Review

ABSTRACT
The role of bioengineering in industrial processes cannot be ignored, especially in circumstances where the processes are uneconomical, demand high energy, use finite resources and emit huge amounts of carbon dioxide. Ethylene is a two-carbon unsaturated hydrocarbon that is industry's most important building block for polyester fibres, plastics, and ethylene glycol. For decades, ethylene production has relied on steam-cracking without many improvements, especially on issues of environmental impact and adoption of appropriate renewable approaches. This paper discusses selected microbial pathway modifications as novel approach to developing systems that could be alternatives to conventional ethylene production systems. Bioengineering of the ethylene pathway is suggested in view of the need to meet the criteria of high efficiency, increase sustainability and ensure product qualities and quantities that can exceed the existing approach.
Key words: Ethylene, crude glycerol, tolerance, synthesis, bioengineering.
INTRODUCTION
PLANT AND MICROBIAL METABOLIC PATHWAY STUDIES
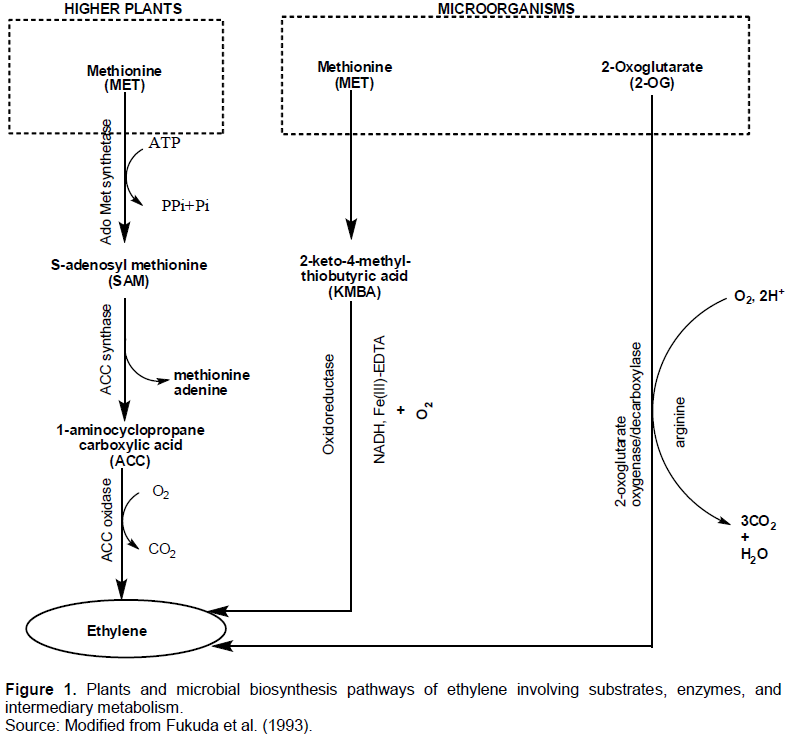
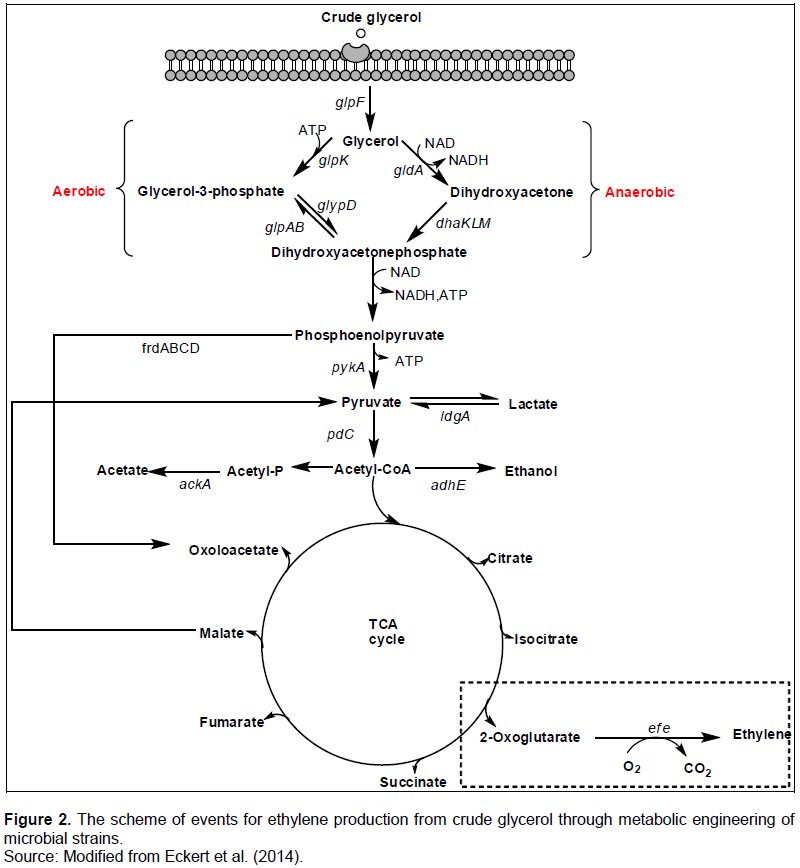
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Alshammari A, Kalevaru VN, Bagabas A, Martin A (2016). Production of ethylene and its commercial importance in the global market. Petrochemical Catalyst Materials, Processes, and Emerging Technologies. Hershey, PA: Engineering Science Reference pp. 82-115. |
|
|
American Chemical Society (2016). Beyond the ethylene steam cracker. Industrial Chemistry & Engineering. |
|
|
Amghizar I, Vandewalle LA, Van Geem KM, Marin GB (2017). New trends in olefin production. Engineering 3(2):171-178. |
|
|
Araki S, Matsuoka M, Tanaka M, Ogawa T (2000). Ethylene formation and phenotypic analysis of transgenic tobacco plants expressing a bacterial ethylene-forming enzyme. Plant & Cell Physiology 41:327-334. |
|
|
Ballester AR, González-Candelas L (2020). EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection. Journal of Fungi 6(3):175. |
|
|
Chen X, Liang Y, Hua J, Tao L, Qin W, Chen S (2010). Overexpression of bacterial ethylene-forming enzyme gene in Trichoderma reesei enhanced the production of ethylene. International Journal of Biological Sciences 6(1):96-106. |
|
|
Chiyanzu I, Cowan DA, Burton SG (2010). Immobilization of Geobaccillus pallidus RAPc8 nitrile hydratase (NHase) reduces substrate inhibition and enhances thermostability. Journal of Molecular Catalysis B: Enzymatic 63(3-4):109-115. |
|
|
Eckert C, Xu W, Xiong W, Lynch S, Ungerer J, Tao L, Gill R, Maness PC, Yu J (2014). Ethylene-forming enzyme and bio ethylene production. Biotechnology Biofuels 7(1):1-11. |
|
|
Fukuda H, Ogawa T, Tanase S (1993). Ethylene production by microorganisms. Advances in Microbial Physiology 35:275-306. |
|
|
