ABSTRACT
Bacterial wilt caused by Ralstonia solanacearum is a major constraint to production of potatoes (Solanum tuberosum L). To date, there are no known conventional bactericides which provide effective control of this soil borne pathogen. This study aimed at investigating the antibacterial activity of eugenol and thymol which are the major components of thyme and clove essential oils, respectively against R. solanacearum. The pathogen was isolated from potato tubers exhibiting bacterial wilt symptoms. The inhibitory effect of these compounds was assayed by disc diffusion and minimal inhibition concentration (MIC) methods. Combinational test was also performed using chequerboard assay. At a concentration of 100 mg mL-1, thymol and eugenol had inhibition zones of 27 and 17 mm, respectively. The MIC of eugenol and thymol were determined as 275 and 175 µg mL-1, respectively and both had bactericidal effect against R. solanacearum. Combinational test revealed an additive effect indicating that their combined use does not significantly enhance inhibition. Both compounds have a potential to be exploited as antibiotic for the management of bacterial wilt disease albeit thymol at a lower concentration. Exploitation of these volatile compounds in vivo however faces challenges which can be overcome through nanoencapsulation by an appropriate nanocarrier.
Key words: Essential oils, inhibitory effect, combinational test, chequerboard assay, bactericidal effect.
Potato (Solanum tuberosum L.) is one of the most important food crops which provides a greater potential for food security and incomes around the world (Gildemacher et al., 2009). The productivity of the crop is however limited by both biotic and abiotic stresses. Major biotic stresses affecting potato yield are diseases which include bacterial wilt, late blight and early blight (Muthoni and Nyamongo, 2009). Out of these, bacterial wilt has no known bactericide commercially released for use which can efficiently manage to control the disease and it leads to high reduction in crop yields. This is caused by Ralstonia solanacearum, a Gram-negative bacterium which not only causes bacterial wilt disease in potato but also on more than 200 other plant species that include tomato, eggplant, tobacco, geranium, pepper and banana (Aguk et al., 2018; Hayward, 1991; Meng, 2013; Muthoni et al., 2014). R. solanacearum is mainly spread through infected seed stocks but can also spread from infested to healthy fields by farm equipment, irrigation water and plant-to-plant through the rhizosphere (Patil et al., 2012). Damaged roots arising from wounds caused by soil borne organisms like the nematodes or wounds formed by the emergence of lateral roots can also spread R. solanacearum by acting as an avenue for infection (Karim and Hossain, 2018). Once it gets into the roots, it colonizes the xylem in the vascular bundles and blocks water uptake resulting in wilting and eventual death (Meng, 2013). The bacteria can survive in water and in residues of infected plants for a long period of time making its control very difficult (Pradhanang et al., 2000).
Despite concerted efforts to control the disease, bacterial wilt continues to cause serious economic challenges in many countries in the world (Aguk et al., 2018; Karim and Hossain, 2018). Worldwide, the pathogen infestation in potatoes has been reported to lead to yield losses of between 33 and 90% and up to $1 billion yearly in lost revenue (Elphinstone, 2005). In Kenya, the disease has been reported to affect over 70% of all potato farms leading to yield losses of between 50 and 100% (Muthoni et al., 2014). Despite using a wide range of management practices such as use of chemical pesticides, phytosanitary measures, breeding for resistance and bio antagonists, dealing with the pathogen still remains a challenge (Karim and Hossain, 2018; Meng, 2013). These limitations may be due to the complexity of R. solanacearum pathogenicity which enhances its resistance to the available control methods. Essential oils however have different antibacterial mechanisms including inhibition of swarming motility, biofilm formation, cell membrane damage and cell death making it a powerful tool in reducing bacterial resistance (Chen et al., 2019). So far, there is no report on a known conventional bactericide which has shown effective control of this soil borne pathogen. Therefore, there is need to explore other biologically sustainable control methods which can form part of the integrated management system of the pathogen. Previously, R. solanacearum species complex was known to include a number of phenotypically diverse strains which were grouped into five biovars and five pathogenic races. It was later reclassified into Phylotypes I, II, III and IV based on sequence analysis. In the recent taxonomic revision, the R. solanacearum species complex has been reclassified into three distinct species which include R. solanacearum (Phylotype II) which causes potato brown rot (Bacterial wilt), Ralstonia pseudosolanacearum (Phylotype I and III) and Ralstonia syzygii (Phylotype IV) (Paudel et al., 2020).
Plants produce more than 100,000 known secondary metabolites some of which function in protecting plants against predators while others act against pathogens such as viruses, bacteria and fungi (Bassolé and Juliani, 2012). Eos are secondary metabolites whose antimicrobial activity against R. solanacearum have been reported in several studies (Ji et al., 2005; Oboo et al., 2014). Previous studies have revealed that EOs containing major components such as citral, carvacrol, cinnamaldehyde, thymol or eugenol have the highest activity against pathogenic microbes (Bassolé and Juliani, 2012). However, the inhibitory activity of some of these compounds including eugenol against R. solanacearum has not been investigated.
