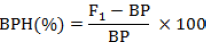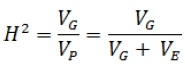ABSTRACT
A 2x2 diallel mating design was set up to estimate quantitative genetic parameters after leaf artificial inoculation tests in nursery and field for three consecutive years. F12 and F40 families of two reciprocal crosses viz., F12: ♀SCA12 × â™‚ICS40 and F40: ♀ICS40 × â™‚SCA12 were used to study heterosis (mid-parent), heterobeltiosis (better-parent), heritability (narrow-sense and broad-sense) and to quantify total polyphenols (TPP) and total flavonoids (TF) contents after inoculation of the leaves of parental and hybrid cocoa genotypes with Phytophthora megakarya. The results of heterosis and heterobeltiosis exhibited hybrid vigor of 86.66 and 85%, respectively. The highest heritability was identified [narrow-sense heritability (h²: 0.668 for F12 and 0.682 for F40) and broad-sense heritability (H²: 0.765 for F12 and 0.672 for F40)] for necrosis length in 2014. With regards to low narrow-sense heritability, it ranged from 0.019 to 0.162 while corresponding broad-sense heritability ranged from 0.476 to 0.562 for biochemical compounds. Among parent clones, ICS40 and SCA12 proved to have the general combining abilities (GCA). The use of these parental lines and understanding the gene flow of tolerance to black pod disease (Bpd) in cocoa will help researchers to develop a targeted breeding approach in generating new generation of tolerant genotypes cocoa.
Key words: Cocoa, heterosis, heterobeltiosis, heritability, black pod disease, tolerance.
Cocoa tree (Theobroma cacao L.) is a woody plant belonging to the Magnoliopsideae class. Because of its ecological requirements, cocoa farming has been largely practiced in Africa (73% of world production, with 4.552 million tons), where four countries are among the five best producers in the world: Côte d'Ivoire (42%), Ghana (19%), Nigeria ranked behind Indonesia (7%), Ecuador (6.5%) and Cameroon (6%) (Anonymous, 2017). Although Cameroon is the fourth world cocoa producer, her cocoa crop is experiencing production setbacks due to black pod disease (Bpd) caused by an oomycete denominated Phytophthora megakarya (Nyasse, 1997). Currently, disease control relies heavily on the use of chemical fungicides which has in turn increased the production costs and reduced growers’ profits. In addition, there is a growing concern on the impact of chemical fungicides on the environment, which are costly and polluting for cocoa farmers. Genetic control based on hybridization allows the development and adoption of genotype tolerants to Bpd. This control strategy is the most preferred to improve production (Djocgoue et al., 2010; Ondobo et al., 2017). Diallel cross analysis for a fixed set of populations provides a basis for preliminary determination of heterotic groups. Thus, knowledge on the nature and magnitude of genotypic and phenotypic variability present in the crop species plays a vital role in formulating a successful breeding program to evolve superior cultivars. Hence, heterosis per se is commercially useful.
However, heterosis (mid-parent) and heterobeltiosis (better-parent) are useful in shaping the direction that future hybrid breeding program should take. It also identifies the cross combinations which are promising in conventional breeding programs. Studies on heterosis would help to generate breeding strategies of hybrid cocoa production. Obtaining genetic information from diallel cross progenies is a common practice of plant breeders working with allogamous crops.
However, the estimation of heritability serves as a useful guide to the breeder. By relying on heritability, the breeder is able to appreciate the production of the variation that is due to genotypic (broad-sense heritability) or additive (narrow-sense heritability) effects. If heritability of a character is very high, selection for the character should be fairly easy. Thus, estimates of heritability are useful in predicting the transmission of characters from the parents to their offspring (Parikh et al., 2016). The present study was conducted firstly to estimate heterosis (mid-parent and better-parent) and heritability (broad-sense and narrow-sense) for necrosis length traits in a 2x2 diallel set of cocoa leaf and secondly to identify hybrids of T. cacao that are tolerant to Bpd and to observe good aptitudes for the combination of parent.
Plant material
The experiment was conducted at the Cocoa Development Corporation (SODECAO) experimental field at Mengang Station. Two cocoa parental clones viz., SCA12 (moderately tolerant to Bpd and Forastero group) and ICS 40 (sensitive to Bpd and local Trinitario group) were used to produce two reciprocal offspring populations (F12: ♀SCA12 × â™‚ICS40 and F40: ♀ICS40 × â™‚SCA12) by hand-pollination techniques for three seasons in three years (2012/2013, 2013/2014 and 2014/2015). These parents were crossed using 2x2 full diallel mating design. The parents were grafted in the nursery using bud wood.
