Full Length Research Paper

ABSTRACT
Drought is one of the main constraints facing crops and affecting their production. In a trial to investigate and understand drought effect, differential display experiments were performed where; about 107 DD-fragments have been isolated from wild barley plants subjected to drought treatment. These fragments were categorized according to their gene expression into four categories: 1) up regulated genes; 2) up and down regulated; 3) down regulated and 4) down and up regulated genes. The isolated fragments were cloned and sequenced. Sequence analysis using GeneBank database revealed that some fragments have significant similarities with starch branching enzyme I (sbeI) gene; glycosyl transferase family 1 gene; Retrotransposons Ty3-gypsy subclass; gag-polypeptide of LTR copia-type retroelement gene; IMP cyclohydrolase/phosphoribosylaminoimidazolecarboxamide formyltransferase and unknown proteins. Some other fragments have significant similarity with Triticum aestivum gene for TaAP2-B, complete cds and 3B chromosome, clone BAC TA3B63N2. These results indicate that starch metabolism, transposon elements and purine metabolism might have been affected by drought stress.
Key words: Wild barley, differential display, drought; starch metabolism, purine metabolism.
INTRODUCTION
Barley (Hordeum vulgare L) is an important cereal crops. It is ranked as the fourth crop in the 2007 world wide cereal crops’ ranking. It is one member of the grass family. Barley is used for brewing malts for beer, animal feed and certain distilled beverages, and it is a component of human health foods. From evolution point of view, the scientists have hypothesized that the Hordeum spontaneum is the ancestor plant for all types of barley. Since the stone age, barley has been cultivated and consumed. It can grow in many areas because of its adaptability and durability (Fastnaught, 2001). It has many health benefits such as in heart diseases, diabetes, lower level of cholesterol, levels of blood sugar and also as vitamins and antioxidants (Smith, 2004). Barley is
mainly grown in the north coastal regions of Egypt under rain fed conditions and in the newly reclaimed lands under irrigation. The area of barley production has been increased, especially in the newly reclaimed lands. Barley is a grain cereal that grown in semi-arid areas. These areas are known with harsh environmental conditions such as drought. Barley can be germinated and grown in such sever conditions. The early growth stages are considered as the most susceptible period to drought stress and the big challenge in the plant productivity (Amini, 2013). Despite of all these difficulties, barley can grow in these semi-arid areas like Egypt. According to Stanca et al. (2003), barley gene content has several good gene candidates that enhance the adaptability of plants to live in the wide harsh environmental conditions.
Drought stress is considered as one of the main constraints facing the production of plant crops in the newly reclaimed areas of Egypt. In drought stress, plants suffer wilting from decreased photosynthetic activity, and osmotic fluctuations. Also, they are susceptible during the reproductive stage, reducing yield and fertility (Ashraf, 2010). Many efforts have been made to improve crop production under the conditions of drought stress. Unlike animals, plants cannot escape from environmental stresses that might cause severe damages. Therefore, plants have several adaptation mechanisms that help to tolerate osmotic stress coming from heat, salt and/or drought stresses. These adaptation mechanisms are performed through induction of a plant molecular response (Cattivelli et al., 2011). According to Cattivelli et al. (2011), plant molecular response is performed in three consequence steps; starts with the perception of the external environmental stress followed by signal transduction process to induce selective gene(s) as a final step to accumulate specific protective molecule. Induction of these stress related gene(s) is regulated and controlled by transcriptional elements. These molecular responses improve the ability of the plant to adapt such harsh environmental conditions. In the last 20 years, several stress-related DNA fragments and genes have been fully characterized.
The productivity of the plant is too sensitive to the water insufficiency which result from nearly all types of environmental stresses specially the drought stress. Drought stress might cause yield loss more than any other factor that cause loss in the crop yield. Drought stress has a direct effect on plant development where it affects the water/nutrient relationship, respiration and photosynthesis processes (Farooq et al., 2009). Drought causes reduction in transpiration, photosynthesis, and also other biochemical processes related to development, crop productivity and plant growth (Tiwari et al., 2010). Moreover, drought stress has a direct effect on plant morphology where it causes reduction in the leaf size, prolongation of stem and induction of root proliferation (Farooq et al., 2009). The increase in the osmoprotectants synthesis such as soluble sugar and proline enables plants to survive through drought stress. This increase is considered as one of the common metabolic adaptations (Waseem et al., 2011).
There are several drought-related genes that have been characterized, such as element-binding DNA fragments during drought stress, aquaporin, late embryogenesis abundant proteins (LEA’s) and dehydrin family. According to the newly discovered biotechnological techniques, drought tolerance mechanisms can be enhanced through breeding, marker-assisted techniques and genetic engineering and gene manipulation for drought related gene(s) (Farooq et al., 2009). Many studies have been done on the effect of drought stress on barley growth, production and its adaptation against this stress (Cattivelli et al., 2011; Amini, 2011; Vaezi et al., 2010).
Differential display technique (DD-PCR) is a powerful technique for studying the co-current expression of up and down regulated genes and subsequently isolating those genes. The differential display technique was described for the first time by Liang and Pardee (1992). The differential display technique has been used to study the gene expression in barley, where differentially expressed genes have been isolated from shoots of barley seedlings treated with cold, drought and ABA (Malatrasi et al., 2006).
Also, barley plants were subjected to drought for four days to compare the palea, lemma and awn transcriptoms (Abebe et al., 2010). El-Shehawi et al. (2011) have used the same technique in wheat to study and isolate cDNAs to represent tissue specific genes. They have isolated, cloned and sequenced many tissue specific cDNAs fragments.