Among the components of clove EO, eugenol accounts for up to 95% of the oil (Nurdjannah and Bermawie, 2012). The antibacterial effect of clove EO has been attributed to the presence of its major component, eugenol (Chaieb et al., 2007). Eugenol is mainly concentrated in the buds and flowers of clove (Zingiber aromaticum). A part from clove, eugenol is also found in herbal plants such as thyme, ginger and turmeric (Charan et al., 2015; Khalil et al., 2017). Chemically, the IUPAC name of eugenol is 4-Allyl-2-methoxyphenol and it belongs to a class of phenylpropanoids (C10H12O2) and hence can be produced commercially. It is also known as caryophyllic acid, allylguaiacol or 4-allylcatechol-2-methyl ether (Charan et al., 2015). Thymol is a phenol compound and a major volatile component of EO extracted from medicinal plants thyme, basil and oregano. It is a colorless crystalline compound whose IUPAC name is 2-isopropyl-5-methylphenol and can also be commercially synthesized. Thyme EO has been shown to inhibit the growth of Aspergillus niger and Aspergillus tubingensis both of which are fungal infections in grapes (Císarová et al., 2016). Thymol also has inhibitory effect on pathogenic plant bacterial species Erwinia carotovora and Xanthomonas compestris (Nesrollahi and Razavi, 2017).
When major components of EOs interact, they may have additive, antagonistic or synergistic antibacterial effects (Mourey and Canillac, 2002). Bassolé and Juliani (2012)reported that the efficacy of several combinations of EOs and their components were shown to increase when combined which took advantage of their additive and synergistic effects. Pei et al. (2009)reported a synergistic effect against Escherichia coli when eugenol and thymol was combined in the ration of 1:4, respectively. This significantly lowered the effective concentration of the components by 50% than when each component was used alone. The effect of combining eugenol and thymol against R. solanacearum has however not been studied. The present study therefore aimed at investigating the antibacterial activity of individual and mixed fractions of eugenol and thymol against R. solanacearum.
Eugenol ≥98% (E51791) and thymol ≥99% (16254) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Other chemicals, such as triphenyl tetrazolium chloride (TZC) peptone, glucose, casamino-acid and agar were purchased from Legacy Lab. Africa.
Isolation and detection of R. solanacearum
R. solanacearum was isolated in October 2020 from potato samples exhibiting bacterial wilt symptoms collected from field grown crops in Nyandarua County in Kenya. The procedure for isolation and purification was according to Mutimawurugo et al., (2019). The isolates were grown on triphenyl tetrazolium chloride (TZC) media (peptone 10 g, glucose 2.5 g, casamino-acid 1 g, agar 18 g, TZC 50 mg in 1 L of distilled water). The TZC medium was used to differentiate cultures of R. solanacearum into virulent and avirulent colony types as described by Champoiseau et al., (2009). Polymerase chain reaction (PCR) was used to confirm the identity of the virulent isolates using primer pair (Nmult:21:2F/Nmult:22:RR) which is specific to R. solanacearum species (Fegan and Prior, 2005).
Evaluation of the antibacterial activity of eugenol and thymol
The antibacterial activity of eugenol, thymol and their combinations were evaluated using disc diffusion and broth microdilution methods as described subsequently. Dimethylsulfoxide (DMSO) alone was used as vehicle control in all the assays (Valliammai et al., 2020).
Evaluation of the antibacterial activity by disc diffusion
Antibacterial activity of thymol and eugenol against R. solanacearum was confirmed by disc diffusion test also known as the Kirby-Bauer assay method (Bauer et al., 1966)as described by Ambrosio et al. (2019). The test was performed in sterile 90 mm diameter Petri dishes containing solid and sterile casamino acids-peptone-glucose (CPG) nutrient agar medium. Before being tested, the solubility of thymol and eugenol were enhanced by dissolving them in DMSO. Thymol and eugenol were tested at six levels: 1000, 800, 600, 400, 200 and 100 mg mL-1 concentrations in DMSO. Sterile 6 mm diameter filter paper discs (Whatman paper No. 5) were impregnated with 10 μl/disk of eugenol or thymol at the different concentrations. The discs were then placed on the surface of the medium which had been flood inoculated with 100 μL of overnight microbial suspension (108 CFU mL-1) using a sterile swab. The overnight culture of the bacterium was prepared by diluting to 108 CFU mL-1 using a spectrophotometer (Jenway 6305, UK) at a wavelength of 625 nm. A negative control included was a filter disc soaked in 10 μl of DMSO, while the positive control consisted of gentamycin antibiotic discs (10 μg/disk). After 24-h incubation at 28°C without lighting, the antibacterial activity of the antimicrobials was evaluated by measuring the diameter of the inhibition zone in mm. Each experiment had three replicates and the mean diameter of the inhibition zone was recorded.