Plant inoculation
P. megakarya-lebdi isolate [obtained from the Central Laboratory of Phytopathology at IRAD (Research Institute for Agricultural Development)] was grown on V8 agar medium (20% V8 vegetable juice, 0.3% Ca2CO3, 1.5% agar and 1000 ml distilled water) and incubated in the dark at 25 ± 2°C. Artificial inoculation of leaves of 22 genotypes was carried out in 2013, 2014 and 2015 (three-season period) adapted from Djocgoue et al. (2006). They were then inner surface sterilized with ethanol 70%. Three treatments were performed on each parental genotype and hybrid: (i) healthy, (ii) scarified (iii) and infected leaves. Agar disks (6 mm diameter) cut from 5-day-old fungal and straminopilous isolates were laid on the midrib after creating wounding with a sterilized razor blade. The scars were then covered with cotton that had been immersed in sterilized water. The necrosis length was measured at two days interval with a graduated ruler after inoculation. The samples were wrapped with aluminum foil every six days after inoculation.
Estimation of heterosis and heterobeltiosis
The values of heterosis over mid and better parents from F1 were calculated for length necrosis.
(i) Mid-parent heterosis (Zahour, 1992) was computed using the formula:
(ii) Better-parent heterosis (or heterobeltiosis) was calculated using the procedure of Tang and Xiao (2013), following the formula below:
In these formulas, MPH (%) represents the percentage of mid-parent heterosis, BPH (%) is the better-parent heterosis percentage, F1 refers to the means of hybrid genotype, MP is the means of mid-parent and BP is the means of better-parent.
Estimation of heritabilities (h² and H²)
For necrosis length, two types of expression of heritability were calculated from formulas suggested by Falconer and Mackay (1996):
(i) Narrow-sense heritability (h²) measures the variance additive genetic.

(ii) Broad-sense heritability (H²) measures any genetic variance.

In these formulas, is the genetic variance, is the phenotypic variance, is the additive genetic variance and is the environmental variance.
Extraction and total polyphenolic contents
The total polyphenolic (TPP) content was determined using the Folin−Ciocalteu method (Georgé et al., 2005) with some modifications. Leaves (0.5 g) were mixed with 2 ml of acetone/water (70:30, v/v) and incubated at room temperature. The mixture was centrifuged (Labofuge 400R) at 4500 rpm for 5 min. The supernatants were applied three successive washes with cyclohexane eliminate lipids, carotenoids and chlorophyll (Wieslaw et al., 1988). 2.5 ml of Folin-Ciocalteu (1:10) reagent was added to 0.5 ml of diluted crude extract (1:10) in a test tube. The mixture was incubated for 2 min in the dark and at room temperature. Then, 2 ml of Na2CO3 (75 g/L) were added and the whole was stirred on a vortex and then incubated for 15 min at 50°C in a water bath in the dark. Afterwards, the mixture was cooled in an ice cube tray, the absorbance at 760 nm was immediately measured (Jenway 6305 spectrophotometer) against a blank in which the extract was replaced by distilled water. Results were expressed as gallic acid equivalents (GAE).
Extraction of flavonoids
This was done following the extraction protocol described by Counet et al. (2004), with minor modifications. The plant material (cocoa leaves) lyophilized (50 mg) and crushed was extracted with 2 ml of the acetone/water/acetic acid (70:28:2 v/v/v). After upheaval at vortex, the mixture was incubated at room temperature for 1 h in the dark. The bottom was centrifuged (Labofuge 4000R) at 4500 rpm for 5 min and the supernatant was recovered and constituted the raw extracts. Raw extract undergone three successive washes with 1 x 3 ml of hexane in order to remove lipids, carotenoids and chlorophyll (Wieslaw et al., 1988).