Also, Eissa et al. (2007) have isolated, characterized, cloned and sequenced some ESTs from wild barley (H. spontaneum L.) which was subjected to 250 mM NaCl using DD technique. On the other hand, DD-PCR technique has been used in drought stress studies on different plants such as sugarcane culms (Iskandar et al., 2011); the effect of ABA on wild type Vicia villosa (Abou Ali et al., 2010).
The aim of the current study was to investigate the effect of drought stress on wild barley (H. spontaneum L.), and to isolate and identify some drought related gene fragments and candidates from barley treated shoots (under drought stress).
MATERIALS AND METHODS
Plant materials
The wild barley (H. spontaneum L.) seeds were washed for 1 min with 70% ethanol by shaking and then removed, followed by soaking in 20% sodium hypochlorite (commercial Clorox) supplemented with few drops of Tween 20 for 10 min. The seeds were washed several times using sterilized distilled water and then germinated on MS medium (Duchefa Biochemie, M0222.0050) for 8 days. For treatment, seedlings were dehydrated for 0, 3 and 10 h on 3 mm Whatman filter paper at room temperature according to Gao et al. (2008). The control seedlings were left in the MS media. After that, the treated and control seedlings were harvested, and quick frozen in liquid nitrogen, then stored at -80°C until RNA extraction.
Isolation of RNA and differential display-polymerase chain reaction (DD-PCR)
The TriPure Isolation Reagent (Cat. No. 11 667 157 001, Roche- Germany) was used for total RNA isolation from the shoots of drought treated wild barley (H. spontaneum L.) plants (3 and 10 h) and shoots of control barley plants; about 200 mg tissues were used in each RNA extraction. The DD-PCR reactions were described in RNAmap kit (GenHunter Corporation, USA). Each cDNA reaction contained 2 μg total RNA, 2 μM anchor primer (T11C or T11G), 1.0 μl of dNTPs (10 mM), and 1.0 μl MMLV reverse transcriptase (200 U/μl). The reverse transcription (RT) reactions were performed for 50 min at 42°C followed by incubation for 15 min at 70°C. The list of primers sequences is shown in Table 1.
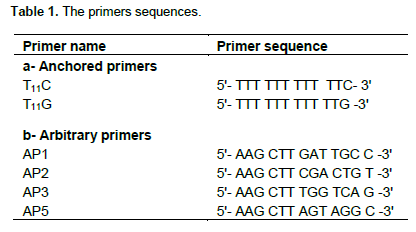
PCR amplification of cDNA
One micro lilter of each cDNA reaction was used for PCR amplification in the presence of 1.0 μl of dNTPs (25 μM), 1.0 μl anchored primer (2 μM), the same as in the RT reaction, 1.0 μl arbitrary primer (2 μM namely AP1, AP2, AP3 or AP5), 1 μl MgCl2 (25 mM) and 0.3 μl Taq DNA polymerase (5 U/ μl) (Fermantas, USA). The PCR program was: 4 min at 94°C (1 cycle); 1 min at 94°C, 2 min at 42°C, 1 min at 72°C (40 cycles); 10 min at 72°C (1 cycle). The products of PCR were loaded on a 6% acrylamide gel after denaturation of PCR products at 90°C for 5 min. The silver staining system (Promega, USA) has been used for gels visualization.
Isolation of DD fragments and cloning
The differentially display bands were cut from the gel and extracted according to Abou Ali et al. (2010). The nomenclature of the isolated fragment was depending on the primers combination that were used in the PCR reaction. For example, the primers combination between anchor primer G and arbitrary primer 3 showed 6 differentially display fragments and the fragment number 1 was named G3-1. This system of nomenclature was followed in all isolated fragments. Then the bands were re-amplified using the same PCR reactions conditions as mentioned before. The pGEM-T easy system (Promega, USA) was used for cloning. Callard et al. (1996) method was used for the plasmid isolation, the modified boiling method. For right colonies screening, EcoR1 digestions were used (Sambrook et al., 1989).
Sequencing of the cDNA insert
Sanger et al. (1977) method was used for sequencing. The basic local alignment search tool (BLAST) in the GeneBank databases was used for comparing nucleotide sequence or the deduced amino acid sequence of each sequenced clones (http://www.ncbi. nlm.nih.gov) according to Altschul et al. (1997). The conserved rejoin alignments were done according to Marchler-Baue et al. (2013).
RESULTS AND DISCUSSION
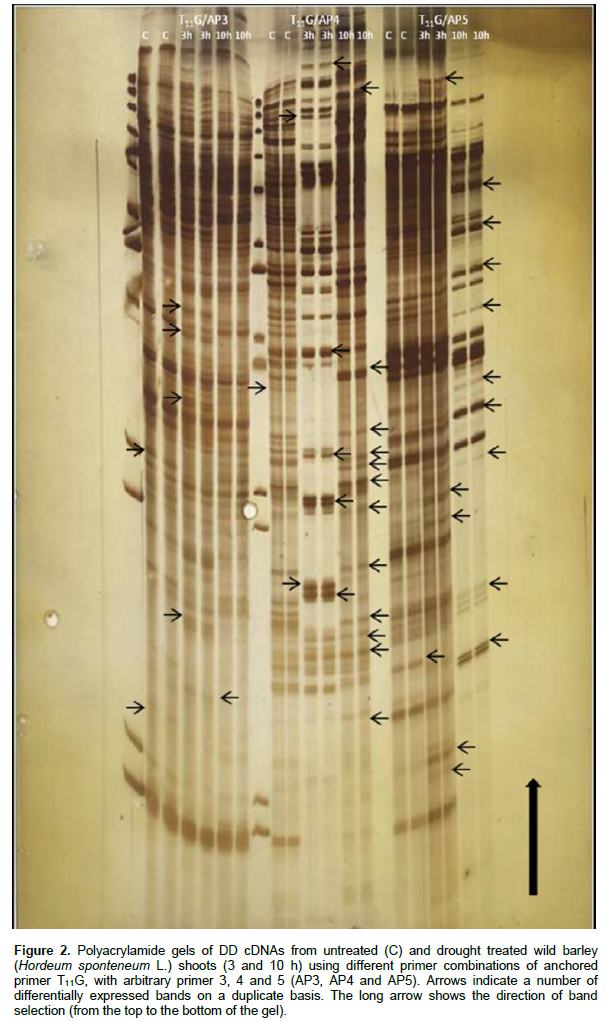
Wild barley (H. spontaneum L) seedlings were subjected to drought stress treatment and subsequently differential mRNA display technique was used to isolate genes and fragments that might be related to drought stress treatment from treated seedlings. The PCR products of different primer pair combinations were loaded on sequencing gels and the cDNA fragments were visualized by silver staining (Figures 1 and 2). One hundred and seven fragments were found to be differentially displayed due to drought treatment of the barley.