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC)
MIC was determined by micro broth dilution assay using resazurin test (Elshikh et al., 2016). Stock solutions of eugenol and thymol were prepared separately by dissolving each in DMSO to make a stock concentration of 800 μg mL-1 then serial two-fold dilution were prepared to reach final concentrations ranging from 400 to 200 μg mL-1 (400, 375, 350, 325, 300, 275, 250, 225 and 200) for eugenol and 200 to 100 μg mL-1 (200, 187.5, 175, 162.5, 150, 137.5, 125, 112.5 and 100) for thymol, respectively. The ranges were determined from a pre-experiment using a standard two-fold micro broth dilution. A volume of 100 μL of sterilized CPG medium was distributed from the 1st to the 9th well in each row. The same volume (100 μL) of each stock concentration was transferred to the first well microtitre plate in triplicate. Then, 100 μL of the thymol/eugenol-broth mixture was transferred from the 1st well to the 2nd well in the next row and so on till the 8th well and the last 100 μL of the antibiotic broth mixture discarded. A suspension (10 μL) of R. solanacearum at the concentration of 108 CFU mL-1 was then added to each of the wells. MIC was then determined after incubation for 24-h at 28°C using resazurin test. This was performed by adding 20 µL of resazurin solution at 0.015% m/v per well and the results recorded after further incubation at 28°C for 1 h. Each experiment had two treatments (thymol and eugenol) with a positive and negative control. For the positive control, 10 μL of R. solanacearum was added to 100 μL of nutrient broth lacking the test compound (essential oil) to check the growth of incubated bacteria. A well containing only 100 μL of broth was prepared as a negative control to check for sterility of the medium (Hasani and Hasani, 2018). Each experiment was carried out in triplicate and in three independent repeats and the results expressed as mean ± standard deviation. For wells showing inhibitions, 20 mL of the mixture were transferred to TZC agar plates. Then MBC was determined as the lowest concentration where no growth was observed on the TZC plates after 24-h incubation at 28°C (Xue and Knoxville, 2015). MBC/MIC ratio of 1-2 and 4-16 was considered bactericidal and bacteriostatic, respectively (Oulkheir et al., 2017).
Effect of combined eugenol and thymol on the inhibition of R. solanacearum
The interaction between thymol and eugenol on their antibacterial effectiveness against R. solanacearum was performed based on the fractional inhibitory concentration (FIC) index obtained through checkerboard assay using 96-well microtitre plates (Sarah and Lobna, 2017). The test concentration range for the synergy test was between 0 and 400 μg mL-1. The final volume in each well was 100 µL comprising 50 µL of each diluted compound. Subsequently, 10 µL of bacterial suspension containing 1 × 108 CFU/mL of R. solanacearum was added to each well. The microtitre plate was incubated at 28°C for 24-h and MIC was read as the lowest concentration of the agent at which no visible growth occurred. The MIC of the combinations was determined using resazurin test. Positive control was prepared by adding 10 μL of R. solanacearum in a well containing 100 μL of nutrient broth while negative control had only 100 μL of broth (Hasani and Hasani, 2018). For comparison of MIC and FIC values, tests were made in triplicate and repeated three times. FIC index was then calculated for the thymol/eugenol combination using the following standard equations (Odds, 2003):
FIC index = FICA + FICB
where FICA and FICB (where A and B are thymol and eugenol respectively) are the minimum concentrations that inhibited the bacterial growth for antibacterial compounds A and B, respectively. Results were provided in tabular form. The results of the FIC index were interpreted as Synergy (FIC index value of ≤0.5), Additive (FIC index value of 0.5 < FIC ≤ 4), or Antagonism (FIC index value of FIC > 4) (Odds, 2003).
Isolation and identification of R. solanacearum
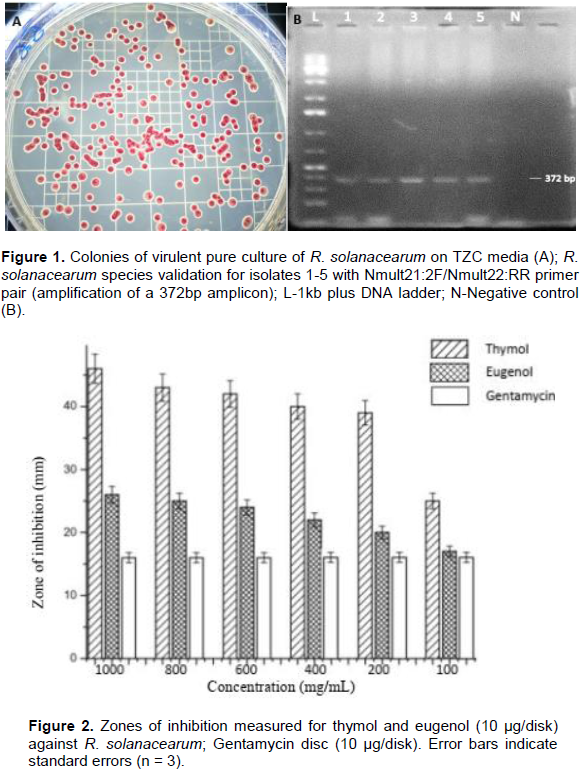
Morphological and molecular tests were used to ascertain the identities of the suspected R. solanacearum isolates. After 48-h incubation, TZC agar differentiation test was used to distinguish virulent from avirulent isolates. The virulent isolates were identified based on their characteristic red center and whitish margin (Figure 1). The identity of the virulent isolates was further confirmed by PCR using primers specific to R. solanacearum species (Figure 1) (Fegan and Prior, 2005).