Total flavonoids contents
The total flavonoids (TF) content was determined using the aluminum trichloride (AlCl3) method (Ayoola et al., 2008). 2 ml of distilled water and 150 μl of a 5% (w/v) sodium nitrite solution (NaNO2) were added to 500 μL of diluted extract (1/10). The mixture was then incubated for six min in the dark and at ambient temperature (25 ± 2°C). 150 μL of the AlCl3 solution (10% in methanol; m/v) was added. After incubation for 6 min, at room temperature and in the dark, 2 ml of 4% (w/v) sodium hydroxide (NaOH) and 200 μL were rapidly added to each tube. The reaction medium (5 ml) was homogenized before being incubated in the dark for 15 min, and the absorbance as recorded at 510 nm using a UV spectrophotometer (Jenway 6305). Quercetin was used for the construction of a standard curve and the results were expressed as mg quercetin equivalent (QE) per grams dry weight (DW) of plant material. The analyses were performed in triplicate.
Statistical analysis
Data obtained from the observation were analyzed. SPSS (version 24.0 for windows) was used to perform analysis of variance (ANOVA). The significance of differences was determined by Tukey’s multiple comparison technique. The Rho Spearman correlation was used to determine the correlation between all the parameters evaluated in the different treatments. Hierarchical classification and principal component analysis (PCA) of necrotic surface area data was performed with SPAD 5.5 software package.
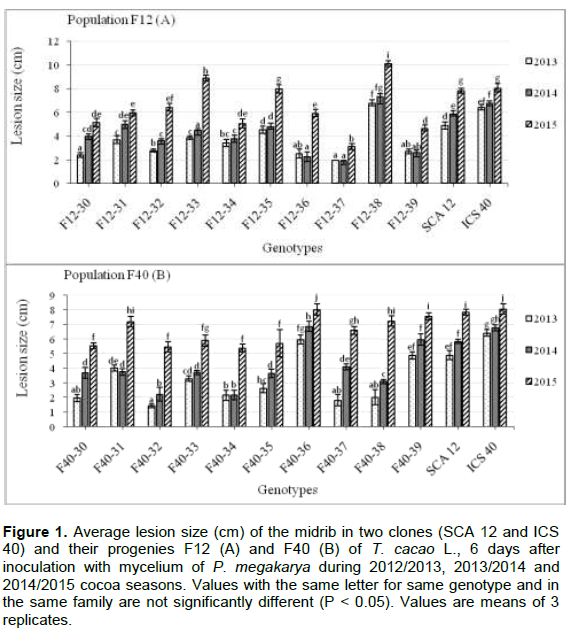
Length of necrosis
The development of necrosis for each of the three years: 2013, 2014 and 2015 was observed in parents (SCA12 and ICS40) and in hybrid populations F12 and F40 (Figure 1). However, individuals F12-36 [2.53±0.4 cm (2013), 2.26±0.42 cm (2014) and 5.93±0.31 cm (2015)], F12-37 [1.98±0.01 cm (2013), 1.93±0.35 cm (2014) and 3.13±0.25 cm (2015)] and F12-39 [2.70±0.21 cm (2013), 2.60±0.35 cm (2014) and 4.67±0.25 cm (2015)] showed a decrease in the development of necrosis at the level of 2014 (Figure 1A). For this purpose, the same slides were observed in individuals F40-31 [4.03±0.21 cm (2013), 3.77±0.23 cm (2014) and 7.20±0.36 cm (2015) and F40-34 [2.18±0.35 cm (2013), 2.17±0.32 cm (2014) and 5.40±0.26 cm (2015)] (Figure 1B). However, F12-38 and F40-36 are considered highly susceptible to ICS 40 (moderately sensitive).

Heterosis and heterobeltiosis
Six days after infection of the leaves, the hybrid F12 and F40 populations showed a positive heterosis effect of 86.66% (Table 1) on a continuous way (during the years 2013, 2014 and 2015). Likewise, genotypes F12-33 (+11.95) and F12-35 (+0.63), which showed a negative heterosis effect in the year 2013, on the other hand, they were again effective (positive heterosis) in the years 2014 and 2015.Positive heterobeltiosis was manifested by a performance of hybrid genotypes superior to that of the better parent (SCA12). Only 85% of hybrid genotypes displayed negative values for the parameter studied after three years of evaluation (Table 1). Individuals F12-35 (-7.55) and F40-39 (-0.20) in 2015 showed that their positive heterobeltiosis is very small because their value was nearby to 0. Moreover, in hybrids F12-38 and F40-36 a continuously negative heterobeltiotis (periods 2013, 2014 and 2015) (Table 1).

