Full Length Research Paper
ABSTRACT
Lead neurotoxicity has been studied in animal models to induce neurodegeneration. The study was aimed to evaluate the attenuating effects of quercetin on spatial memory impairment and degenerative changes in hippocampus of lead exposed rats. Thirty (30) Wistar rats were randomly divided into six groups of five rats per group (each, n = 5). Groups I (control) and II were administered distilled water and lead, respectively for 42 days. Lead was co-administered with Quercetin and Succimer for Groups III and VI, respectively for 42 days. Groups IV and V were administered lead for 21 days and treated with Quercetin for Group IV and Succimer for Group V for another 21 days. The administration was orally, once per day and lasted for 42 days. Spatial memory was assessed using Eight-arm radial water maze (8-ARWM) test. The rats were anaesthetized with Ketamine at 75 mg/kg intraperitoneally and euthanized. Brain was harvested, processed, Haematoxylin and Eosin stains were used for histo¬logical study of the hippocampus and cells were counted. Results showed spatial memory was impaired and histopatho¬logy such as vacuolation, pyknotic and reduction in pyramidal cells count in CA3 region of hippocampus of lead exposed rats. How¬ever, treat¬ment with quercetin improved spatial memory and histopatho¬logy. Thus, quercetin could be used as an antidote in lead neurotoxicity.
Key words: Lead, quercetin, memory, neurodegeneration.
INTRODUCTION
Neurode generation has been extensively studied in animal models (Colpo et al., 2017)as one of the main factors mediating neurodegenerative diseases like Parkinson’s disease and Alzheimer’s disease (Zhou et al., 2020). Exposure to various environmental toxicants (Mostafalou and Abdollahi, 2018)from anthropogenic activities (Almeida et al., 2019)like mining, smelting, crystal and ceramic industries, batteries production and recycling, coatings, plastic stabilizers and fertilizers, release heavy metals (Karri et al., 2020)that induced neuronal degeneration (Colpo et al., 2017). Lead is widely distributed (Zhou et al., 2020)and its exposure induces neurotoxicity from neurodevelopmental disorders (Feng et al., 2019)to severe neurodegenerative lesions (Zhou et al., 2020), leading to learning, memory and cognitive impairments (Feng et al., 2019). Lead can easily penetrates the neural tissue (Bressler and Goldstein, 1991), directly affecting the neurons and synapses or indirectly affecting neuronal connections via glial cells (Kasten-Jolly et al., 2011). The pathogenesis of lead-induced neurodegeneration are multifactorial, relating to cytotoxicity, changes in storage and release of neurotransmitters, energy metabolism disorders, oxidative stress, neuroinflammation and induction of apoptosis (Lidsky and Schneider, 2003). Previous study reported reduction in number of pyramidal neurons in degenerated sub-regions like CA1 and CA3 of the hippocampus mediated impaired activities of the hippocampus including storage and retrieval of information in lead exposed rats (Iliyasu et al., 2015). The earlier study of Khaled et al. (2014a) also reported neuronal damage in brain cortex and neurodegeneration of CA1 and CA3 regions of the hippocampus in lead exposed rats. In this region of the brain, scar formation, demyelination and neuronal atrophy (Soltaniniejad et al., 2003) could result from even low (0.2%) exposure to lead (Noor et al., 2012).
Succimer, also known as Meso-2,3-dimercaptosuccinic acid (DMSA), is a water-soluble and sulfhydryl containing compound which is an effective oral chelator of heavy metals (Aposhian, 1983). DMSA has a large therapeutic window and is the least toxic of the dithiol compounds (Graziano, 1986), and has been used since the 1950s as an antidote for lead poisoning in Russia, Japan, and the People’s Republic of China (Muckter et al., 1997).
Quercetin is a member of the flavonols under the class of flavonoids (Zaplatic et al., 2019). It has been investigated and found to be present in vegetables, fruits, and nuts (Guo and Bruno, 2015). Quercetin crosses blood brain barrier (BBB) and protective neural tissues effectively (Ossola et al., 2009). The neuroprotective mechanisms of quercetin are via metal chelation (Mira et al., 2002), direct scavenging of free radicals (Bindoli et al., 1985)and inhibition of Aβ aggregation (Zeng et al., 2004).
The scavenging activity of quercetin is due to the number of hydroxyl substitutions, correlating with its electron-donating ability (Boots et al., 2008). Quercetin hydroxyl group at C3 position is responsible for the inhibition of lipid peroxidation levels (Morel et al., 1993)and chelation of metal ions (Ishige et al., 2001). The three hydroxyl groups in A and B rings of quercetin are responsible for the quenching of the free radicals that cause oxidative stress (Husain et al., 1987). The study was aimed to evaluate the attenuating effects of quercetin on spatial memory impairment and degenerative changes in hippocampus of lead exposed rats.