Expression patterns of DD fragments
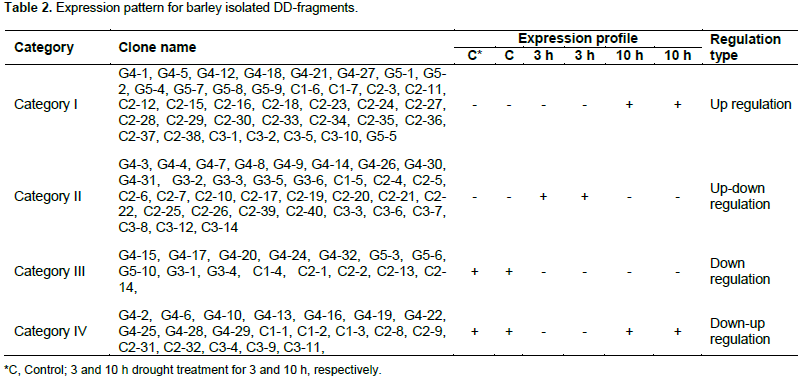
The differentially displayed fragments were isolated and classified into four categories according to their patterns of expression: category i) included 37 DD- fragments which were observed to be up-regulated in seedling shoots in response to drought treatment across the time; category ii) included 34 DD- fragments which were up-regulated after 3 h and then down-regulated in shoots after 10 h of drought treatment; category iii) included 15 DD- fragments which were down regulated in shoots of treated seedlings across time, whereas, category iv) included 21 DD- fragments which down-regulated after 3 h and then up-regulated in shoots after 10 h of drought treatment (Table 2). Thirty (30) differentially display fragments were detected in the primers combination between anchor primer G and arbitrary primer 4 while, the primers combination between anchor primer G and arbitrary primer 5 gave 10 differentially display fragments. Seven differentially display fragments were observed in the primers combination between anchor primer C and arbitrary primer 1 while, the primers combination between anchor primer C and arbitrary primer 2 gave 40 differentially display fragments. Finally, the primers combination between anchor primer C and arbitrary primer 3 gave 14 differentially display fragments.
Cloning and sequencing
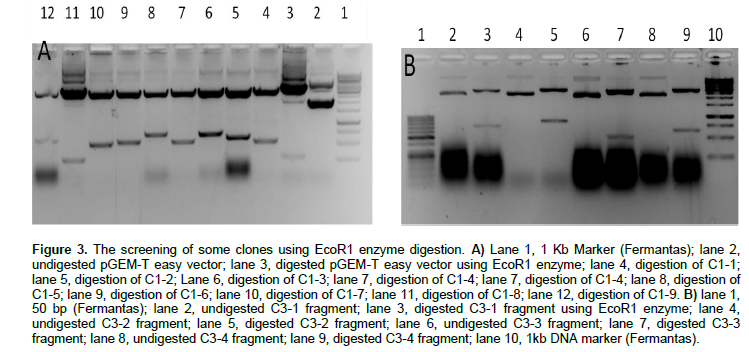
The DD fragments were eluted, and cloned in pGEM-T easy vector. The cloning was confirmed using EcoR1 Digestion (Figure 3A and B). Twenty five (11, 4, 10) cloned fragments have been sequenced. Figure 4 shows the results of the sequencing.




Sequence alignment using Genebank databases
The resulted sequences were subjected to alignment analysis using GeneBank databases and the analysis results revealed significant similarities between the sequences of the isolated fragments and sequences of different genes recorded in the GeneBank databases. Table 3 summarizes the sequence alignment analysis results.