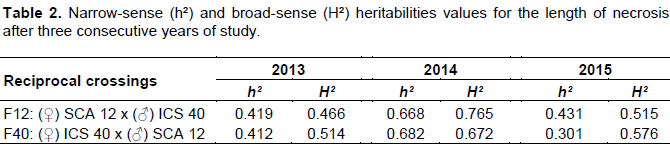
Broad-sense and narrow-sense heritabilities
Narrow-sense and broad-sense heritabilities estimates for necrosis character and biochemical parameters were obtained for the two populations from reciprocal crosses. Values of heritability in 2014 period were h² = 0.686 and H² = 0.765 for F12 and h² = 0.682 and H² = 0.672 for F40. These values were the highest of the other two years (2013 and 2015) (Table 2). Concerning (additive gene effects) and broad-sense (additive + dominance + epistasis effects), the values obtained in the (♀) SCA12 x (♂) ICS40 family (h² = 0.431 and H²= 0.515 for 2015) was not significant to that obtained in the (♀) ICS40 x (♂) SCA12 (h² = 0.301 and H²= 0.576 for 2015). The observation was the same for the 2013 and 2014 periods (Table 2).

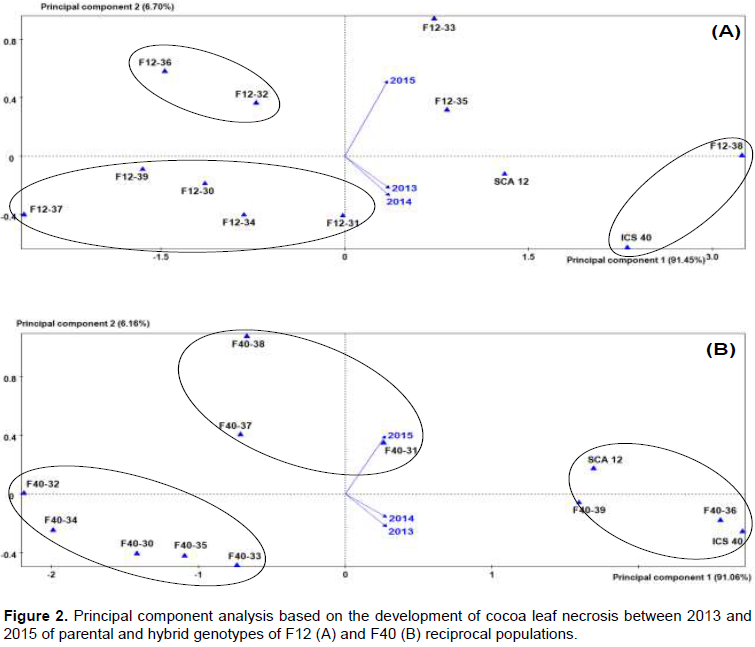
Principal component analysis (PCA) and hierarchical cluster
In the F12 family, the first two principal components (PC) generated from all data represented 98.15% of the total variability of necrosis during three years of study. The year 2015 was the dominant feature of the first axis [PC1 (91.45% of total variability)], while necrosis in the years 2013 and 2014 was characterized as the highest of the second axis [PC2 (6.70% of total variability)] (Figure 2A).The F40 family was dominated by necrosis in 2014 with 91.06% total variability, while necrosis progressed rapidly in the years 2013 and 2015 with a PC2 characterized by a total variability of 6.16% (Figure 2B). However, genotypes F40-36, F40-39, SCA12 and ICS40 which formed a group had higher necrosis sizes in all years 2013, 2014 and 2015.

