Full Length Research Paper

ABSTRACT
Woodiness disease is the most important disorder of passion fruit worldwide. The causal agent in Brazil is the Cowpea aphid-borne mosaic virus (CABMV), and despite the economic relevance of passion fruit for agriculture there have been recently very few studies about this virus in Brazil and worldwide. This work reveals the phylogenetic relationships of 10 newly identified CABMV isolates from Bahia State, the region where CABMV was first identified (in that time reported as PWV) in South America before its outbreak. The coat protein of 10 CABMV isolates (CABMV-Lns1 - CABMV-Lns10) from Livramento de Nossa Senhora Country, Bahia State, were sequenced and presented very close identity between themselves (nucleotide: 97 to 99%, amino acid: 95 to 100%). They are phylogenetically closely related to Brazilian CABMV, however forming an isolated cluster within the Brazilian clade. According to previous evidences, our data demonstrate that CABMV-Lns are more closely related to isolates from Southern rather than from Northern Africa. Other two isolates from Bahia State clustered separately from CABMV-Lns, but together with isolates from other Brazilian regions thus suggesting that CABMV-Lns are a strain likely restricted to Bahia. The characterization of new populations of CABMV enables greater resolution of the evolution of viruses causing woodiness disease in passion fruit vines. Our data shed light on an as yet unexplored population of CABMV in Brazil and contributes to the understanding of its evolutionary history.
Key words: Passion fruit, Livramento de Nossa Senhora, Bahia, phylogenetics, woodiness disease.
Abbreviation: CABMV, Cowpea aphid-borne mosaic virus; PWV, passion fruit woodiness virus; EAPV, East asia passiflora virus.INTRODUCTION
The passion fruit is one of the most important crops in Brazil. It belongs to Passiflora genus and Passifloraceae family. Viral diseases are the most severe in terms of passion fruit yield loss (Sokhandan et al., 1997; Gibbs et al., 2008b). This is mainly due to the lack of both quick and accurate diagnosis as well as effective methods for disease control (Andrade and Pio-Ribeiro, 2001). In Brazil and Africa woodiness disease is caused by Cowpea aphid-borne mosaic virus (CABMV) (McKern et al., 1994; Nascimento et al., 2006; Maciel et al., 2009). This is in contrast to Australia where the passion fruit woodiness virus (PWV) is the most common causal agent (Sokhandan et al., 1997) and also to Asia where the same disease is caused by the East asia passiflora virus (EAPV) (Iwai et al., 2006). Its genus Potyvirus, family Potyviridae, contains more agriculturally important plant viruses than any other (Adams et al., 2005). Although in the past 15 years molecular studies about Brazilian CABMV have been carried out (Novaes and Rezende, 2003; Nascimento et al., 2004; Barros et al., 2011; Cerqueira-Silva et al., 2012; Nicolini et al., 2012), surprisingly none of them has phylogenetically investigated the virus from the region of largest passion fruit production in Brazil, the Livramento de Nossa Senhora County. Exploring the molecular variation of the coat protein (CP) in CABMV helps to better understand variability among strains, isolates and related species, to elucidate the evolutionary history of CABMV as well as assists to shed light on disease outbreaks in the course of passion fruit dispersion (Iwai et al., 2006; Gibbs et al., 2008a; Gibbs et al., 2008b). Bahia State is the place where passion fruit woodiness disease was firstly identified in Brazil during the 70’s (Chagas et al., 1981) before its dissemination to most of the countries’ fields. This led us to hypothesize whether Bahia State might host phylogenetically different isolates than the rest of Brazil that could be due to genomic mutation, recombination with other viral species, selective pressure or other evolutionary events. In this study we report a phylogenetic characterization of a population of CABMV from the State of Bahia, in comparison to other viruses causing fruit woodiness disease in passion fruit vines around the world.