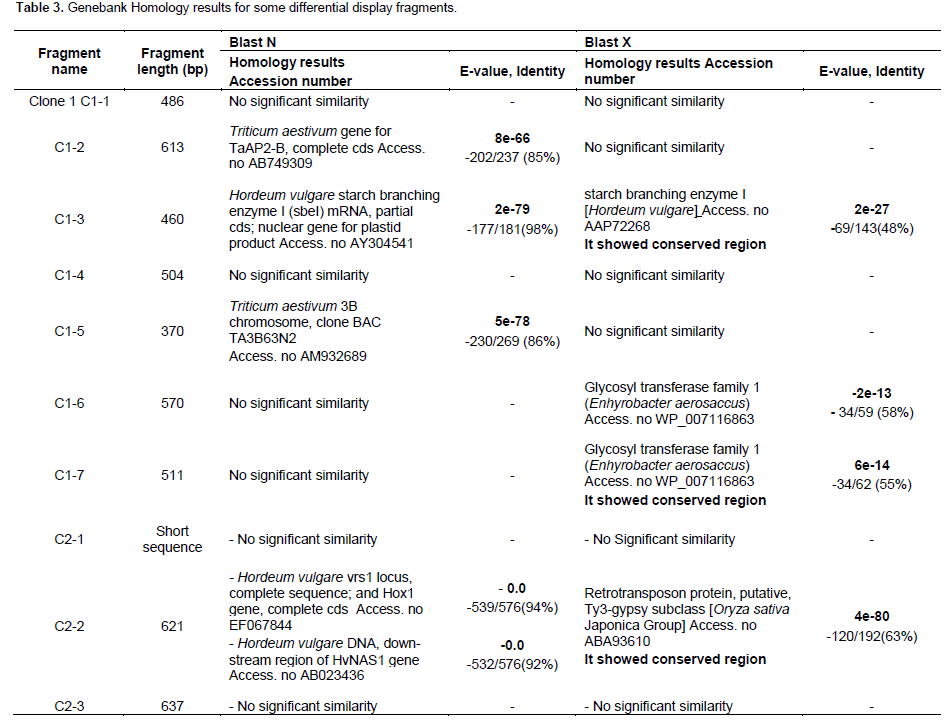
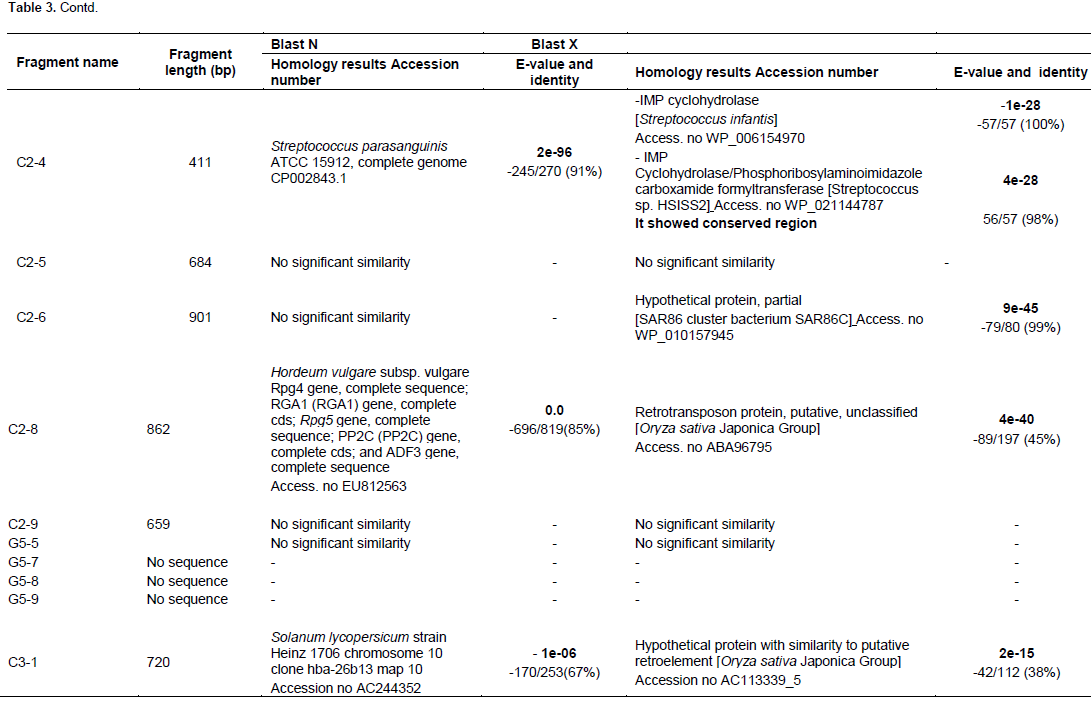
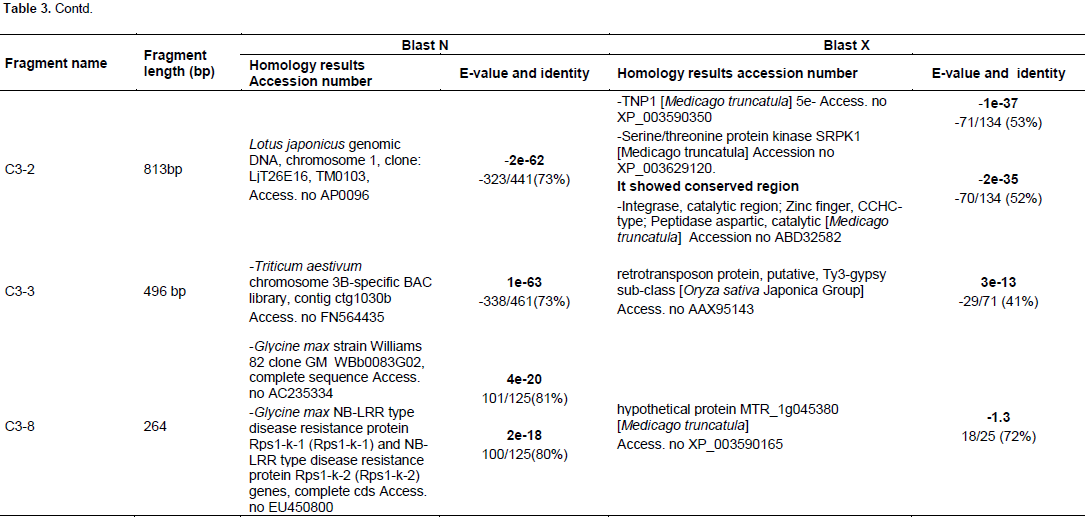
Starch metabolism
The first fragment to be discussed is C1-3 (460 bp) which
has down and up regulation pattern. The expression pattern decreased after 3 h drought treatment then increased after 10 h treatment. Blastn sequence analysis revealed significant similarities between C1-3 isolated fragment and starch branching enzyme I (sbeI) Genebank Accession No. AY304541 isolated from H. vulgare (Table 3). Also, blastx analysis of the same fragment using GeneBank databases revealed high similarities between it and the starch branching gene sebl isolated from H. vulgare Genebank Accession No. AAP72268. The results of sequence analysis using blastx and conserved blast alignments (Marchler-Baue et al., 2013) analysis tools revealed that C1-3 fragment has conserved regions with alpha amylase catalytic domain found in bacterial and eukaryotic branching enzymes (BEs) and 1,4 alpha glucan branching enzyme. This gene is responsible for introducing alpha-1,6 branch points to form amylopectin.