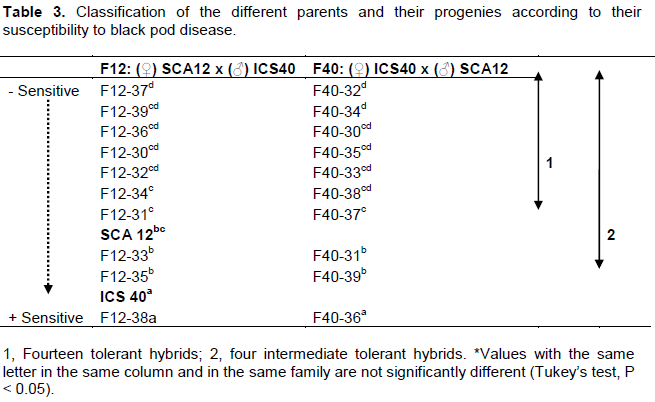
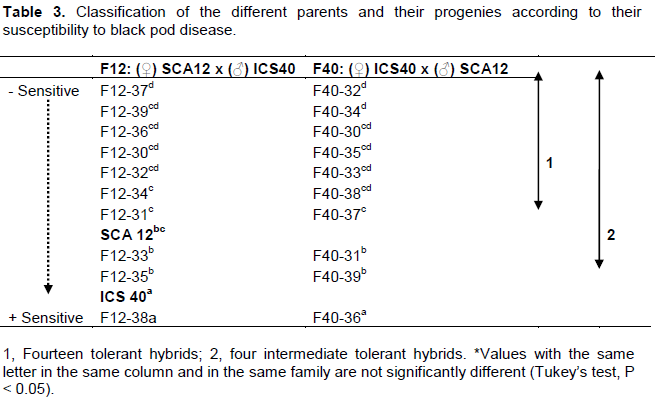
Cluster analysis (5% dissimilarity) also classified genotypes by their affinities with the necrosis length character studied. The first cluster composed of genotypes that shared only 37% similarity among them. The second cluster consisted of two parents (SCA12 and ICS40) and eight hybrids correlated with 37.5% similarities. On the other hand, F12-38 (cluster 3) and F12-37 (cluster 4), differentiated themselves from other genotypes by their behavior (performance for F12-38 and vulnerable for F12-37) (Figure 3). The ranking of parents and their offspring for necrosis length allowed classify more efficient genotypes other than the best parent “SCA12”. F12, F40, families respectively presented 7 (F12-37> F12-39 >F12-36 > F12-30 > F12-32 >F12-34 > F12-31) and (F40-32 > F40-34 >F40-30 > F40-35 > F40-33 > F40-38 > F40-37) genotypes with the susceptibility level less than the best parent (Table 3). Intermediate genotypes with the susceptibility level less than the sensitive moderately parent “ICS40” presented 2 (F12-33 > F12-35) and 2 (F40-31 > F40-39) (Table 3).

Phenols and flavonoids
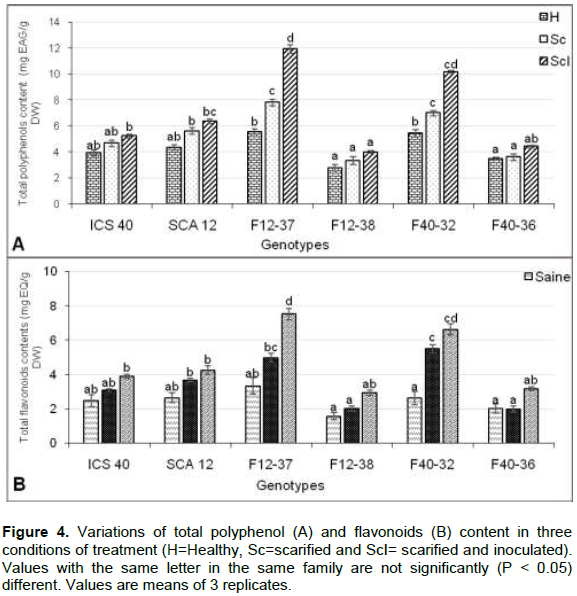
Biochemical component in the leaves of hybrids and parents inoculated with P. megakarya (Figure 4) showed significant variations among the healthy, scarified, scarified and inoculated, non-inoculated and parents for total polyphenols and flavonoids content between tolerant and susceptible hybrids. Tolerant hybrids, F12-37 and F40-32, had accumulated relatively higher TPP (5.44 and 11.91 mg EAG/g DW) and TF content (2.63 and 7.53 mg EAG/g DW), respectively. This was contrary to susceptible hybrids recorded low content (Figure 4). However, Table 4 shows the heritability values in the narrow-sense [h² was low (varying between 0.019 to0.063)] and in the broad-sense [H² was high (ranging from 0.476 to 0.562)] of the accumulation of biochemical compounds of two reciprocal populations.