Evaluation of the antibacterial activity by disc diffusion
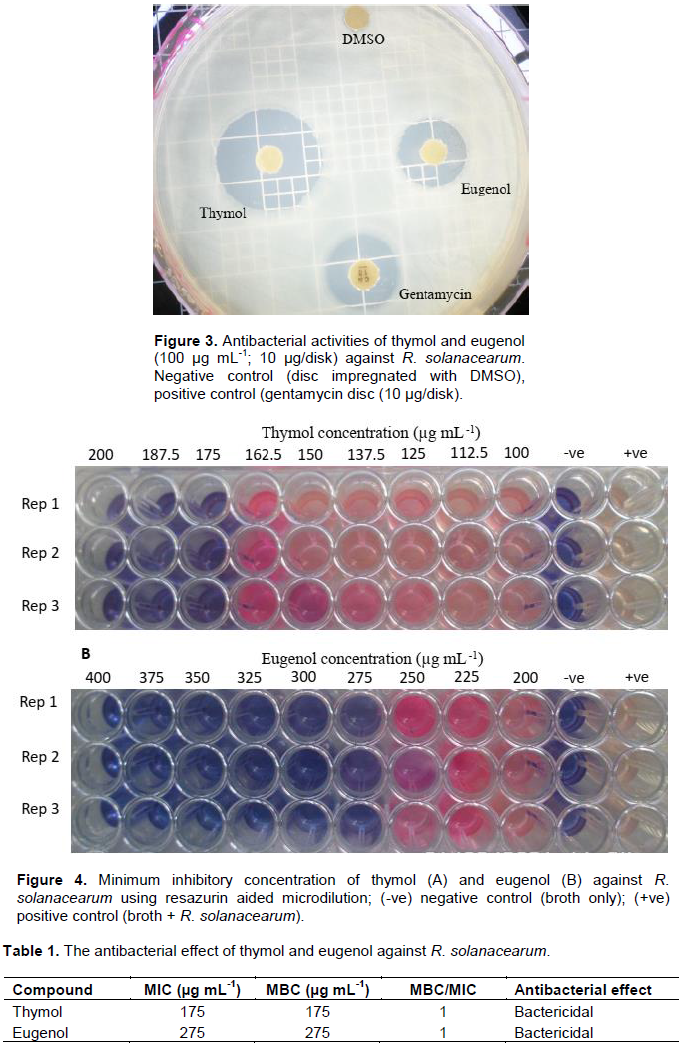
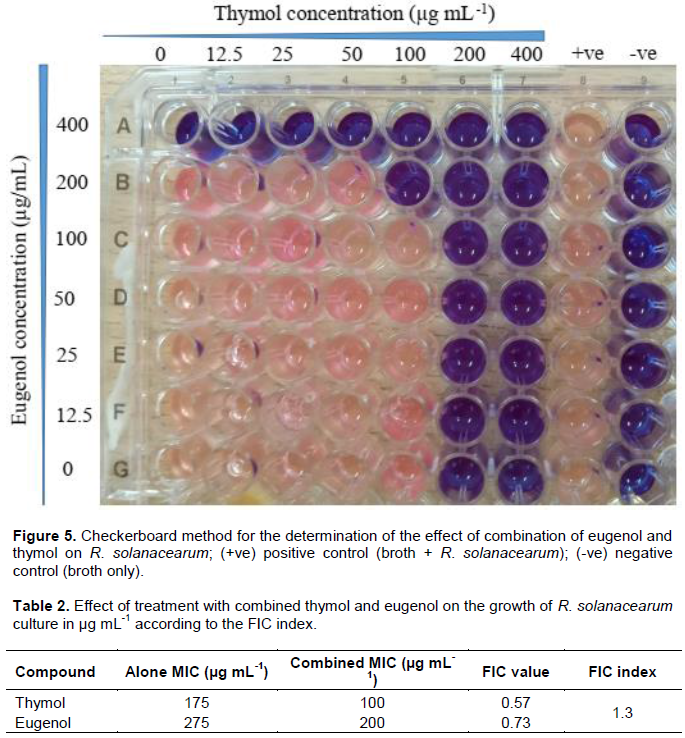
The antimicrobial activity of thymol against R. solanacearum was higher than that of eugenol for all the ranges of concentrations (Figure 2). At the concentration of 100 mg mL-1, thymol exhibited a zone of inhibition of 26 ± 0.6 mm against R. solanacearum in comparison to eugenol and gentamycin which had inhibition zones of 17 ± 0.4 and 20 ± 0.5 mm, respectively (Figure 3).
Minimum inhibitory concentration and minimum bactericidal concentration
The MIC and MBC was used to evaluate the antibacterial activity of eugenol and thymol in relation to virulent R. solanacearum isolate. After 24-h incubation, the MIC was determined as the lowest concentration where inhibition occurred (blue resazurin colour remained unchanged) (Figure 4). For the MBC, 20 µl of the contents of the wells where inhibition occurred were plated on CPG medium and incubated at 28°C for 24-h.The MIC value was determined as 175 and 275 mg mL-1 for thymol and eugenol, respectively. For the MBC, there was no growth in all the wells where inhibition occurred. The lowest concentration for each of the compounds was therefore taken as the MBC. Thymol and eugenol were found to have bactericidal effect on R. solanacearum (Table 1).
Effect of combined eugenol and thymol on the inhibition of R. solanacearum
Checkerboard assay was performed to study the antibacterial interaction between thymol and eugenol against R. solanacearum. The lowest concentration which inhibited the growth of bacteria was found when 100 mg mL-1 of thymol combined with 200 mg mL-1 of eugenol (Figure 5). The FIC value for the most effective combination was determined as 0.57 and 0.73 for thymol and eugenol which resulted to a FIC index of 1.3 (Table 2).