MATERIALS AND METHODS
Acquisition of chemical substances/reagents/equipment
Quercetin (Sigma-Aldrich, USA), Succimer (Sigma-Aldrich, USA) and Lead (II) acetate trihydrate (Honeywell, Germany) were purchased.
Experimental animals
Thirty (30) male Wistar rats reared in Department of Pharmacology Animal House Centre, Ahmadu Bello University, Zaria, Nigeria, were used. The rats were housed in well ventilated plastic cages at room temperature in hygienic conditions under natural light and dark schedule and fed on standard laboratory diet for a period of two weeks and water was allowed ad libitum. Ethical clearance with the approval number of ABUCAUC/2018/087 was obtained from Committee on Animal Use and Care, Ahmadu Bello University, Zaria.
Experimental procedures
The oral LD50 of quercetin for rats is 161 mg/kg body weight (Sigma-Aldrich, 2018). Based on this report, 37% (60 mg/kg) of the oral LD50 was administered. The oral LD50 of Lead (II) acetate trihydrate from the result of the present acute toxicity study is 3808 mg/kg for Wistar rats. Based on this, 3.28% (125 mg/kg) of the LD50 was administered. The dosage of 10 mg/kg body of Succimer was administered as a standard drug according to the prescription of Alan and Miller (1998).
Experimental design
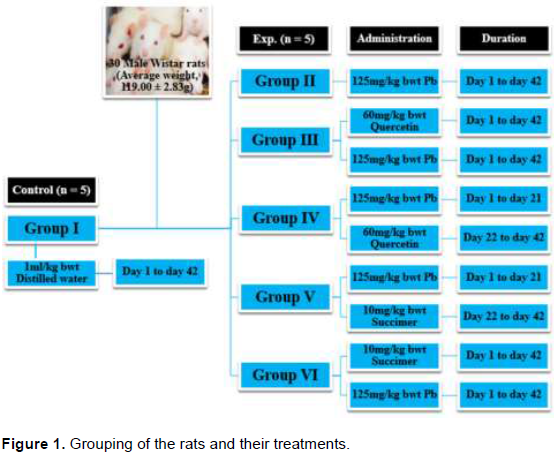
The 30 Wistar rats (average weight, 119.00 ± 2.83 g) were randomly divided into six groups (each, n = 5). Groups I (control) and II (Pb2+) were administered distilled water (H2O: 1 ml/kg) and Lead (Pb: 125 mg/kg), respectively for 42 days. Lead (Pb: 125 mg/kg) was co-administered (COA) with Quercetin (Q: 60 mg/kg) and Succimer (S: 10 mg/kg) for Groups III (Q + Pb2+ COA) and VI (S + Pb2+ COA), respectively for 42 days. In treatment (TRT) groups, Groups IV (Pb2+ + Q TRT) and V (Pb2+ + S TRT) were administered Pb: 125 mg/kg for 21 days and then treated with Q: 60 mg/kg for Group IV and S: 10 mg/kg for Group V for another 21 days. The administration was orally, once per day and lasted for 42 days (Figure 1).
Eight-arm radial water maze (8-ARWM) test
Spatial memory was assessed based on the protocol of Penley et al. (2013). The rats were trained before the administrations, the test was carried out during administrations, latencies to locate the platforms were recorded and average of the four trials was subjected to statistical analysis.
Animals sacrifice
The rats were euthanized by deeply anaesthetized with Ketamine at a dose of 75 mg/kg intraperitoneally (IP) (Molina et al., 2015). Brain was harvested and fixed in Bouin’s fluid. Tissues were processed and stained with Hematoxylin and Eosin, according to the methods of Sheehan and Hrapchak (1980).
Histopathological assessment
The sections were viewed under light microscope and photomicrographs were made using digital Am scope (MD-900) microscope camera. Pyramidal neurons in CA3 region of the hippocampus were counted using Digimizer image analysis software according to the previous reported methods (Iliyasu et al., 2015).
Data analysis
The data obtained were analyzed using One-way analysis of variance (ANOVA) to compare the mean differences followed by Tukey Post-hoc test where necessary. P-value less than 0.05 was considered significant. The results were analyzed and graphs were plotted using GraphPad Prism, Version 8.0.1.