Starch branching enzymes (SBEs) have been found in different plant developing stages in the barley storage tissues (Sun et al., 1998). During different stages of development, the plants need starch branching activities. SBEs found in many isoforms, they can be classified into two major groups based on catalytic and structural properties, the first group is SBE family II or A and the other group, is SBE family I or B. SBEI was up-regulated under salt treatment in rice while SBEIII was down-regulated on the other hand, the contents of glucose and fructose increased (Theerawita et al., 2012). When barley plant was subjected to drought, many genes involved in carbohydrates metabolism were down regulated (Abebe et al., 2010). Normally, starch synthesis is inhibited under water deficit condition (Mahajan and Tuteja, 2005) but alpha- amylase is found to be up regulated in treated Vitis vinifera plants under cold stress (Xin et al., 2013). The current results show that the SBEI expression was affected by drought.
Glycosyl transferase gene family 1 (cell wall metabolism)
The results of blast x analysis have revealed that the two fragments, C1-6 and C1-7 have significant similarities with glycosyl transferase gene family 1 isolated from Enhyrobacter aerosaccus Genebank Accession No. WP_007116863 with E-value 12e-13 and 6e-14, respectively. The results of blastx and conserved blast alignments revealed that C1-6 and C1-7 fragments have conserved regions with glycosyl transferase GTF domain (Marchler-Baue et al., 2013). Their expression was found to be up regulated in the differential expression experiment. The glycosyl transferase enzymes (GTFs) (EC2.4) are enzymes that catalyze the glycosyl (sugar) residues transfer during biosynthesis and degradation of glycolipids, glycoproteins and polysaccharides to the acceptor which regulate the acceptors properties such as solubility, activity and transport within the cells. The glycosyltransferase have been classified into more than 90 gene families according to catalytic specificity, sequence similarities and consensus sequences. UDP glycosyltransferase gene was found to be up-regulated transcripts in cold treated V. vinifera plants (Xin et al., 2013).
Glycosyl transferase family 1 and 2 have been isolated from drought treated Gossypium herbaceum roots (Ranjan et al., 2012).
Transposons elements
The results of the differential display experiments revealed that the DD- fragment C2-2 (621 bp) showed down regulation after 3 and 10 h treatment. In blastx, the isolated fragment gave significant similarities with retrotransposon protein, putative, Ty3-gypsy subclass (Oryza sativa Japonica Group, Genebank Accession No. ABA93610.) It showed conserved region with RT_superfamily including putative active site, putative nucleic binding site, putative NTP binding site and RT_LTR retrotransposon protein (Marchler-Baue et al., 2013). On the other hand, the results of blastn analysis showed similarities between the isolated fragment and H. vulgare vrs1 locus, complete sequence and Hox1 gene. Retrotransposon elements were found to be down regulated in chickpea under drought stress (Deokr et al., 2011). The results of the differential display experiments revealed that the C3-3 fragment (496 bp) was up and down regulated under the treatment condition where, it has expression pattern up in 3 h and down in 10 h under drought treatment. The results of Blastx analysis revealed that the isolated fragment has significant similarities with retrotransposon protein, putative, Ty3-gypsy subclass (O. sativa Japonica Group) but with different Genebank Accession number AAX95143, while, it gave similarities in blastn analysis with H. vulgar BAC 631P8 Genebank Accession No. DQ249273 DD- fragment C2-8 (862 bp) that has up and down regulation pattern and gave similarities with retrotansposon protein putative, unclassified (O. sativa Japonica Group, Genebank Accession No. ABA96795 in blastx analysis while, in blastn analysis, it has a significant analysis with H. vulgare subsp. vulgare Rpg4; RGA1 gene; Rpg5 gene; PP2C gene; and ADF3 gene, Genebank Accession No. EU812563. However, DD- fragment C3-1 (720 bp) that was up-regulated in the differential expression experiment showed significant similarities in blastx with hypothetical protein with similarity to putative retroelement (O. sativa Japonica Group) but with different Genebank Accession No.AAM08859 and AC113339. Using blastn analysis, DD- fragment C3-1 showed similarities with Solanum lycopersicum strain Heinz 1706 chromosome 10 clone hba-26b13 map 10, Genebank Accession No. AC244352
Grandbastien (1998) showed that the retrotransposons activation by external change and stresses is common in eukaryotes, including plants. He suggested that several well-characterized plant retrotransposons transcriptional activation seemed to be linked tightly to active molecular pathways by stress which is under the cis regulatory sequences activation. The retrotransposons could play a role in their regulatory features modification. Different biotic and abiotic stresses activated most transposable elements in plant (Le et al., 2007) such as BARE-1 increased under drought in barley and then decreased (Kalendar et al., 2000), which showed the transposons activation played as stress adaptation. Wang et al. (2011) have isolated four retrotransponson elements from rice subjected to drought. Eissa el al. (2007 have isolated BARE-2 and partial BAGY-2 retrotransposons elements from wild barley (H. spontaneum L.) under salinity, it was down regulated.