Correlation coefficients
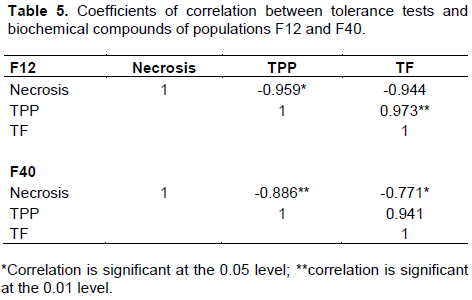
The trials that were carried out revealed significant correlation in the two families studied. The significant negative correlation was noticed between the necrosis length and biochemical component [F12: TPP (r = -0.959, P<0.05) and TF (r = -0.944) and F40: TPP (r = -0.886, P<0.01) and TF (r = -0.771, P<0.05)] (Table 5). Besides, a significant positive correlation between TPP and TF (F12 and F40 populations, P<0.05) was recorded (Table 5).

In this study, we attempted to estimate heritability (narrow-sense and broad-sense), heterosis (mid-parent and better-parent) and understand the role of metabolites in the interaction T. cacao L./P. megakarya. The length of necrosis and the biochemical compounds (TPP and TF) were determined in the leaves of the parental and hybrid genotypes of T. cocoa L. In order to achieve this objective, the production of the plant material (grafting and hand-pollination) was the fundamental element of this work. The experiment and field work took three consecutive years of assessment of the tolerance level of F12 and F40 populations in nurseries and fields. Necrotic lesion size was observed 48 h after inoculation of the fungal mycelia. This confirmed that this duration is necessary for the interaction between the two entities and the beginning of resistance of cocoa tissues. The length of necrosis was favored by the vascularization of midrib which facilitates the progression of the disease. These results have been already obtained by Effa et al. (2016) on the leaves attached to the plant (cocoa) in nursery and field. This result supports the findings of Nana et al. (2011), after the evaluation of necrotic lesions on cocoa pod cortex (SNK10 and SNK413).
Heterosis [mid-parent (86.66% for F12-40)] and heterobeltiosis [better-parent (85% for F12-40)] were evaluated for the development of necrosis character during three cocoa seasons (2013, 2014 and 2015), exhibited hybrid vigor, indicating good general aptitude for the combination of parents in reciprocal crosses (F12: ♀SCA12 × ICS40, F40: ♀ICS40 × â™‚SCA12). From this, it is therefore suggested that these individuals demonstrate hybrid vigor, which would imply the presence of the additive and dominant gene effect in the transmission of character (Djocgoue et al., 2006). The results of the present investigation can be linked to those obtained previously by Manga et al. (2016). Such results show that tolerance to P. megakarya is controlled by the action of additive, dominant and epistatic genes. However, the role of dominant gene effect is more significant than that of additive gene effect. These observations corroborate those obtained by Ondobo et al. (2017) on the cocoa tree. Mohammadi et al. (2011) working on wheat, showed that the involvement of genes with additive and dominant effects in the estimation of heterosis (HPM and HMP) could be important in the selection of varieties.
These results matched with those of Djocgoue et al. (2010) and Manga et al. (2016), which showed a variation of strongly inheritable characters (h² > 0.4). The work of Ram and Jabeen (2016) have shown that high values in a narrow sense heritability for a particular trait indicate that genes are largely controlled in an additive model in Abelmoschus esculentus L. Moench and Zea mays L., respectively. In the studied character, the absence of a significant difference between the heritability values from reciprocal crossing portrays the absence of maternal heritability. This finding suggested that the transmission of this character would not be cytoplasmic but nuclear (Ondobo et al., 2017). In fact, nuclear transmission of characters involved genetic additive effects. Recent studies by Parikh et al. (2016) showed that additive genetic effects are more important than non-additive effects (dominance and epistasis) for powdery mildew resistance in wheat. The high heritability values reported in this study suggest that most of the phenotypic variance in Bpd resistance observed in the two reciprocal crosses is attributable to genetic effects. Similar results were observed for quantitative traits in tobacco (Aleksoska and Aleksoski, 2012), where it was concluded that high broad-sense and narrow-sense heritability were indicative of traits being controlled by genetic factors as opposed to environmental effects.
Estimates of heritabilities for broad-sense and narrow-sense are useful tools to predict the genetic gain resulting from a specific reciprocal crossing. However, general combining abilities (GCA) are predominant for intra- and inter-family selections. This means that the tolerance character is essentially transmitted additively (Ondobo et al., 2017). Principal component analysis (PCA) and dendrogram based on necrosis length was used to categorize all the families; each group consists of similar individuals characterized by low and high length of necrosis. The tolerant hybrids characterized by low and intermediate length of necrosis were considered as elites. This result confirms a good aptitude (cross) towards parental gene combinations (Cilas et al., 2004). Plant biochemical constituents have received considerable attention in relation to disease resistance. Most plants synthesize toxic compounds such as phenols and flavonoids during normal development, and their role in the resistance mechanism has been reported earlier by many authors (Nyadanu et al., 2013; Manga et al., 2016). These can be used as markers for selection of tolerance genotypes. Tolerant plants, when subjected to biotic stress, showed elevated levels of free phenolics and contained more lignin. Their role in resistance mechanisms was previously reported by Effa et al. (2016).