RESULTS
Histopathological assessment of the hippocampus
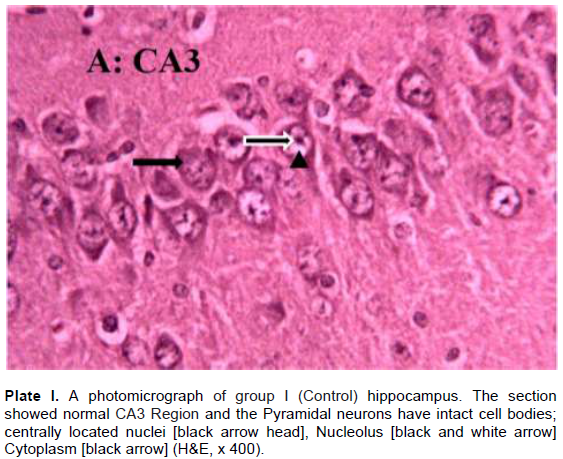
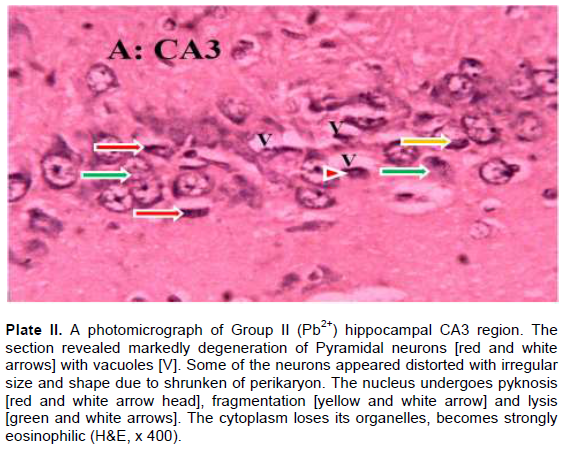
The results revealed that the CA3 region of hippocampus in Group I (control) had normal architecture of neurons whose cell bodies were all intact (Plate I). However, the CA3 region in group II: Pb2+ (Plate II) revealed histological alterations markedly in Pyramidal neurons with vacuoles. Some of the neurons appeared distorted with irregular size and shape due to shrunken of perikaryon. The nucleus undergoes pyknosis and the cytoplasm becomes strongly eosinophilic. The alterations in sections of hippocampus in groups: III: Q + Pb2+ COA (Plate III), IV: Pb2+ + Q TRT (Plate IV), V: Pb2+ + S TRT (Plate V), and VI: S + Pb2+ COA (Plate VI), were mild when compared with group II that revealed features of degeneration.
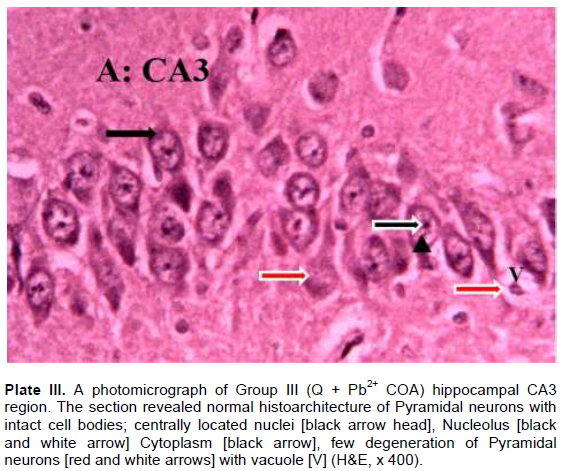
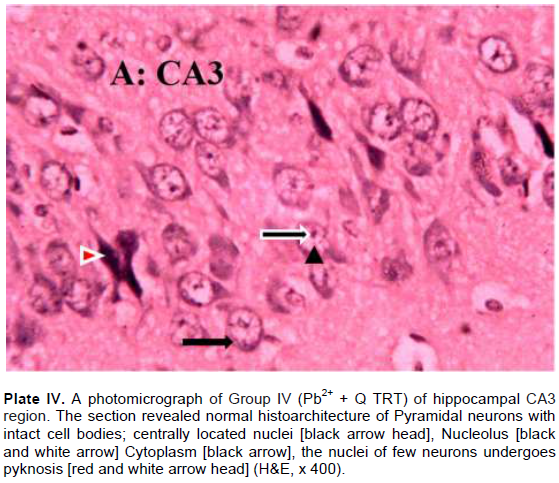
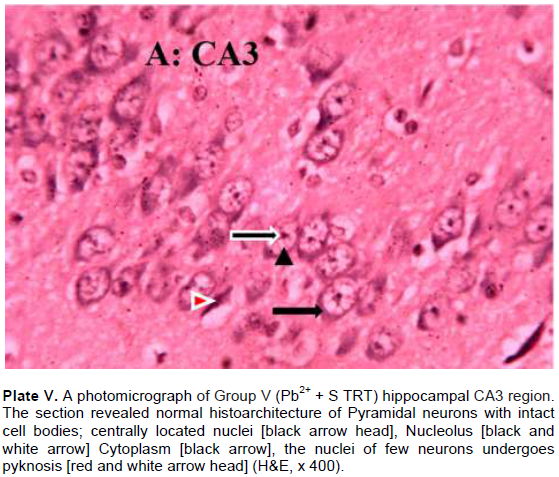
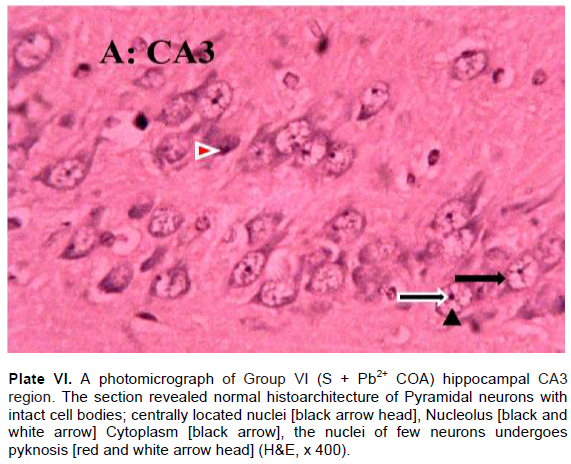
Pyramidal neurons count in CA3 region of the hippocampus
The results showed significant decrease in Pyramidal cell count hippocampal CA3 region in groups II: Pb2+ when compared with group I (control) at p < 0.05 (Figure 2). There were significant increases in Pyramidal cell count in groups: III: Pb2+ + Q TRT, IV: Pb2+ + Q TRT, V: Pb2+ + S TRT, and VI: S + Pb2+ COA, when compared with group II (p < 0.05). There was significant increase in Pyramidal cell count in group VI: S + Pb2+ COA when compared with groups: IV: Pb2+ + Q TRT and V: Pb2+ + S TRT (p < 0.05).