Tomita et al. (2011) have suggested that BARE-1 element in Hordeum was the active at the recent time while the transposable elements majority of highcopy seemed to be inactive which might be interesting to perceive the adaptive evolution processes of plant. Polok and Zielinski (2011) have confirmed the transposons role in speciation in barley and many mobile transposons in the past are became stable at present.
IMP cyclohydrolase Nucleotide metabolism (synthesis of nitrogenous compounds)
DD-fragment C2-4 (411 bp) showed up-regulation in 3 h treatment and then down regulation in 10 h. This fragment has similarities with IMP cyclohydrolase (Streptococcus infantis) Genebank Accession No. WP_006154970 IMP cyclohydrolase/ Phosphoribosylaminoimidazolecarboxamide formyltransferase (Streptococcus sp. HSISS2] Genebank Accession No. WP_021144787 while, blastn analysis revealed similarities with Streptococcus parasanguinis ATCC 15912, Genebank Accession No. CP002843. According to Marchler-Baue et al. (2013) analysis method, It has conserved regions for IMPCH[cd01421], Inosine monophosphate cyclohydrolase domain, PurH[COG0138], AICAR transformylase/IMP cyclohydrolase PurH (only IMP cyclohydrolase domain in Aful), purH [TIGR00355], phosphoribosylaminoimidazolecarboxamide formyltransferase/IMP cyclohydrolase; PurH\ is bifunctional: IMP cyclohydrolase (EC 3.5.4.10) and purH[PRK00881], bifunctional phosphoribosylaminoimidazolecarboxamide formyltransferase/IMP cyclohydrolase. The enzyme aminoimidazole carboxamide ribonucleotide transformylase/inosine monophosphate cyclohydrolase (purHJ) (ATIC) encodes a bifunctional protein that catalyzes the last two steps of the de novo purine biosynthetic pathway. The biosynthetic pathway of purine has been considered as an important target for antiviral, anticancer and antimicrobial drug development because of the purine nucleotides critical role in the synthesis of RNA and DNA (Zhang et al., 2008).
Abou Ali et al. (2010) isolated 2 DD- fragments from shoots of V. villosa wild plant treated with abscisic acid which showed similarities with ATP binding/ kinase/ uracil phosphoribosyltransferaseuridine kinase (Arabidopsis thaliana) and IMP dehydrogenase/GMP reductase (Medicago truncatula), respectively. IMP dehydrogenase catalyzes the rate-limiting step in de novo biosynthesis of guanine nucleotides and has an essential role in providing necessary precursors for DNA and RNA synthesis. IMP cyclohydrolase/phosphoribosylaminoimidazolecarboxamide formyltransferase is one of the housekeeping enzymes involved in nucleotide metabolism. In the past, it was thought that the housekeeping genes are expressed in all cells at a consistent level. But, it is proved that the expression of these genes is very dynamic to meet the metabolic demands of plant cells. These changes can have an effect on plant development, so there is now a great concern about their expression. The housekeeping genes have important activities in survival of plant under stress (Moffatt and Ashihara, 2002). The identification mechanisms by which plant cells monitor and respond to their basic metabolic requirements rely on the integration of transcript, protein and metabolite profiles of different cell types (Fiehn et al., 2001). Unfortunately, the housekeeping genes expression levels complicate their analysis because small differences in activity (2 to 3 folds) may cause large metabolic impacts so they are difficult to be reproducibly detected.
Fragments with chromosomes similarities
The differentially display fragment C1-2 (613 bp), has down and up regulation expression pattern, showed similarities in blastn with Triticum aestivum gene for TaAP2-B, complete cds Genebank Accession No. AB749309. While C1-5 fragment (370 bp), has up and down regulation expression pattern, showed similarities with T. aestivum 3B chromosome, clone BAC TA3B63N2 Genebank Accession No. AM932689. Both fragments gave no significant similarities in blastx analysis.
Unknown proteins
Fragment C2-6 (901 bp) showed up regulation in 3 h treatment and then down regulation in 10 h treatment in the differential display experiment. It was cloned and sequenced. In the blastx sequence analysis, it gives significant similarities with hypothetical protein, partial sequence (SAR86 cluster bacterium SAR86C, Genebank Accession No. WP_010157945 while it does not give any significant similarities in blastn. C3-8 (264 bp) differentially displayed fragment was up regulated after 3 h of drought treatment then down regulated after 10 h. The blastx analysis showed similarities with hypothetical protein MTR_1g045380 (Medicago truncatula, Genebank Accession No. XP_003590165 while, the blastn analysis gave similarities between the C3-8 isolated fragment and Glycine max strain Williams 82 clone GM_WBb0083G02, complete sequence Genebank Accession No. AC235334 Glycine max NB-LRR type disease resistance protein Rps1-k-1 (Rps1-k-1) and NB-LRR type disease resistance protein Rps1-k-2 (Rps1-k-2) genes, complete cds Genebank Accession No. EU450800. Fragments C3-2 (813 bp) expression pattern was up regulated. It gave similarities with TNP1 (M. truncatula) Genebank Accession No. XP_003590350, hypothetical protein MTR_5g060670 (M. truncatula) Genebank Accession No. XP_003614875, Serine/threonine protein kinase SRPK1 (M. truncatula) Genebank Accession No. XP_003629120, integrase, catalytic region; Zinc finger, CCHC-type; Peptidase aspartic, catalytic (M. truncatula), Genebank Accession No. ABD32582. MTR_8g073400 included several domains, zf-CCHC; zinc knuckle, rve; Integrase core domain, gag_preintegrs; GAG-preintegrase domain, UBN2; gag-polypeptide of LTR copia-type, PKc_like; protein kinases, catalytic domain. All of these genes are shared together with UBN2 [pfam14223], gag-polypeptide of LTR copia-type domain. Three Serine/ threonine protein kinases were isolated from treated V. vinifera plants under cold stress and was found to be up regulated (Xin et al., 2013). These kinds of kinases regulate several biological processes in plant cells and were found to be localized in the cell wall and also, responded to the external environmental challenges (Morris and Walker, 2003). In Arabidopsis, SNF1-type serine/threonine protein kinase of wheat was found to enhance tolerance to multi- stress (Mao et al., 2010).