The potato bacterial wilt pathogen, R. solanacearum was successfully isolated and identified by both morphological and molecular means. The isolation was done using TZC medium which is useful in distinguishing R. solanacearum among other bacteria during isolation, and for distinguishing virulent colonies from avirulent mutant ones during purification of cultures (Kelman, 1954). The virulent colonies were identified based on their characteristic fluidal and white with pink centers as reported by Aley and Elphinstone (1995). The morphologically determined virulent isolates were further confirmed to belong to R. solanacearum species (previously classified as phylotype II) using species specific primers (Nmult21:2F/Nmult22:RR) which amplified a 372 bp in the intergenic spacer region between the 16S and 23S rRNA genes (Fegan and Prior, 2005).
The methods used in assessing the antimicrobial activity of EOs include agar dilution, disc diffusion and broth dilution (Perricone et al., 2015). Using the disc diffusion assay, thymol was shown to have the highest inhibition zones in all the tested concentrations in comparison to both eugenol and gentamycin which was used as a positive control (Figure 2). There was no growth inhibition in the disc impregnated with DMSO, an indication that it has no activity against R. solanacearum.
Similar observation was reported previously by Li et al. (2016). In the previous study using agar diffusion, a strong antibacterial activity of thyme oil was reported against 120 clinical strains of bacteria of the genera Escherichia, Enterococcus, Pseudomonas and Staphylococcus (Sienkiewicz et al., 2012). Using disc diffusion test to study the antibacterial and antifungal activity of eugenol, Pavesi et al. (2018)reported inhibition zones of 12.1, 4.2 and 3.2 mm for Candida albicans, E. coli and Staphylococcus aureus, respectively. This variation in results may be due to the disc size, amount of compound impregnated into the disc and the volume of agar used. Disc diffusion test provides only qualitative results and neither quantification of the antimicrobial response nor distinguishing bacteriostatic from bactericidal effect of the antimicrobial is possible with this method (Nasir et al., 2015). In addition to the disc diffusion test, it has been recommended that the MIC or cidal concentration values of the test compound be established (Orchard and Van Vuuren, 2017).
The antimicrobial activities of eugenol and thymol were evaluated for their MIC and MBC using micro broth dilution assay (Figure 5). The MIC of thymol and eugenol was determined as 175 mg mL-1 and 275mg mL-1 respectively using the resazurin indicator dye. Resazurin dye is a redox indicator and has been used to determine the MIC of a number of antibacterial compounds (Elshikh et al., 2016; Kokina et al., 2019). In the presence of active bacterial cells, blue non-fluorescent resazurin is reduced to pink fluorescent resorufin which can further be reduced to uncolored nonflourescent hydroresorufin and hence can give a direct measure of the metabolic activity of the bacteria (O’Brien et al., 2000). The MIC of thymol was significantly lower than that of eugenol indicating that it has a higher antibacterial activity against R. solanacearum. The high antibacterial activity of thymol in comparison to eugenol has also been reported against E. coli (Pei et al., 2009). This results were in the same range with a previous study by Sarah and Lobna (2017)which reported the MIC values (128 to 256 μg mL-1) of thymol against different isolates of S. aureus (Sarah and Lobna, 2017). However in a different study to evaluate the antibacterial effect of eugenol and thymol against E. coli using two-fold micro broth dilution, the MIC was determined as 1600 and 400 mg mL-1, respectively (Pei et al., 2009). This difference in results with the current study can be attributed to the different microbes tested and the different experimental conditions used. The MIC of eugenol against a number of other pathogens have also been reported including S. aureus, Salmonella enterica and fungi including Cladosporium (MIC: 100 µg mL-1), Aspergillus (MIC:100 µg mL-1) and Cladosporium species (MIC: 350 µg mL-1) (Kit-Kay et al., 2019). This is however the first study in which the MIC of eugenol has been reported against R. solanacearum. A study by Jeyakumar and Lawrence (2021)on the mechanism of action of eugenol against E. coli demonstrated that it alters membrane permeability of the bacteria which then leads to leakage of intracellular contents and hence death of the bacteria. Thymol on the other hand inhibits the bacterial growth by increasing the permeability of its cytoplasmic membrane to adenosine triphosphate leading to the release of lipopolysaccharides (Pei et al., 2009).
The lowest concentration of the sample that resulted to ≥99.9% kill of the initial inoculum was considered as MBC as described by Oulkheir et al. (2017)and was determined as 175 and 275 mg mL-1 for thymol and eugenol, respectively. This was similar to their respective MIC’s which indicated that both thymol and eugenol has bactericidal effect against R. solanacearum and hence both can be considered as potential candidates for use in controlling R. solanacearum. Determination of both MIC and MBC of these compounds is important in the determination of the effective concentration in vitro, which in turn will help in recommending the dosage and to assess the potential for using these compounds for crop protection in agriculture (Cai et al., 2018).