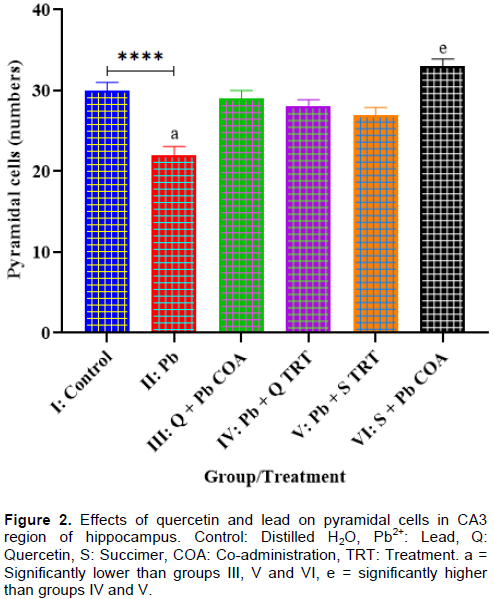
Effects of quercetin and lead on spatial memory
The mean latencies from the average of four trials to locate the platforms revealed insignificant differences across the groups in weeks I, II, III, IV and VI (p > 0.05) except in week V where the mean latency was significantly higher in group II (Pb2+) than Groups: IV (Pb2+ + Q TRT) and V (Pb2+ + S TRT) at p < 0.05 (Figure 3).
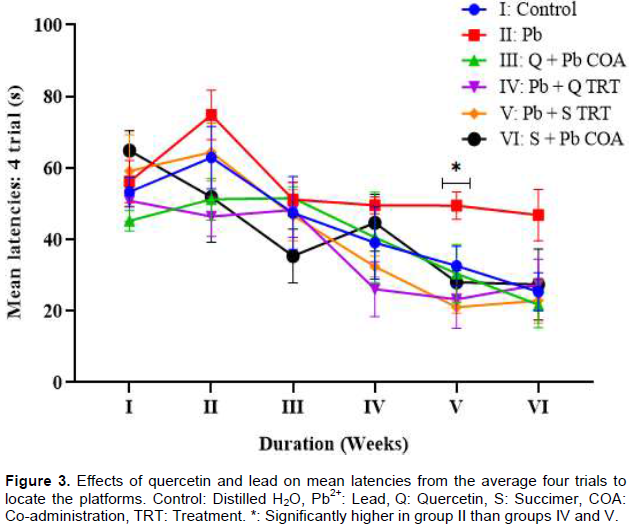
DISCUSSION
The histopathology revealed degeneration of CA3 region of hippocampus, manifested reduction in number of pyramidal cells in lead exposed rats. The present findings learn credence to previous study of Naqi-Syed (2014) who reported lead-induced behavioural impairments that could be attributed to hippocampal damage revealing the effects in degeneration and neuronal loss of pyramidal cells was evident, particularly in the deeper regions where empty spaces and vacuoles were encountered. Khaled et al. (2014b) had observed neuronal damage in brain cortex and hippocampus with neurodegeneration of CA1 and CA3 regions resulted from scar formation, demyelination and neuronal atrophy (Soltaniniejad et al., 2003) due to low levels of lead exposure (Noor et al., 2012). Zook et al. (1980) have reported gliosis and necrosis of hippocampal neurons in lead exposure rats. Brinck and Wechsler (1985) incubated slices of guinea pig hippocampus in a lead containing medium and reported vacuolation and shrinkage of pyramidal cells in the outer parts of the CA4 neurons. Loss of cellular integrity in CA3 region of the hippocampus may be accountable for the clinical features of lead such as disorders in emotional response, memory and learning (Schneider et al., 2012). The effects of lead induced alterations in nerve fibers and overall myelin structures were associated with a decline in cognitive function (Peters, 1996) due to decrease in propagation of impulse along nerve fibers (Bondan et al., 2006). The study revealed the attenuating effects of quercetin and succimer (standard drug) against lead-induced histological alterations in CA3 region of hippocampus in lead exposed rats. However, the effects of succimer were more potent than quercetin especially in pyramidal neurons count. Quercetin treatment reduced histopathological alterations on neurons, axon structure, density and morphology induced by sodium arsenate (Mesram et al., 2019). Similar observation was reported by Flora et al. (2014), following co-administration of quercetin with arsenic that quercetin treatment showed improvement in overall histoarchitecture of brain, with normal sized glial cells, pyramidal cells with mild pyknosis and cytoplasmic vacuolization and thus increasing the number of neurons. Mesram et al. (2019)reported that quercetin antioxidant properties against toxicants could ameliorate histological alterations (Yuhan et al., 2012; Jiang et al., 2016).