According to Marchler-Baue et al. (2013), it showed a conserved domain with UBN2 [pfam14223], gag-polypeptide of LTR copia-type. This family is found in plants and fungi, and contains LTR-polyproteins. In blast N, it gave similarities with Lotus japonicus genomic DNA, chromosome 1; clone LjT26E16, TM0103, Genebank Accession No. AP009652.
Fragments with no significant similarities
Six DD fragments, C1-1 fragment (486 bp) with down and up regulation; C1-4 fragment (504 bp) with down regulation; C2-3 (637 bp) with up-regulation; C2-5 (684 bp) with up and down regulation; C2-9 fragment (659 bp) with down and up regulation and G5-5 with up regulation, gave no significant similarities neither in blast N nor blast X.
CONCLUSION
When wild barley plants were subjected to drought, they exhibit tolerance through some genes expression. Several drought–related stress fragments have been isolated and categorized into four categories according to their expression: up; up and down: down and down and up regulation. These fragments were found to be involved in starch metabolism, cell wall metabolism, transposon element and purine metabolism. These genes especially starch metabolism genes help in the grain production. Also, there are some isolated fragments that showed similarities with drought-related stress genes; their function are unpredictable. To determine the individual regulated genes role in drought stress response, some biotechnological technique like RNAi could be used to silencing these genes and then evaluates the importance of each one of these gene using molecular and physiological techniques under drought stress. This study in the laboratory cannot give a full picture about understanding the crop response to drought and the drought stress transcriptome alone but with some field experiments combined with some physiological studies in the future will be needed in future studies to provide a clear picture in the molecular mechanisms which control the drought stress adaptation. Numbers of important genes were identified in this study, and it will be a good start for doing more biotechnological studies. This study will serve as source for discovery of new candidate genes with different gene expression and can be used in further work for manipulating drought tolerance crops. In the future, the transformation of economic crops with some of identified genes will increase stress tolerance to drought and also some other abiotic stresses.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abebe T, Melmaiee K, Berg V, Wise RP (2010). Drought response in the spikes of barley: gene expression in the lemma, palea, awn, and seed. Funct. Integ. Geno. 10(2):191-205. |
|
|
Abou Ali RMI, Shanab GML, Haider AS, Eissa HF, Salem AM (2010). Characterization and expression of abscisic acid inducible gene (s) in wild legumes. Egypt J. Genet. Cytol. 39(1):57-72. |
|
|
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucl. Acids Res. 25:3389-3402. |
|
|
Amini R (2013). Drought stress tolerance of barley (Hordeum vulgare L.) affected by priming with PEG. IJFAS J. 2(20):803-808 |
|
|
Ashraf M (2010). Inducing drought tolerance in plants: Recent advances. Biotech. Adv. 28:169-183. |
|
|
Callard D, Lescure B, Mazzolini L (1996). A method or the elimination of false positives generated by mRNA differentially display technique. Biotech. 16(6):1096-1103. |
|
|
Cattivelli L, Ceccarelli S, Romagosa I, Stanca M (2011). Abiotic stresses in barley: problems and solutions. pp 282-306. In: Ullrich SE (ed) Barley: production, improvement, and uses, Wiley-Blackwell |
|
|
Eissa HF, Saleh OM, Dyer WE (2007). Genomic characterization of stress related genes from wild barley. Egypt J. Genet. Cytol. 36(1):1-23. |
|
|
El-Shehawi AM, Elseehy MM, Hedgcoth C (2011). Isolation and sequence analysis of wheat tissue-specific cdnas by differential display. Plant Mol. Biol. Rep. 29(1):135-148. |
|
|
Farooq M, Wahid A, Kobayashi N, Fujita D, Basra SMA (2009). Plant Drought Stress: Effects, Mechanisms and Management. Agro. Sust. Dev. 29(1):185-212 |
|
|
Fastnaught CE (2001). Barley fibre. In: Cho, S., Dreher, M. (Eds.). Handbook of Dietary Fibre. Marcel Dekker, New York, pp. 519-542. |
|
|
Fiehn O, Kloska S, Altmann T (2001). Integrated studies on plant biology using multiparallel techniques. Curr. Opin. Biotech. 12:82-86. |
|
|
Gao J, Xiao Q, Yin L, He G (2008). Isolation of cDNAs clones for genes up-regulated in drought-treated Alternanthera philoxeroides root. Mol. Biol. Rep. 35:485-488. |
|
|
Grandbastien MA (1998). Activation of plant retrotransposons under stress conditions. Trends Plant Sci. 3(5):181-187. |
|
|