MATERIALS AND METHODS
Virus and plant resources
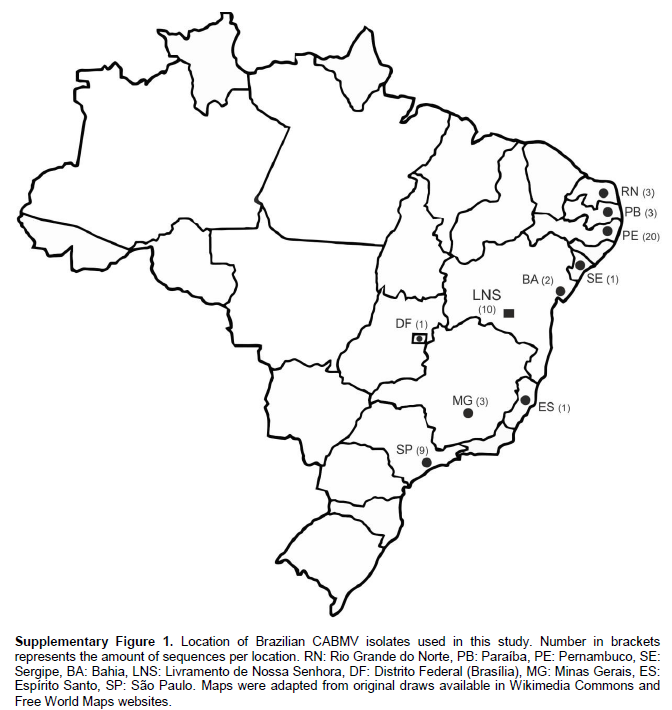
Ten CABMV isolates were collected from different passion fruit crop fields in the farming region of Livramento de Nossa Senhora Country, Bahia State, Brazil (13° 38′ 34″ S, 41° 50′ 27″ W). Ground extract from symptomatic leaves was used for inoculation on healthy passion fruit plants for maintenance, following the method described by Novaes and Rezende (2003). The 10 CABMV isolates here reported were named: CABMV-Lns1 to CABMV-Lns10. Several sequences from passion fruit woodiness disease-related viruses were acquired from public databases (Supplementary Table 1 and Figure 1).
RNA extraction, PCR, cloning and sequencing
RNA extraction was performed following the Arabidopsis Functional Genomics Consortium Trizol protocol (Arabidopsis Functional Genomics Consortium – AFGC, 2013). cDNA synthesis and PCR (Promega), DNA purification (PCR Purification Kit – NORGEN, Wizard SV Gel and PCR Clean-Up System – Promega) and cloning (pGEM-T Easy Vector – Promega, Max Efficiency DH5α Competent Cell – Invitrogen) were performed as essentially as described in the manufacturer’s protocol. Newly designed primers for both cDNA synthesis and PCR were: CABMV8364-F (5’-CCTTTCCTTCTACGATG-3’), CABMV9389-R (5’-CAACCGGGGTATGGCCTC-3’), CABMV8359–F (5’-GGCATCCTTTCCTTCTATG-3’) and CABMV9402–R (5’-GGCATCCTTTCCTTCTATG-3’).
Sequencing and phylogenetic analysis
Sequencing of at least three independent clones from each isolate was performed twice by Macrogen (South Korea). The sequences were manipulated using DNA Baser Sequence Assembler v2 (DNA Baser Sequence Assembler v2.x, 2010). Phylogenetic tree was constructed using MEGA5 (Tamura et al., 2011). For nucleotide-based tree, Neighbor-Joining (NJ) and BioNJ pairwise distances matrix were estimated by maximum composite likelihood (MLC), estimated under bootstrap values higher than 70% (1,000 replications).