The study also revealed impairment in spatial memory by increasing the latency to locate the platforms in 8-arm radial water maze (8-ARWM) in lead exposed rats. The increase in latency is an indicator of memory impairment, being significant and correlated with neuropathology observed, indicating hippocampal degeneration in lead exposed rats. The observations are in agreement with earlier study that lead exposure in Wistar rats induced hippocampal damage and impaired learning abilities in Morris Water Maze (MWM) test (Iliyasu et al., 2019). The interference of lead with developing nervous system can cause permanent learning and behavioural disorders, linking the induction of neurodegeneration via free radicals generated from lead and other heavy metals exposure (Sasaki et al., 2003; Heo and Lee, 2004).
The spatial learning and memory require hippocampal N-methyl-D-aspartate (NMDA) receptor (Ohno et al., 1992; McNamera and Skelton, 1993) and alteration of this receptor due to lead exposure damage mitochondria, microfilaments, and microtubules in hippocampal neurons and myelin sheath degeneration (Jett and Guilarte, 1995; Xu et al., 2009). Neurogenesis and processes of differentiation of new cells in the dentate gyrus of hippocampus are inhibition in during early development exposure to lead (Jaako-Movits et al., 2005), and thus could be responsible for the spatial memory impairment. The hippocampus and prefrontal cortex alterations might be the basis of the learning and memory impairments induced by chronic lead exposure (Mansouri et al., 2013). The study however, revealed the attenuating effects of quercetin and succimer (standard drug) on improving spatial memory impairment by reducing the latency to locate the platforms in lead-exposed rats. Khan et al. (2018) demonstrated the ameliorative effect of quercetin against lipopolysaccharide (LPS)-induced neuroinflammation mediated neurodegeneration and memory deficits in the cortical and hippocampal regions of the adult mouse brain. Memory improvement was also observed by Patil et al. (2003) after quercetin administration intraperitoneally for seven days in mice suffering from age-related or lipopolysaccharide treatment-induced cognitive impairment. Quercetin at a dose of 50 mg/kg improved spatial memory impairment in the 8-arm radial maze test and reduced neuronal cell death in the hippocampal CA1 area induced by repeated cerebral ischemia (Fengling et al., 2007). Nageshwar et al. (2017) reported the neuroprotective and memory enhancing effects of quercetin by inhibiting sodium fluoride-induced neurodegeneration in developing brain of rat. Quercetin possesses multiple neuroprotective activities, such as suppression of neuroinflammation and neuronal apoptosis, promoting neuronal survival, as well as improving learning and memory functions (Dey et al., 2017; Ali et al., 2018; Shal et al., 2018). The activation of CREB and induction of neurogenesis by quercetin enhances learning and memory as a compensatory mechanism for neuronal cell death in the brain of Aβ1-42 injected rats (Karimipour et al., 2019). The recovery of memory by quercetin after the induction of lesions in the brain have been described for its ability to activate signaling pathways and to induce vascular effects
capable of promoting new nerve cell growth in the hippocampus (Spencer, 2009).
CONCLUSION
The study concluded that the effects of quercetin and succimer (standard drug) in lead exposed rats, improved memory impairment and degenerative changes in hippocampus when compared with lead exposed rats only. Thus, quercetin could be a potential antidote in lead neurotoxicity.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors are grateful for the provision of reagents from Histology Laboratory of the Department of Human Anatomy, Ahmadu Bello University, Zaria-Nigeria and support of the management of Kogi State University, Anyigba, Nigeria.