Iskandar HM, Casu RE, Fletcher AT, Schmidt S, Xu J, Maclean DJ, Manners JM, Bonnett GD (2011). Identification of drought-response genes and a study of their expression during sucrose accumulation and water deficit in sugarcane culms. BMC Plant Biol. 11:12. |
|
|
Kalendar R, Tanskanen J, Immonen S, Nevo E, Schulman A (2000). Genome evolution of wild barley (Hordeum spontaneum) by BARE-1 retrotransposon dynamics in response to sharp microclimatic divergence. Proc. Natl. Acad. Sci. USA, 97:6603-6607. |
|
|
Le Q, Melayah D, Bonnivard E, Petit M, Grandbastien MA (2007). Distribution dynamics of the Tnt1 retrotransposon in tobacco. Mol. Genet. Genom. 278(6):639-651. |
|
|
Liang P, Pardee AB (1992). Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Sci. 257: 967-971. |
|
|
Mahajan S, Tuteja N (2005). Cold, salinity and drought stresses: an overview. Arch. Biochem. Biophys. 444:139-158. |
|
|
Malatrasi M, Corradi M, Svensson JT, Close TJ, Gulli M, Marmiroli N (2006). A branched-chain amino acid aminotransferase gene isolated fromHordeum vulgare is differentially regulated by drought stress. Theo. Appl. Genet. 113(6):965-976. |
|
|
Mao X, Zhang H, Tian S, Chang X, Jing R (2010). TaSnRK2.4, SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. J. Exp. Bot. 61:683-696. |
|
|
Marchler-Bauer A, Lu S, Anderson JB, Chitsaz F, Derbyshire MK, DeWeese-Scott C, Fong JH, Geer LY (2013). CDD: conserved domains and protein three-dimensional structure. Nucl. Acids Res. 41(D1):D384-352. |
|
|
Moffatt BA, Ashihara H (2002). Purine and Pyrimidine Nucleotide Synthesis and Metabolism. In The Arabidopsis Book, Somerville CR, Meyerowitz EM eds (Rockville, MD: American Society of Plant Biologists). |
|
|
Morris ER, Walker JC (2003). Receptor-like protein kinases: the keys to response. Curr. Opin. Plant Biol. 6:339-342. |
|
|
Polok K, Zielinski R (2011). Mutagenic treatment induces high transposon variation in barley (Hordeum vulgare L.). Acta. Agric. Slov. 97(3):179-188. |
|
|
Ranjan A, Pandey N, Lakhwani D, Kumar DN, Pathre U, Sawant S (2012). Comparative transcriptomic analysis of roots of contrasting Gossypium herbaceum genotypes revealing adaptation to drought. BMC Genom. 13:680. |
|
|
Sambrook J, Fritsch EF, Maniatis T (1989). Molecular cloning: A laboratory manual, 2 nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. |
|
|
Sanger F, Nicklen S, Coulson AR (1977). Biochemistry DNA sequencing with chain-terminatinginhibitors (DNA polymerase/ nucleotide sequences/ bacteriophage 4X174). Proc. Nati. Acad. Sci. USA. 74:5463-5467. |
|
|
Smith A (2004). Oxford Encyclopedia of Food and Drink in America (New York: Oxford University Press). pp. 77-85. |
|
|
Stanca AM, Romagosa I, Takeda K, Lundborg T, Terzi V, Cattivelli L (2003). Diversity in abiotic stresses. pp. 179-199. In von Bothmer R, Knüpffer H, van Hintum T, Sato K (ed.) Diversity in barley (Hordeum vulgare L.), Elsevier |
|
|
Sun C, Sathish P, Ahlandsberg S, Jansson C (1998). The Two Genes Encoding Starch-Branching Enzymes IIa and IIb Are Differentially Expressed in Barley. Plant Physiol. 118:37-49. |
|
|
Theerawita C, Boriboonkaset T, Chaum S, Supaibulwatana K, Kirdmanee C (2012). Transcriptional regulations of the genes of starch metabolism and physiological changes in response to salt stress rice (Oryza sativa L.) seedlings. Physiol. Mol. Biol. Plants, 18(3):197-208. |
|
|
Tiwari JK, Munshi AD, Kumar R, Pandey RN, Arora A, Bhat JS, Sureja AK (2010). Effect of salt stress on cucumber: Na+/K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta. Physiol. Plant 32:103-114 |
|
|
Tomita M, Okutani A, Beiles A, Nevo E (2011). Genomic, RNA, and ecological divergences of the Revolvertransposon-like multi gene family in Triticea. BMC Evol. Biol. 11:269. |
|
|
Vaezi B, Bavei V, Shiran B (2010). Screening of barley genotypes for drought tolerance byagro-physiological traits in field condition. Afr. J. Agric. Res. 5(9):881-892 |
|
|
Wang WS, Pan YJ, Zhao XQ, Dwivedi D, Zhu LH, Ali J, Li ZK (2011). Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J. Exp. Bot. 62(6):1951-1960. |
|
|
Waseem M, Ali A, Tahir M, Nadeem MA, Ayub M, Tanveer A, Ahmad R, Hussain M (2011). Mechanism of drought tolerance in plant and its management through different methods. Cont. J. Agric. Sci. 5(1):10-25 |
|
|
Xin H, Zhu W, Wang L, Xiang Y, Fang L et al. (2013). Genome Wide Transcriptional Profile Analysis of Vitis amurensis and Vitis vinifera in Response to Cold Stress. PLoS ONE 8(3): e58740. |
|
|
Zhang Y, Morar M, Ealick SE (2008). Structural Biology of the Purine Biosynthetic Pathway. Cell Mol Life Sci. 65(23):3699-3724. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


