovations in Biological and Chemical Sciences 1:32-35. |
|
|
Qian H, Peng X, Han X, Ren J, Zhengwei F (2013). Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana. Journal of Environmental Sciences 25(9):1947-1955. |
|
|
Rajeshwari A, Kavitha S, Sruthi Ann A, Deepak K, Anita M, Natarajan C, Amitava M (2015). Cytotoxicity of Aluminum oxide nanoparticles on Allium cepa root tip- effects of oxidative stress generation and bio uptake. Environmental Science and Pollution Research 22(14):11057-66. |
|
|
Rajiv S, Erobin J, Saranya V, Nainawat M, Sharma M, Makwana P, Gayathri C, Bharath L, Singh M, Kumar M, Mukherjee M, Chandrasekaran N (2015). Comparative cytotoxicity and genotoxicity of cobalt (II, III) oxide, iron (III) oxide, silicon dioxide, and aluminum oxide nanoparticles on human lymphocytes in vitro. Human and Experimental Toxicology 35(2):170-183 |
|
|
Rico CM, Duarte S, Garden M, Peralta-Videa JR, Gardea-Torrcsdcy JL (2011). Interaction of nanoparticles with edible plants and their possible implications in the food chain. Journal of Agricultural and Food Chemistry 59(8):3485-3498. |
|
|
Saha N, Gupta SD (2017). A Glimpase on Silver Nanoparticles Genotoxicity in Higher Plants. Global Journal of Nanomedicine 2(2):1-2. |
|
|
Salam HMH (2012). Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea mays L.). International Research Journal of Biotechnology 3(10):190-197. |
|
|
Schins RP, Knaapen AM (2007). Genotoxicity of poorly soluble particles. Inhalation Toxicology 19(Suppl 1):189-198. |
|
|
Sendra M, Moreno-Garrido G, Yeste P, Gatica JM, Blasco J (2016). Behaviour of CeO2 nanoparticles and bulk and their toxicity in freshwater and seawater microalgae. Universidad de Alicante pp. 99-101. |
|
|
Sharma P, Jha AB, Dubey RS, Pessarakli M (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. Journal of Botany 217037:1-26. |
|
|
Sheykhbaglou R, Sedghi M, Shishevan MT, Sharifi RS (2010). Effects of nano-iron oxide particles on agronomic traits of soybean. Notulae Scientia Biologicae 2(2):112-113. |
|
|
Siddiqui MH, Mohamed HW, Mohammad F, Mutahhar YK (2015). Role of Nanoparticles in Plants. Nanotechnology and Plant Sciences 19-35 |
|
|
Silva JR, Smyth TJ, Moxley DF, Carter TE, Allen NS, Rufty TW (2000). Aluminum accumulating at nuclei of cell in the root tip. Fluorescence detection using lumogallion and confocal laser scanning microscopy. Plant Physiology 123(2):543-552. |
|
|
Singh N, Manshian B, Jenkins GJ, Griffiths SM, Williams PM, Maffeis TG, Wright CJ, Doka SH (2009). The DNA damaging potential of engineered nanomaterials. Biomaterials 30(23-24):3891-3914. |
|
|
Sjorgen CA, Larsen PB (2017). SUV2, which encode an ATR-related cell cycle checkpoint and putative plant ATRIP, is required for aluminum-dependant root growth inhibition in Arabidosis. Plant, Cell and Environment 40(9):1849-1860. |
|
|
Song G, Gao Y, Wu H, Hou W, Zhang C, Ma H (2012). Physiological effect of anatase TiO2 nano-particles on Lemna minor. Environmental Toxicology and Chemistry 31(9):2147-2152. |
|
|
Srinivas A, Rao PJ, Selam G, Murthy PB, Reddy PN (2011). Acute inhalation toxicity of cerium oxide nanoparticles in rats. Toxicology Letters 205(2):105-115. |
|
|
Sudhakar R, Ninge Gowda KN, Govindappa V (2001). Mitotic Abnormalities Induced by Silk Dyeing Industry Effluents in the Cells of Allium cepa. Cytologia 66(3):235-239. |
|
|
Tavares AM, Louro H, Antunes S, Quarre S, Simar S, De Temmerman PJ, Verleysen E, Mas J, Jense KA, Norppa H, Nesslany F, Silva MJ (2014). Genotoxicity evaluation on nanosized titanium dioxide, synthetic amorphous silica and multi-walled carbon nanotubes in human lymphocytes. Toxicology in Vitro 28(1):60-90. |
|
|
Trouiller B, Reliene R, Westbrook A, Solaimani P, Schiestl RH (2009). Titanium Dioxide Nanoparticles induce DNA Damage and Genetic Instability in vivo in Mice. Cancer Research 69(22):8784-8789 |
|
|
Turkez H, Geyikoglu F (2007). An in vivo blood culture for evaluating the genotoxicity of titanium dioxide: the response of antioxidant enzymes. Toxicology and Industrial Health 23(1):19-23. |
|
|
Van Breusegem F, Dat JF (2006). Reactive oxygen species in plant cell death. Plant Physiology 141:384-390. |
|
|
Vranová E, Inzé D, Van Breusegem F( 2002). Single transduction during oxidative stress. Journal of Experimental Botany 53(372):1227-1236. |
|
|
Wu H, Yin J, Wamer WG, Zeng M (2014). Reactive oxygen species-related activities of nano-iron metal and nano-iron oxides. Journal of Food and Drug Analysis 22(1):86-94. |
|
|
Yuliang Z, Bing W, Weiyue F, Chunli B (2010). Nanotoxicology: toxicological and biological activities of nanomaterials. In: Encyclopedia of Life support Systems (EOLSS): nanoscience and nanotechnologies. Paris: UNESCO-EOLSS Publisher. |
|
|
Zheng L, Hong S, Lu C (2005). Effect of TiO2 on strength of naturally aged seeds and growth of spinach. Biological Trace Element Research 104(1):83-92. |
|
|
Zhongwen Chen, Jun-Jie Yin, Yu-Ting Zhou, Zhang Y, Song L, Song M, Hu S, Gu N (2012). Dual enzyme-like activities of iron oxide nanoparticles and their implication for diminishing cytotoxicity. ACS Nano 6(5):4001-4012. |
|
|
Zia-ur-Rehman M, Qayyum MF, Akmal F, Maqsood MA, Rizwan M, Waqar M, Azhar M (2018). Nanomaterials in Plants, Algae, and Microorganisms. Recent Progress of Nanotoxicology in Plants- Science Direct pp. 143-174. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