RESULTS
Analysis of coat protein sequence
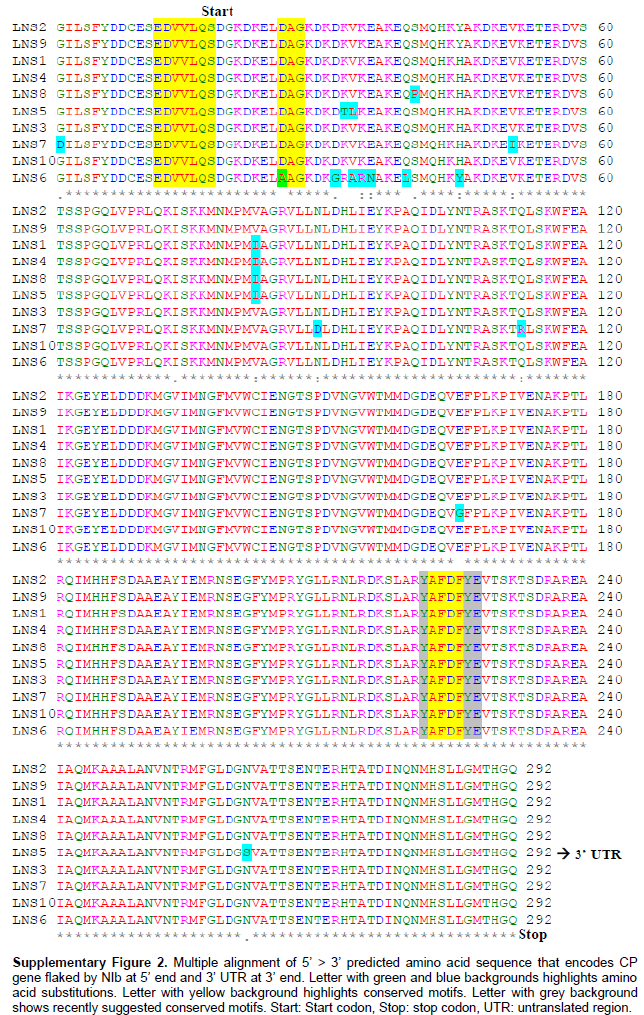
Complete CP coding sequence containing 828 nucleotides and 275 amino acids were identified in all ten CABMV-Lns isolates, in agreement with the sequenced CABMV genomes (Mlotshwa et al., 2002; Barros et al., 2011). Very little variation was found amongst the nucleotide sequence of CABMV-Lns isolates (identity from 97 to 99%) (Table 1). Comparison of CABMV-Lns and other CABMV isolates showed sequence identity varying from 78 to 94%, where the lowest identity was found with CABMV-Mor, CABMV-Iba and CABMV-Monguno isolates (78%) and the highest with CABMV-Brs isolate (94%). The identity of amino acid sequences amongst CABMV-Lns isolates ranged from 95 to 100% identity (Table 1). When compared to other CABMV isolates, it ranged from 80 to 96%, when the least identity was found with CABMV-Bnt isolate (80%) and the highest with CABMV-TC1 isolate (96%). The CP conserved motifs of potyviruses were entirely conserved in nearly all CABMV-Lns isolates, with exception of CABMV-Lns6 which has a substitution of aspartic acid by an alanine residue at the DAG motif (Supplementary Figure 2).
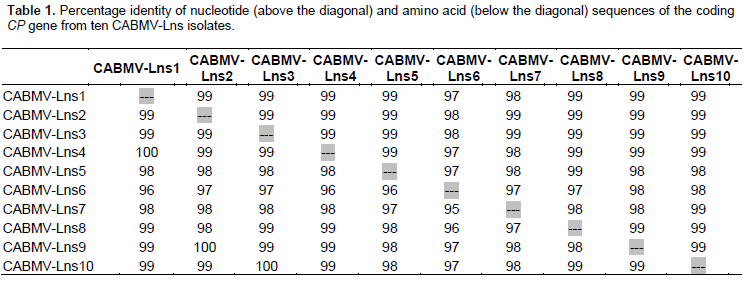
Phylogenetic profile suggests CABMV-Lns as a strain restricted to Bahia State
Maximum-likelihood genealogies of nucleotide sequences of the CP demonstrate that all CABMV isolates clustered within a monophyletic clade that comprehends sequences from Brazil and Africa. CABMV-Lns isolates formed an isolated cluster inside the CABMV mono-phyletic clade (Figure 1). Interestingly they clustered separately from CABMV-Jgr and CABMV-Itb isolates also identified in Bahia State, which are more closely related to isolates from the States of Pernambuco and Paraíba, located around 1,200 km and 1,350 km away from Livramento de Nossa Senhora, respectively. PWV and EAPV (Iwai et al., 2006), which are well known different species from CABMV, formed a distinct clade also separately from CABMV-Lns (Figure 1).