REFERENCES
|
Alan L, Miller ND (1998). Dimercaptosuccinic Acid (DMSA), A Non-Toxic, Water-Soluble Treatment For Heavy Metal Toxicity. Alternative Medicine Review 3(3):199-207. |
|
|
Ali T, Kim T, Rehman SU, Khan MS, Amin FU, Khan M, Kim MO. (2018). Natural dietary supplementation of anthocyanins via PI3K/Akt/Nrf2/HO-1 pathways mitigate oxidative stress, neurodegeneration, and memory impairment in a mouse model of Alzheimer's disease. Molecular Neurobiology 55(7):6076-6093. |
|
|
Almeida LACB, deCunha AM, daUrbano MR, Buzzo ML, Camargo AEI, Peixe TS, Aschner M, Barbosa F, daSilva AMR, Paoliello MMB (2019). Blood reference values for metals in a general adult population in southern Brazil. Environmental Research 177:108646. |
|
|
Aposhian HV (1983). DMSA and DMPS Water soluble antidotes for heavy metal poisoning. The Annual Review of Pharmacology and Toxicology 23(1):193-215. |
|
|
Bindoli A, Valente M, Cavallini L (1985). Inhibitory action of quercetin on xanthine oxidase and xanthine dehydrogenase activity. Pharmacological Research Communications 17(9):831-839. |
|
|
Bondan EF, Lallo MA, Trigueiro AH, Ribeiro CP, Sinhorini IL, Graca DL (2006). Delayed Schwann cell and oligodendrocyte remyelination after ethidium bromide injection in the brainstem of Wistar rats submitted to streptozotocin diabetogenic treatment. Brazilian Journal of Medical and Biological Research (5):637-646. |
|
|
Boots AW, Haenen GR, Bast A (2008). Health effects of quercetin: from antioxidant to nutraceutical. European Journal of Pharmacology 585(2-3):325-337. |
|
|
Bressler JP, Goldstein GW (1991). Mechanisms of lead neurotoxicity. Biochemical Pharmacology 41(4):479-484. |
|
|
Brinck U, Wechsler W (1985). Microscopic examination of hippocampal slices after short term lead exposure in vitro. Neurotoxicology and Teratology 11(6):539-543. |
|
|
Colpo GD, Ribeiro FM, Rocha NP, Teixeira, AL (2017). Chapter 42-Animal Models for the Study of Human Neurodegenerative Diseases. In Animal Models for the Study of Human Disease, Academic Press. pp. 1109-1129. |
|
|
Dey A, Bhattacharya R, Mukherjee A, Pandey DK (2017). Natural products against Alzheimer's disease: pharmaco-therapeutics and biotechnological interventions. Biotechnology Advances 35(2):178-216. |
|
|
Feng C, Liu S, Zhou F, Gao Y, Li Y, Du G, Chen Y, Jiao H, Feng J, Zhang Y, Bo D, Li Z, Fan G (2019). Oxidative stress in the neurodegenerative brain following lifetime exposure to lead in rats: Changes in lifespan profiles. Toxicology 411:101-109. |
|
|
Fengling P, Kenichi M, Keiichi I, Kyouko M, Yurika T, Kensuke O, Takashi E, Yoshihisa K, Nobuaki E, Katsunori I, Michihiro F (2007). Neuroprotective Effects of Quercetin and Rutin on Spatial Memory Impairment in an 8-Arm Radial Maze Task and Neuronal Death Induced by Repeated Cerebral Ischemia in Rats. Journal of Pharmacological Sciences 104(4):329-334. |
|
|
Flora SJS, Dwivedi N, Deb U, Kushwaha P, Lomash V (2014). Effects of co- exposure to arsenic and dichlorvos on glutathione metabolism, neurological, hepatic variables and tissue histopathology in rats. Toxicology Research 3(1):23-31. |
|
|
Graziano JH (1986). Role of 2,3-dimercaptosuccinic acid in the treatment of heavy metal poisoning. Medical Toxicology 1(3):155-162. |
|
|
Guo Y, Bruno RS (2015). Endogenous and exogenous mediators of quercetin bioavailability. Journal of Nutritional Biochemistry 26(3):201-210. |
|
|
Heo HJ, Lee CY (2004). Protective Effects of Quercetin and Vitamin C against Oxidative Stress-Induced Neurodegeneration. Journal of Agricultural and Food Chemistry 52(25):7514-7517. |
|
|
Husain RS, Cillard J, Cillard P (1987). Hydroxyl radical scavenging activity of flavonoids. Phytochemistry 26(9):2489-2491. |
|
|
Iliyasu MO, Ibegbu AO, Sambo JS, Musa SA, Akpulu PS (2015). Histopathological changes on the hippocampus of adult Wistar rats exposed to lead acetate and aqueous extract of Psidium guajava leaves. African Journal of Cellular Pathology 5:26-31. |
|
|