The antibacterial efficacy can be enhanced if two EO components get mixed together or with other known antimicrobials (Kalemba and Synowiec, 2019). When EO compounds interact, they may lead to additive, antagonistic, or synergistic effects (Odds, 2003). The antimicrobial properties of a number EO components and their combinations have been reported in several studies (Bassolé and Juliani, 2012). For example, eugenol was reported to exhibit a synergistic antibacterial activity against E. coli when in combination with thymol (Charan et al., 2015). The checkerboard assay in this study revealed an additive effect when thymol was combined with eugenol against R. solanacearum (FIC =1.3) (Table 2). This meant that the sum of the individual effect of eugenol and thymol is equal to their combined effect. A similar combination against E. coli was however synergistic (Pei et al., 2009). Lambert et al. (2001)reported an additive effect when carvacrol and thymol were combined against Pseudomonas aeruginosa and S. aureus and they showed that the overall inhibition could be attributed mainly to the additive antimicrobial action of those two compounds. This variation in the effect is because the effect of combining antibacterial compounds is dependent on the combination of the components tested and the microbe to which it is tested against (Bassolé et al., 2010). The effective concentration of thymol when in combination with eugenol was however reduced by 43% against R. solanacearum while that of eugenol was reduced by 27%. This additive effect of thymol/eugenol may be due to the fact that thymol disintegrated the outer membrane of R. solanacearum, making it easier for eugenol to enter the cytoplasm and combine with proteins (Pei et al., 2009).
The utilization of EO compounds in controlling soil borne phytopathogens in the field is limited by their instability when exposed to environmental and physiological factors which may lead to the loss of their potency (Kit-Kay et al., 2019). In order therefore to utilize eugenol and thymol in controlling R. solanacearum in potato farms, there is need for a proper delivery system. Studies have shown that encapsulating EOs in biodegradable polymer shells has a potential of protecting them from volatilization and sustenance of their antioxidant and antimicrobial properties (Jamil et al., 2016).
This study has shown that both eugenol and thymol have antibacterial properties against R. solanacearum. The MIC of thymol (175 μg mL-1) was however found to be lower than that of eugenol (275 μg mL-1). Combinational test using checkerboard assay revealed an additive effect when the two compounds were mixed against R. solanacearum indicating that their combined use does not enhance inhibition. The use of these essential oil compounds in controlling R. solanacearum in vivo may however be limited due to their volatility, hydrophobicity, rapid degradation as well as poor solubility in water. These challenges can be overcome through nano-encapsulating them into lipid or polymer based organic nanocarriers.
The authors have not declared any conflict of interests.
The authors acknowledge the financial support from the African Union through the Pan African University, Institute for Basic Sciences and Technology.
REFERENCES
|
Aguk JA, Karanja N, Schulte-Geldermann E, Bruns C, Kinyua Z, Parker M (2018). Control of bacterial wilt (Ralstonia solanacearum) in potato (Solanum tuberosum) using rhizobacteria and arbuscular mycorrhiza fungi. African Journal of Food, Agriculture, Nutrition and Development 18(2):13371-13387.
Crossref
|
|
|
|
Aley E, Elphinstone J (1995). Culture media for Ralstonia solanacearum isolation, identification and maintenance. Fitopatologia 30:126-130.
|
|
|
|
|
Ambrosio CM, Ikeda NY, Miano AC, Saldaña E, Moreno AM, Stashenko E, Contreras-Castillo CJ, Da Gloria EM (2019). Unraveling the selective antibacterial activity and chemical composition of citrus essential oils. Scientific Reports 9(1):1-13.
Crossref
|
|
|
|
|
Bassolé IHN, Juliani HR (2012). Essential oils in combination and their antimicrobial properties. Molecules 17(4):3989-4006.
Crossref
|
|
|
|
|
Bassolé IHN, Lamien-Meda A, Bayala B, Tirogo S, Franz C, Novak J, Nebié RC, Dicko MH (2010). Composition and Antimicrobial Activities of Lippia multiflora Moldenke, Mentha x piperita L. and Ocimum basilicum L. Essential Oils and Their Major Monoterpene Alcohols Alone and in Combination. Molecules 15(11):7825-7839.
Crossref
|
|
|
|
|
Bauer AW, Kirby WMM, Sherris JC, Turck M (1966). Antibiotic Susceptibility Testing by a Standardized Single Disk Method. American Journal of Clinical Pathology 45(4):493-496.
Crossref
|
|
|
|
|
Cai L, Chen J, Liu Z, Wang H, Yang H, Ding W (2018). Magnesium oxide nanoparticles: Effective agricultural antibacterial agent against Ralstonia solanacearum. Frontiers in Microbiology 9(4):1-19.
Crossref
|
|
|
|
|
Chaieb K, Hajlaoui H, Zmantar T, Kahla-Nakbi AB, Rouabhia M, Mahdouani K, Bakhrouf A (2007). The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): a short review. Phytotherapy Research 21(6):501-506.
Crossref
|
|
|
|
|
Champoiseau PG, Jones JB, Allen C (2009). Ralstonia solanacearum Race 3 Biovar 2 Causes Tropical Losses and Temperate Anxieties. Plant Health Progress 10(1):35.
Crossref
|
|
|
|
|
Charan RMR, Srinivasan V, Selvaraj S, Mahapatra SK (2015). Versatile and Synergistic Potential of Eugenol: A Review. Pharmaceutica Analitica Acta 6(6):1-6.
|
|
|
|
|
Chen J, Mao S, Xu Z, Ding W (2019). Various antibacterial mechanisms of biosynthesized copper oxide nanoparticles against soilborne Ralstonia solanacearum. RSC Advances 9(7):3788-3799.