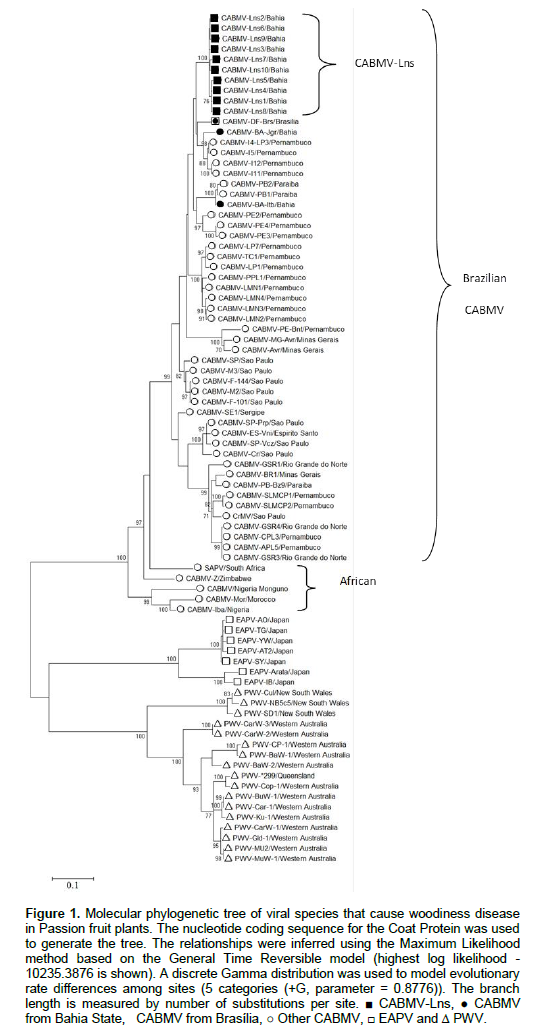
DISCUSSION
The high identity of nucleotide and amino acid sequences amongst the ten CABMV-Lns isolates is strong evidence that they belong to the same strain of CABMV. This information is of great relevance for purposes of disease control, once most of the attempts to overcome woodiness disease in passion fruit such as premunization with mild strains of CABMV/PWV or pathogen-derived resistance have been hindered by variant strains of the same virus (Novaes and Rezende, 2003; Alfenas et al., 2005, Trevisan et al., 2006; Cerqueira-Silva et al., 2014). On the other hand, CABMV-Lns clustered separately from the isolates CABMV-Jgr and CABMV-Itb, which were also collected in Bahia State. As CABMV-Jgr and CABMV-Itb clustered together with isolates from rather far regions, the data suggest that CABMV-Lns are likely a strain restricted to Bahia State. This finding proposes either a single introduction of this strain to the area or a constraint in dissemination of this strain to other places by plant material, since there is no evidence that the virus is seed-transmitted in Passion fruit. According to the demarcation criteria for the Potyvirus genus (King et al., 2011), our data support the hypothesis that CABMV-Lns isolates are a different strain from CABMV-Prp, CABMV-Cr, CABMV-Vni, CABMV-Z, CABMV-Iba, CABMV-Monguno, CABMV-Mor, CABMV-MG-Avr, CABMV-PE-Bnt (80 to 89% amino acid identity). Additionally, our data confirm the higher sequence identity and closer phylo-genetic relationship of CABMV-Lns with Southern African CABMV rather than isolates from the North of Africa, which was also previously suggested (Nascimento et al., 2006). The slave trade between Africa and South America is the likely event for introduction of CABMV in Brazil (Gibbs et al., 2008b), by which infected biological material could have been transported from the South rather than the North of Africa. However, we are aware that the limited number of CABMV sequences currently available, associated to the little information about African CABMV infecting Passion fruit plants, significantly narrow down the possibilities of drawing more precise conclusion on this regard. With reference to the important CP conserved motifs of potyviruses, the DAG motif is related to transmissibility of potyviruses by insects (Abdullah et al., 2009). Isolate CABMV-Lns6 shows a substitution of aspartic acid by alanine it to the DAG motif (Supplementary Figure 1). Mutations in this region significantly reduced transmissibility by insects in previous studies (Abdullah et al., 2009; López-Moya et al., 1999). However, unfortunately transmissibility experiments to investigate the consequences of the mutated DAG motif on the transmissibility capacity of CABMV-Lns6 were not suitable before the submission of this work.