Iliyasu MO, Ibegbu AO, Sambo SJ, Musa SA, Akpulu SP, Animoku AA, Mustapha M (2019). The Study of the Behaviour, the Hippocampus and Cerebellar Cortex of Adult Wistar Rats Exposed to Lead and Treatment with Psidium guajava Leaf Extract. IBRO Reports 6:S54-S345. |
|
|
Ishige K, Schubert D, Sagara Y (2001). Flavonoids protect neuronal cells from oxidative stress by three distinct mechanisms. Free Radical Biology and Medicine 30(4):433-446. |
|
|
Jaako-Movits K, Zharkovsky T, Romantchik O (2005). Developmental lead exposure impairs contextual fear conditioning and reduces adult hippocampal neurogenesis in the rat brain. International Journal of Developmental Neuroscience 23(7):627-635. |
|
|
Jett DA, Guilarte TR (1995). Developmental lead exposure alters N-methyl-D-aspartate and muscarinic cholinergic receptors in the rat hippocampus: An autoradiographic study. Neurotoxicology 16(1):7-18. |
|
|
Jiang W, Huang Y, Han N, He F, Li M, Bian Z, Liu J, Sun T, Zhu L (2016). Quercetin suppresses NLRP3 inflammation activation and attenuates histopathology in a rat model of spinal cord injury. Spinal Cord 54(8):592-596. |
|
|
Karimipour M, Rahbarghazi R, Tayefi H, Shimia M, Ghanadian M, Mahmoudi J, Bagheri HS (2019). Quercetin promotes learning and memory performance concomitantly with neural stem/progenitor cell proliferation and neurogenesis in the adult rat dentate gyrus. International Journal of Developmental Neuroscience 74:18-26. |
|
|
Karri V, Schuhmacher M, Kumar V (2020). A systems toxicology approach to compare the heavy metal mixtures (Pb, As, MeHg) impact in neurodegenerative diseases. Food and Chemical Toxicology 139:111257. |
|
|
Kasten-Jolly J, Heo Y, Lawrence DA (2011). Central nervous system cytokine gene expression: Modulation by lead. Journal of Biochemical and Molecular Toxicology 25(1):41-54. |
|
|
Khaled R, Khaled H, Mubarak A, Rudolf M, Wolf-Dieter R (2014a). Thymoquinone ameliorates lead-induced brain damage in Sprague Dawley rats. Experimental and Toxicologic Pathology 66(1):13-17. |
|
|
Khan A, Ali T, Rehman SU, Khan MS, Alam SI, Ikram M, Muhammad T, Saeed K, Badshah H, Kim MO (2018). Neuroprotective Effect of Quercetin against the Detrimental Effects of LPS in the Adult Mouse Brain. Frontiers in Pharmacology 9:1383. |
|
|
Lidsky TI, Schneider JS (2003). Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 126(1):5-19. |
|
|
Mansouri MT, Naghizadeh B, López-Larrubia P, Cauli O (2013). Behavioral deficits induced by lead exposure are accompanied by serotonergic and cholinergic alterations in the prefrontal cortex. Neurochemistry International 62(3):232-239. |
|
|
McNamera RK, Skelton RW (1993). The neuropharmacological and neurochemical basis of place learning in the Morris water maze. Brain Research Reviews 18(1):33-49. |
|
|
Mesram N, Umamaheshwari P, Reddy KP (2019). Quercetin reverses sodium arsenate induced oxidative stress, behavioural and histological alterations in brain of rat. Journal of Pharmaceutical Sciences and Research 11(6):2267-2274. |
|
|
Mira L, Fernandez MT, Santos M, Rocha R, Florêncio MH, Jennings KR (2002). Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radical Research 36(11):1199-1208. |
|
|
Molina HH, Reznick AZ, Kaufman H, Aizenbud D (2015). Periodontal cytokines profile under orthodontic force and extracorporeal shock wave stimuli in a rat model. Journal of Periodontal Research 50(3):389-396. |
|
|
Morel I, Lescoat G, Cogrel P, Sergent O, Pasdeloup N, Brissot P, Cillard P, Cillard J (1993). Antioxidant and iron-chelating activities of the flavonoids catechin, quercetin and diosmetin on iron-loaded rat hepatocyte cultures. Biochemical Pharmacology 45(1):13-19. |
|
|
Mostafalou S, Abdollahi M (2018). The link of organophosphorus pesticides with neurodegenerative and neurodevelopmental diseases based on evidence and mechanisms. Toxicology 409(1):44-52. |
|
|
Muckter H, Liebl B, Reichl FX (1997). Are we ready to replace dimercaprol (BAL) as an arsenic antidote? Human Experimental Toxicology 16(8):460-465. |
|
|
Nageshwar M, Kirankumar N, Rajkiran RB, Chandrashakar RN, Pratap RK (2017). Quercetin treatment against NaF-induced oxidative stress related neuronal and learning changes in developing rats. Journal of King Saud University Science 29(2):221-229. |