These results are consistent with those of Ondobo et al. (2017), which emphasize that these compounds act as barriers against pathogen invasion and hence constitute part of host resistance mechanisms. This accumulation of flavonoids supported the assertion that these compounds are a major class of polyphenols involved in the resistance of cocoa against
P. megakarya. Among these flavonoids, flavan-3-ol units are a major class with regards to the reaction of cocoa to stress (Effa et al., 2016). This is supported by the findings of Djocgoue et al. (2007) who reported that qualitative analysis of phenolics in leaves of cocoa showed a higher accumulation of the flavonoids (luteolin and apigenin derivatives) and some hydroxycinnamic acid derivatives. The significant negative correlation between metabolites (TPP and TF) and length of necrosis trait reflects the importance of the biochemical compounds in tolerance expression. The findings indicate that TPP and TF play a role in disease resistance against
P. megakarya. The results support the findings of many authors. Ngadze et al. (2012) disclosed that potato varieties with a high content of these compounds in tuber tissues can exhibit tolerance to pathogen attack. Nyadanu et al. (2013) suggested that total polyphenols,
soluble sugars, insoluble sugars, nitrogen, proteins, flavonoid, tannins and lignin are involved in resistance of cocoa to black pod disease caused by
P. palmivora and
P. megakarya.
The overall evaluation of 20 genotypes cocoa hybrids led to identification of the tolerant hybrids with about 85.83% of hybrid vigor. Narrow-sense heritability for Bpd tolerance observed in the current study indicates that additive gene effects are more important than non-additive effects (dominance and epistasis) for Bpd tolerance.
The authors have not declared any conflict of interests.
The authors have not declared any conflict of interests.
REFERENCES
|
Aleksoska AK, Aleksoski J (2012). Heritability of quantitative traits in F1 and F2 progenies of some domestic and foreign tobacco varieties. Agriculturae Conspectus Scientificus 77:207-210.
|
|
|
|
Anonymous (2017). ICCO.Quarterly Bulletin of Cocoa Statistics, Vol. XLII, No. 1, Cocoa year 2016/17, May 2017.
|
|
|
|
Ayoola GA, Ipav SS, Solidiya MO, Adepoju-Bello AA, Coker HAB, Odugbemi TO (2008). Phytochemical screening and free radical scavenging activities of thefruits and leaves of Allanblackia floribunda oliv (Guttiferae). Int. J. Health Res. 2:81-93.
|
|
|
|
Cilas C, Ndounbe-Nkeng M, Bidzanga N, N'goran J (2004). Incidence de la maladie et de résistance sur le terrain. Dans: Cilas C. et Despréaux D. (eds). Amélioration de la résistance des arbres de cacao à la maladie de Phytophthora. Repères, CIRAD, Paris, France. pp. 171-193.
|
|
|
|
Counet C, Ouwerx C, Rosoux D, Collin S (2004). Relationship between Procyanidin and Flavor Contents of Cocoa Liquors from Different Origins. J. Agric. Food Chem. 52:6243-6249.
Crossref
|
|
|
|
Djocgoue PF, Boudjeko T, Mbouobda HD, Nankeu DJ, El Hadrami I, Omokolo DN (2007). Heritability of phenols in the resistance of Theobroma cacao against Phytophthora megakarya, the causal agent of Black Pod Disease. J. Phytopathol. 155:519-525.
Crossref
|
|
|
|
Djocgoue PF, Boudjeko T, Nankeu DJ, Efombagn MIB, Nyasse S, Omokolo DN (2006). Comparative assessment of the resistance of Cocoa (Theobroma cacao L.) progenies from SNK10 x SNK413; ICS84 x ICS95 to Phytophthora megakarya in Cameroon by measuring size of necrotic lesion along the midrib. Plant Pathol. J. 5(3):329-33.