Crossref
|
|
|
|
|
Císarová M, Tan?inová D, Medo J (2016). Antifungal activity of lemon, eucalyptus, thyme, oregano, sage and lavender essential oils against Aspergillus niger and Aspergillus tubingensis isolated from grapes. Potravinarstvo 10(1):83-88.
Crossref
|
|
|
|
|
Elphinstone J (2005). Edited by Allen C, Prior P, Hayward AC. St. Paul, MN: APS Press. In The current bacterial wilt situation: a global overview. In: Bacterial wilt disease and the Ralstonia solanacearum species complex. Edited by Allen C, Prior P, Hayward AC. St. Paul, MN: APS Press.
|
|
|
|
|
Elshikh M, Ahmed S, Funston S, Dunlop P, McGaw M, Marchant R, Banat IM (2016). Resazurin-based 96-well plate microdilution method for the determination of minimum inhibitory concentration of biosurfactants. Biotechnology Letters 38(6):1015-1019.
Crossref
|
|
|
|
|
Fegan M, Prior P (2005). How complex is the "Ralstonia solanacearum species complex". Bacterial wilt disease and the Ralstonia solanacearum species complex 1:449-461.
|
|
|
|
|
Gildemacher PR, Kaguongo W, Ortiz O, Tesfaye A, Woldegiorgis G, Wagoire WW, Kakuhenzire R, Kinyae PM, Nyongesa M, Struik PC, Leeuwis C (2009). Improving potato production in Kenya, Uganda and Ethiopia: A system diagnosis. Potato Research 52(2):173-205.
Crossref
|
|
|
|
|
Hasani M, Hasani S (2018). Nano-encapsulation of thyme essential oil in chitosan-Arabic gum system: Evaluation of its antioxidant and antimicrobial properties. Journal of Food Biochemistry 2(2):75-82.
Crossref
|
|
|
|
|
Hayward AC (1991). Biology and Epidemiology of Bacterial Wilt Caused by Pseudomonas solanacearum. Annual Reviews Phytopathology 29:65-87.
Crossref
|
|
|
|
|
Jamil B, Abbasi R, Abbasi S, Imran M, Khan SU, Ihsan A, Bokhari H (2016). Encapsulation of cardamom essential oil in chitosan nano-composites: In-vitro efficacy on antibiotic-resistant bacterial pathogens and cytotoxicity studies. Frontiers in Microbiology 7(1580):1-10.
Crossref
|
|
|
|
|
Jeyakumar GE, Lawrence R (2021). Mechanisms of bactericidal action of Eugenol against Escherichia coli. Journal of Herbal Medicine 26:100406.
Crossref
|
|
|
|
|
Ji P, Momol MT, Olson SM, Pradhanang PM, Jones JB (2005). Evaluation of thymol as biofumigant for control of bacterial wilt of tomato under field conditions. Plant Disease 89(5):497-500.
Crossref
|
|
|
|
|
Kalemba D, Synowiec A (2019). Agrobiological Interactions of Essential Oils of Two Menthol Mints: Mentha piperita and Mentha arvensis. Molecules 25(1):1-33.
Crossref
|
|
|
|
|
Karim Z, Hossain M (2018). Management of Bacterial Wilt (Ralstonia solanacearum) of Potato: Focus on Natural Bioactive Compounds. Journal of Biodiversity, Conservation and Bioresouce Management 4(1):1-10.
Crossref
|
|
|
|
|
Kelman A (1954). The relationship of pathogenecity in Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 44(12):693-695.
|
|
|
|
|
Khalil AA, Rahman UU, Khan MR, Sahar A, Mehmood T, Khan M (2017). Essential oil eugenol: Sources, extraction techniques and nutraceutical perspectives. RSC Advances 7(52):32669-32681.
Crossref
|
|
|
|
|
Kit-Kay M, Masnah BK, Sunday BA, Raghavendra S, Yew-Beng K, Kavitha M, Madhu KB, Sazali HA, Mallikarjuna RP (2019). A Comprehensive Review on Eugenol's Antimicrobial Properties and Industry Applications: A Transformation from Ethnomedicine to Industry. Pharmacognosy Reviews 13(25):1-9.
|
|
|
|
|
Kokina M, Salevic A, Kaluševic A, Levic S, Pantic M, Dejan P, Šavikin K, Shamtsyan M, Nikšic M, Nedovic V (2019). Characterization, antioxidant and antibacterial activity of essential oils and their encapsulation into biodegradable material followed by freeze drying. Food Technology and Biotechnology 57(2):282-289.
Crossref
|
|
|
|
|
Lambert RJW, Skandamis PN, Coote PJ, Nychas GJE (2001). A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. Journal of Applied Microbiology 91(3):453-462.
Crossref
|
|
|
|
|
Li S, Yu Y, Chen J, Guo B, Yang L, Ding W (2016). Evaluation of the antibacterial effects and mechanism of action of protocatechualdehyde against Ralstonia solanacearum. Molecules 21(6):754.
Crossref
|
|
|
|
|
Meng F (2013). Ralstonia Solanacearum Species Complex and Bacterial Wilt Disease. Journal of Bacteriology and Parasitology 04(02):2-5.
Crossref
|
|
|
|
|
Mourey A, Canillac N (2002). Anti-Listeria monocytogenes activity of essential oils components of conifers. Food Control 13(4-5):289-292.