Guerrero F, Carbonell V, Cossu M, Correddu D, Jones PR (2012) Ethylene synthesis and regulated expression of recombinant protein in Synechocystis sp PCC6803. PLOS One 7(11):e50470. |
|
|
Hamilton AJ, Bouzayen M, Grierson D (1991). Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proceedings of the National Academy of Sciences of the United States of America 88(16):7434-7437. |
|
|
Haribal VP, Chena Y, Neal L, Li F (2018). Intensification of ethylene production from naphtha via a redox oxy-cracking scheme: Process simulations and analysis. Engineering 4(5):714-721. |
|
|
Ishihara K, Matsuoka M, Inoue Y, Tanase S, Ogawa T, Fukuda H (1995). Overexpression and in vitro reconstitution of the ethylene-forming enzyme from Pseudomonas syringae. Journal of Fermentation and Bioengineering 79(3):205-211. |
|
|
Ishihara K, Matsuoka M, Ogawa T, Fukuda H (1996). Ethylene production using a broad-host-range plasmid in Pseudomonas syringae and Pseudomonas putida. Journal of Fermentation and Bioengineering 82(5):509-511. |
|
|
Nagahama K, Ogawa T, Fujii T, Tazaki M, Tanase S, Morino Y, Fukuda H (1991). Purification and properties of an ethylene-forming enzyme from Pseudomonas syringae pv. phaseolicola PK2. Journal of General Microbiology 137(10):2281-2286. |
|
|
Ogawa T, Takahashi M, Fujii T, Tazaki M, Fukuda H (1990). The role of NADH: Fe (III) EDTA oxidoreductase in ethylene formation from 2-keto-4-methylthiobutyrate. Journal of Fermentation and Bioengineering 69(6):287-291. |
|
|
Pattyn J, Vaughan-Hirsch J, Van de Poel B (2020). The regulation of ethylene biosynthesis: a complex multilevel control circuitry. New Phytologist 229(2):770-782. |
|
|
Pirkov I, Albers E, Norbeck J, Larsson C (2008). Ethylene production by metabolic engineering of the yeast Saccharomyces cerevisiae. Metabolic Engineering 10(5):276-280. |
|
|
Ren T, Patel M, Blok K (2006). Olefins from conventional and heavy feedstock: Energy use in steam cracking and alternative processes. Energy 31(4):425-451. |
|
|
Sakai M, Ogawa T, Matsuoka M, Fukuda H (1997). Photosynthetic conversion of carbon dioxide to ethylene by the recombinant cyanobacterium, Synechococcus sp. PCC 7942, which harbours a gene for the ethylene-forming enzyme of Pseudomonas syringae. Journal of Fermentation and Bioengineering 84(5):434-443. |
|
|
Tao L, Dong HJ, Chen X, Chen SF, Wang TH (2008). Expression of ethylene-forming enzyme (EFE) of Pseudomonas syringae pv. glycinea in Trichoderma viride. Applied Microbiology and Biotechnology 80(4):573-578. |
|
|
Van de Poel B, Bulens I, Hertog MLATM, Nicolaï BM, Geeraerd AH, Oppermann Y, Sauter M, De Proft MP (2011). ACC-oxidase is the rate-limiting step in ethylene biosynthesis during postharvest storage of tomato. Acta Horticulturae, 945:429-434. |
|
|
Zhang M, Yu Y (2013) Dehydration of ethanol to ethylene. Industrial & Engineering Chemistry Research 52(28):9505-9514. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