CONCLUSION
Our data precisely demonstrate that CABMV-Lns belong also to CABMV species. These isolates can be classified as a strain of CABMV thus far occurring only in Bahia State and phylogenetically more distant from other isolates occurring in the same region. In addition and accordingly to previous reports, they are more closely related to Southern rather than Northern African CABMV. The exploitation of a larger number of viral samples from other as yet unexplored places in the promising State of Bahia will help to shed light on the events contributing to the observed regional isolation of CABMV-Lns. The whole genome sequencing of CABMV-Lns is ongoing and will allow us to comprehend their evolutionary history with regard to other related viral species around the world.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGEMENTS
This study consists one chapter of the first author’s Master of Science Thesis, funded by the Coordination for the Improvement of Higher Level Personnel (CAPES) in the Graduate Program of Plant Biotechnology at the Federal University of Lavras, MG, Brazil. It was partially funded by the Research Support Foundation of Bahia State (FAPESB, 0852/2006, Undergraduate Research Program-PIC/UESB Scholarship to first author) at the Southwest Bahia State University (UESB). We acknowledge the assistance of Prof. Leandro M. de Freitas (UFBA) for phylogenetic analyses, Allan S. Pereira and Claudio Benicio C. Silva (UESB) for field work and Carlos Roberto Torres (UFLA) for plant/virus maintenance. We also thank Dr. Scott Sinclair for considerably improving the text and the anonymous reviewers for their careful reading of our manuscript and their insightful comments and suggestions.
REFERENCES
|
Abdullah N, Ismail I, Pillai V, Abdullah R, Sharifudin SA (2009). Nucleotide sequence of the coat protein of the Malasian passiflora virus and its 3' non-coding region. Am. J. Appl. Sci. 6(9): 1633-1636. |
|
|
Adams MJ, Antoniw JF, Fauquet CM (2005). Molecular criteria for genus and species discrimination within the family Potyviridae. Arch. Virol. 150(3):459-479. |
|
|
Alfenas PF, Braz ASK, Torres LB, Santana EN, Verônica A, Nascimento S, Zerbini FM (2005). Transgenic Passionfruit expressing RNA derived from Cowpea aphid-borne mosaic virus is resistant to passionfruit woodiness disease. Fitopatol. Brasil. 30(1):33-38. |
|
|
Andrade GP, Pio-Ribeiro G (2001). Estratégias e métodos aplicados ao controle de fitoviroses. In Michereff SJ and Barros (Ed.) Proteção de plantas na agricultura sustentável, v. 3, R. Universidade Federal de Pernambuco, Recife, PE, Brazil, pp. 171-181. |
|
|
Arabidopsis Functional Genomics Consortium - AFGC (2013). Total RNA isolation. AFGC Website. |
|
|
Barros DR, Alfenas-Zerbini P, Beserra JEA, Antunes TFS, Zerbini, FM (2011). Comparative analysis of the genomes of two isolates of cowpea aphid-borne mosaic virus (CABMV) obtained from different hosts. Arch. Virol. 156(6):1085-1091. |
|
|
Boxtel J, Thomas C, Maule AJ (2000) Phylogenetic analysis of two potyvirus pathogens of commercial cowpea lines: implications for obtaining pathogen-derived resistance. Virus Genes 20:71-77. |
|
|