|
|
Naqi-Syed SZ (2014). Pyramidal layer thinning, shrunken neurons and deep vacuolation in hippocampus due to the organic lead induced toxicity. International Journal of Anatomy and Research 2(2):390-93. |
|
|
Noor AWA, Teoh SL, Srijit D, Mohamad FY, Azian AL, Taty AK (2012). Histological observation of the hippocampus and frontal cortex of experimental Sprague-Dawley rats fed with lead in a dose related manner. British Journal of Medical and Health Sciences 1(1):99-106. |
|
|
Ohno M, Yamamoto T, Watanabe S (1992). Effects of intrahippocampal injection of N -Methyl- d-Aspartate receptor antagonists and scopolamine on working and reference memory assessed in rats by a three-panel runway task. Journal of Pharmacology and Experimental Therapeutics 263:943-950. |
|
|
Ossola B, Kaariainen TM, Mannisto PT (2009). The multiple faces of quercetin in neuroprotection. Expert Opinion on Drug Safety 8(4):397-409. |
|
|
Patil CS, Singh VP, Satyanarayan PS, Jain NK, Singh A, Kulkarni SK (2003). Protective effect of flavonoids against aging and lipopolysaccharide-induced cognitive impairment in mice. Pharmacology 69(2):59-67. |
|
|
Penley SC, Gaudet CM, Threlkeld SW (2013). Use of an eight-arm radial water maze to assess working and reference memory following neonatal brain injury. Journal of Visualized Experiments 82(82):1-7. |
|
|
Peters A (1996). Age-related changes in oligodendrocytes in monkey cerebral cortex. Journal of Comparative Neurology 371(1):153-163. |
|
|
Sasaki N, Toda T, Kaneko T, Naomichi B, Mitsuyoshi M (2003). Protective effects of flavonoids on the cytotoxicity of linoleic acid hydroperoxide toward rat pheochromocytoma PC12 cells. Chemico-Biological Interactions 145(1):101-16. |
|
|
Schneider JS, Mettil W, Anderson DW (2012). Differential effect of postnatal lead exposure on gene expression in the hippocampus and frontal cortex. Journal of Molecular Neuroscience 47(1):76-88. |
|
|
Shal B, Ding W, Ali H, Kim YS, Khan S (2018). Anti-neuroinflammatory potential of natural products in attenuation of Alzheimer's disease. Frontiers in Pharmacology 9:548. |
|
|
Sheehan D, Hrapchak B (1980). Theory and practice of histotechnology. 2nd Ed, Battelle Press, Ohio pp. 235-237. |
|
|
Sigma-Aldrich Co (2018). Quercetin, Q4951. Material Safety Data Sheet, Version 4.8. Chemie GmbH Riedstrasse 2:1-7. |
|
|
Soltaniniejad K, Kebriaeezadeh A, Minaiee B, Ostad SN, Hosseini R, Azizi E, Abdollahi M (2003). Biochemical and ultrastructural evidences for toxicity of lead through free radicals in rat brain. Human and Experimental Toxicology 22(8):417-423. |
|
|
Spencer JPE (2009). Flavonoids and brain health: Multiple effects underpinned by common mechanisms. Genes and Nutrition 4(4):243-250. |
|
|
Xu J, Yan HC, Yang B, Tong LS, Zou YX, Tian Y (2009). Effects of lead exposure on hippocampal metabotropic glutamate receptor subtype 3 and 7 in developmental rats. Journal of Negative Results in BioMedicine 8(1):5. |
|
|
Yuhan T, Chao G, Mingyou X, Yanyan L, Liping Z, Di W, Xuefeng Y, Liegang L, Ping Y (2012). Quercetin prevents ethanolinduced dyslipidemia and mitochondrial oxidative damage. Food and Chemical Toxicology 50(5):1194-1200. |
|
|
Zaplatic E, Bule M, Zahid S, Shah A, Uddin S, Niaz K (2019). Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer's disease. Life Sciences 224:109-119. |
|
|
Zeng H, Chen Q, Zhao B (2004). Genistein ameliorates β-amyloid peptide (25-35)-induced hippocampal neuronal apoptosis. Free Radical Biology and Medicine 36(2):180-188. |
|
|
Zhou F, Du G, Xie J, Gu J, Jia Q, Fan Y, Yu, H, Zha Z, Wang K, Ouyang L, Shao L, Feng C, Fan G (2020). RyRs mediate lead-induced neurodegenerative disorders through calcium signaling pathways. Science of the Total Environment 701:134901. |
|
|
Zook BC, London WT, Wilpizeski CR, Sever JL (1980). Experimental lead paint poisoning in nonhuman primates. III. Pathologic findings; Brain Research 189(2):369-376. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