Crossref
|
|
|
|
Djocgoue PF, Mbouobda HD, Boudjeko T, Effa OP, Omokolo ND (2011). Amino acids, carbohydrates and heritability of resistance in the Theobromacacao/Phythophthora megakarya interaction. Phytopathol. Mediterr. 50:370-383.
|
|
|
|
Djocgoue PF, Simo C, Mbouobda HD, Boudjeko T, Nankeu DJ, Omokolo ND (2010). Assessment and heritability of productivity and tolerance level to Phytophthoramegakarya in two hybrid populations of Theobroma cacao. J. Plant Pathol. 92:607-617.
|
|
|
|
Effa OP, Akoa SP, Ondobo ML, Djocgoue PF (2016). Assessment of tolerance level to Phytophthora megakarya in four hybrid populations of Theobroma cacao L. Int. J. Biosci. 3:53-64.
|
|
|
|
Falconer DS, Mackay TFC (1996). Introduction to Quantitative Genetics, Ed 4. Longmans Green, Harlow, Essex, UK. P 125.
|
|
|
|
Georgé S, Brat P, Alter P, Amiot MJ (2005). Rapid determination of polyphenols and vitamin C in plant-derived products. J. Agric. Food Chem. 53:1370-1373.
Crossref
|
|
|
|
Manga NJ, Effa OP, Ondobo ML, Djoko KJC, Djocgoue PF (2016). Heritability of the tolerance to Phytophthora megakarya Bras. and Grif. of Theobroma cacao L. in terms of their necrosis length, phenolic contents and activity of enzymes. Int. J. Bios. 8(5):249-261.
|
|
|
|
Mohammadi M, Ramezanpour S, Navabpour S, Soltanloo H, Kalateharabiand M, Kia S (2011). Genetic analysis and heritabilities of resistance to Mycosphaerella graminicola in wheat. Crop Breed. J. 2:35-42.
|
|
|
|
Nana WL, Tchameni NS, Fokom R, Etoa FX, Debost M (2011). Flavonoïd Compounds Synthesis by Cocoa Fruits (Theobroma cacao L.) in Response to Phytophthora megakarya Infection. Res. J. Agric. Biol. Sci. 7:335-342.
|
|
|
|
Ngadze E, Icishahayo D, Coutinho TA, Van der Waals JE (2012). Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid, and total soluble phenols in resistance of potatoes to soft rot. Plant Dis. 96:186-192.
Crossref
|
|
|
|
Nyadanu D, Akromah R, Adomako B, Kwoseh C, Lowor ST, Dzahini-Obatey H (2013). Biochemical Mechanisms of Resistance to Black Pod Disease in Cocoa (Theobroma cacao L.). Am. J. Biochem. Mol. Biol. 3:20-37.
Crossref
|
|
|
|
Nyasse S (1997). Etude de la diversité de Phytophthora megakarya et caractérisation de la résistance du cacaoyer (Theobroma cacao L.) à cet agent pathogène. Thèse de doctorat, Institut National de Polytechnique de Toulouse (France) 133 p.
|
|
|
|
Ondobo ML, Effa OP, Manga NJ, Djoko KJC, Djocgoue PF, Omokolo ND (2017). Breeding of parental and tolerant hybrids of Theobroma cacao L. to Phytophthora megakarya Bras. and Griff. Int. J. Plant Physiol. Biochem. 9:9-21.
Crossref
|
|
|
|
Parikh L, Mmbaga MT, Kodati S, Blair M, Hui D, Meru G (2016). Broad-sense heritability and genetic gain for powdery mildew resistance in multiple pseudo-F2 populations of flowering dogwoods (Cornus florida L.). Sci. Hortic. 213:216-221.
Crossref
|
|
|
|
Ram RV, Jabeen F (2016). Narrow sense heritability, correlation and path analysis in maize (Zea mays L.). SABRAO J. Breed. Genet. 48:120-126.
|
|
|
|
Tang F, Xiao W (2013). Genetic effects and heterosis of within-boll yield components in upland cotton (Gossypium hirsutum L.). Euphytica 194:41-51.
Crossref
|
|
|
|
Wieslaw O, Chang YL, Antoni WJ, Keith RP (1988). Identification of some phenolic compounds in apples. J. Agric. Food Chem. 36:430-432.
Crossref
|
|
|
|
Zahour A (1992). Eléments d'Amélioration Génétique des Plantes. Ed. Actes: Rabat 232 p.
|