Brand, RJ, Burger, JT & Rybicld, EP (1993) Cloning, sequencing, and expression in Escherichia coli of the coat protein gene of a new potyvirus infecting South African Passiflora. Archives of Virology128:29-41. |
|
|
Brand RJ, Burger JT, Rybicld EP (1993). Cloning, sequencing, and expression in Escherichia coli of the coat protein gene of a new potyvirus infecting South African Passiflora. Arch. Virol. 128(1-2): 29-41. |
|
|
Cerqueira-Silva CBM, Conceição LDHCS, Souza AP, Corrêa RX (2014). A history of passion fruit woodiness disease with emphasis on the current situation in Brazil and prospects for Brazilian passion fruit cultivation. Eur. J. Plant Pathol. 139(2): 255-264. |
|
|
Cerqueira-Silva CBM, Melo JRF, Corrêa RX, Oliveira AC (2012). Selection of pathometric variables to assess resistance and infectivity in the passion fruit woodiness pathosystem. Eur. J. Plant Pathol. 134(3): 489-495. |
|
|
Chagas CM, Kitajima, EW, Lin, MT (1981). Grave moléstia em maracujá amarelo (Passiflora edulis f. flavicarpa) no Estado da Bahia causada por um isolado do vírus do "woodiness" do maracujá. Fitopatol. Bras. 6(2): 259-268. 1981. |
|
|
Coutts, BA, Kehoe, MA, Webster, CG, Wylie, SJ & Jones, RA (2011) Indigenous and introduced potyviruses of legumes and Passiflora spp. from Australia: biological properties and comparison of coat protein nucleotide sequences. Archives of Virology 156:1757-1774. |
|
|
DNA Baser Sequence Assembler v2.x (2010). HeracleSoftware. |
|
|
Freitas DS, Maia IG, Arruda, P, Vega J (2002) Molecular characterization and evolutionary relationships of a potyvirus infecting Crotalaria in Brazil. Archives of Virology 147:411-417. |
|
|
Fukumoto T, Nakamura M, Ohshima K, Iwai H (2012b) Genetic structure and variability of East Asian Passiflora virus population in Amami-O-shima, Japan. J. Phytopathol. 160:404-411. |
|
|
Fukumoto T, Nakamura M, Rikitake M, Iwai, H (2012a) Molecular characterization and specific detection of two genetically distinguishable strains of East Asian Passiflora virus (EAPV) and their distribution in southern Japan. Virus Genes 44:141-148. |
|
|
Gibbs AJ, Mackenzie AM, Wei KJ, Gibbs MJ (2008a). The potyviruses of Australia. Arch. Virol. 153(8):1411-1420. |
|
|
Gibbs AJ, Ohshima K, Phillips MJ, Gibbs MJ (2008b). The prehistory of potyviruses: their initial radiation was during the dawn of agriculture. PloS One 3: e2523. |
|
|
Gioria, R (2003) Caracterização sorológica, biológica e molecular de uma estirpe do Passion fruit woodiness virus que infecta sistemicamente algumas cucurbitáceas. Dissertation, Escola Superior de Agricultura Luiz de Queiroz. |
|
|
Iwai H, Yamashita Y, Nishi N, Nakamura M (2006). The potyvirus associated with the dappled fruit of Passiflora edulis in Kagoshima prefecture, Japan is the third strain of the proposed new species East asian passiflora virus (EAPV) phylogenetically distinguished from strains of Passion fruit woodiness virus. Arch. Virol. 151(4): 811–818. |
|
|
King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (2011). Virus Taxonomy - Ninth Report of the International Committee on Taxonomy of Viruses. Elsevier/Academic Press; London, United Kingdom. |
|
|