Crossref
|
|
|
|
|
Muthoni J, Kabira J, Shimelis H, Melis R (2014). Spread of Bacterial Wilt Disease of Potatoes in Kenya: Who is to Blame? International Journal of Horticulture 4(3):10-15.
Crossref
|
|
|
|
|
Muthoni J, Nyamongo DO (2009). A review of constraints to ware Irish potatoes production in Kenya. A Journal of Horticulture and Forestry 1(7):98-102.
Crossref
|
|
|
|
|
Mutimawurugo MC, Wagara IN, Muhinyuza JB, Ogweno JO (2019). Virulence and characterization of isolates of potato bacterial wilt caused by Ralstonia solanacearum (Smith) in Rwanda. African Journal of Agricultural Research 14(6):311-320.
|
|
|
|
|
Nasir B, Fatima H, Ahmad M, Ihsan-ul-Haq (2015). Recent Trends and Methods in Antimicrobial Drug Discovery from Plant Sources. Austin Journal of Microbiology 1(1):1-12.
|
|
|
|
|
Nesrollahi P, Razavi SM (2017). Herbicidal and Anti Pathogenic Potential of Thymol. Scholars Academic Journal of Biosciences 5(3):183-186.
|
|
|
|
|
Nurdjannah N, Bermawie N (2012). 11-Cloves. In Handbook of Herbs and Spies. (Second E. Peter (Ed.), Woodhead Publishing Series in Food Science, Technology and Nutrition 1:197-215 Woodhead Publishing.
Crossref
|
|
|
|
|
O'Brien J, Wilson I, Orton T, Pognan F (2000). Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. European Journal of Biochemistry 267(17):5421-5426.
Crossref
|
|
|
|
|
Oboo H, Muia A, Kinyua Z (2014). Effect of Essential Oil Plant Extracts on in vitro Growth of Ralstonia solanacearum. Egerton Journal of Science and Technology 14(2073):141-160.
|
|
|
|
|
Odds FC (2003). Synergy, antagonism, and what the chequerboard puts between them. The Journal of Antimicrobial Chemotherapy 52(1):1.
Crossref
|
|
|
|
|
Orchard A, VanVuuren S (2017). Commercial Essential Oils as Potential Antimicrobials to Treat Skin Diseases. In Evidence-based Complementary and Alternative Medicine 2017.
Crossref
|
|
|
|
|
Oulkheir S, Aghrouch M, El-Mourabit F, Dalha F, Graich H, Amouch F, Ouzaid K, Moukale A, Chadli S (2017). Antibacterial Activity of Essential Oils Extracts from Cinnamon, Thyme, Clove and Geranium Against a Gram Negative and Gram Positive Pathogenic Bacteria. Journal of Diseases and Medicinal Plants 3(1):1-5.
|
|
|
|
|
Patil VU, Gopal J, Singh BP (2012). Improvement for Bacterial Wilt Resistance in Potato By Conventional and Biotechnological Approaches. Agricultural Research 1(4):299-316.
Crossref
|
|
|
|
|
Paudel S, Dobhal S, Alvarez AM, Arif M (2020). Taxonomy and Phylogenetic Research on Ralstonia solanacearum Species Complex: A Complex Pathogen with Extraordinary Economic Consequences. Pathogens 9(11):886.
Crossref
|
|
|
|
|
Pavesi C, Banks LA, Hudaib T (2018). Antifungal and antibacterial activities of eugenol and non-polar extract of Syzygium aromaticum L. Journal of Pharmaceutical Sciences and Research 10(2):337-339.
|
|
|
|
|
Pei RS, Zhou F, Ji BP, Xu J (2009). Evaluation of combined antibacterial effects of eugenol, cinnamaldehyde, thymol, and carvacrol against E. coli with an improved method. Journal of Food Science 74(7):M379-M383.
Crossref
|
|
|
|
|
Perricone M, Arace E, Corbo MR, Sinigaglia M, Bevilacqua A (2015). Bioactivity of essential oils: A review on their interaction with food components. Frontiers in Microbiology 6(2):1-7.
Crossref
|
|
|
|
|
Pradhanang PM, Elphinstone JG, Fox RTV (2000). Sensitive detection of Ralstonia solanacearum in soil: A comparison of different detection techniques. Plant Pathology 49(4):414-422.
Crossref
|
|
|
|
|
Sarah MA, Lobna SEH (2017). Combined efficacy of thymol and silver nanoparticles against Staphylococcus aureus. African Journal of Microbiology Research 11(11):450-457.
Crossref
|
|
|
|
|
Sienkiewicz M, ?ysakowska M, Denys P, Kowalczyk E (2012). The antimicrobial activity of thyme essential oil against multidrug resistant clinical bacterial strains. Microbial Drug Resistance 18(2):137-148.
Crossref
|
|
|
|
|
Valliammai A, Selvaraj A, Yuvashree U, Aravindraja C, Karutha Pandian S (2020). sarA-Dependent Antibiofilm Activity of Thymol Enhances the Antibacterial Efficacy of Rifampicin Against Staphylococcus aureus. Frontiers in Microbiology 11(7):1-13.
Crossref
|
|
|
|
|
Xue J, Knoxville U (2015). Essential Oil Nanoemulsions Prepared with Natural Emulsifiers for Improved Food Safety. Doctoral Dissertations University of Tennessee, 194 pp.
View
|
|