Kitjima, EW, de Alcântara, BK, Madureira, PM, Alfenas-Zerbini, P, Rezende, JAM & Zerbini, FM (2008) A mosaic of beach bean (Canavalia rosea) caused by an isolate of Cowpea aphid-borne mosaic virus (CABMV) in Brazil. Archives of Virology 153:743-747. |
|
|
López-Moya JJ, Wang RY, Pirone TP (1999). Context of the coat protein DAG motif affects potyvirus transmissibility by aphids. J. Gen. Virol. 80(12): 3281-3288. |
|
|
Maciel SC, Nakano DH, Rezende JAM, Vieira MLC (2009). Screening of passiflora species for reaction to Cowpea aphid-borne mosaic virus reveals an immune wild species. Sci. Agric. 66(3): 414-418. |
|
|
McKern NM, Strike PM, Barnett OW, Dijkstra J, Shukla DD, Ward CW (1994). Cowpea aphid borne mosaic virus-Morocco and South African Passiflora virus are strains of the same potyvirus. Arch. Virol. 136(1-2):207-217. |
|
|
Mlotshwa S, Verver J, Sithole-Niang I, Kampen TV, Kammen AV, Wellink J (2002). The genomic sequence of Cowpea aphid-borne mosaic virus and its similarities with other potyviruses. Arch. Virol. 147(5): 1043-1052. |
|
|
Nascimento AVS, Santana EN, Braz ASK, Alfenas PF, Pio-Ribeiro G, Andrade GP, Carvalho MG, Zerbini, MF (2006). Cowpea aphid-borne mosaic virus (CABMV) is widespread in passion fruit in Brazil and causes passionfruit woodiness disease. Arch. Virol. 151(9):1797–1809. |
|
|
Nascimento AVS, Souza ARR, Alfenas PF, Andrade GP, Carvalho MG, Pio-Ribeiro G, Zerbini, FM (2004). Análise filogenética de potyvirus causando endurecimento dos frutos do maracujazeiro no Nordeste do Brasil. Fitopatol. Bras. 29(4):378-383. |
|
|
Nicolini C, Rabelo Filho FAC, Resende RO, Andrade GP, Kitajima EW, Pio-Ribeiro G, Nagata T (2012). Possible host adaptation as an evolution factor of Cowpea aphid-borne mosaic virus deduced by coat protein gene analysis. J. Phytopathol. 160(2):82-87. |
|
|
Novaes QS, Rezende JAM (2003). Selected mild strains of Passion fruit woodiness virus (PWV) fail to protect pre-immunized vines in Brazil. Sci. Agric. 60(4):699-708. |
|
|
Novaes, QS, Rezende, JAM (2005) Protection between strains of Passion fruit woodiness virus in sunnhemp. Fitopatologia Brasileira, Brasília 30:307-311. |
|
|
Sokhandan N, Gillings M, Bowyer J (1997). Polymerase chain reaction detection and assessment of genetic variation in New South Wales isolates of Passionfruit woodiness potyvirus. Aust. Plant Pathol. 26(3):155-164. |
|
|
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28(10):2731-2739. |
|
|
Trevisan F, Mendes BMJ, Maciel SC, Vieira MLC, Meletti LMM, Rezende JAM (2006). Resistance to Passion fruit woodiness virus in transgenic Passionflower expressing the virus coat protein gene. Plant Dis. 90(8):1026-1030. |
|
|
Webster CG, Coutts BA, Jones RAC, Jones MGK, Wylie SJ (2007). Virus impact at the interface of an ancient ecosystem and a recent agroecosystem: studies on three legume-infecting potyviruses in the Southwest Australian floristic region. Plant Pathology 56:729-742. |
|
|
Wylie SJ, Jones MG (2011). The complete genome sequence of a Passion fruit woodiness virus isolate from Australia determined using deep sequencing, and its relationship to other potyviruses. Archives of Virology 156:479-482. |
|
APPENDIX
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


