Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
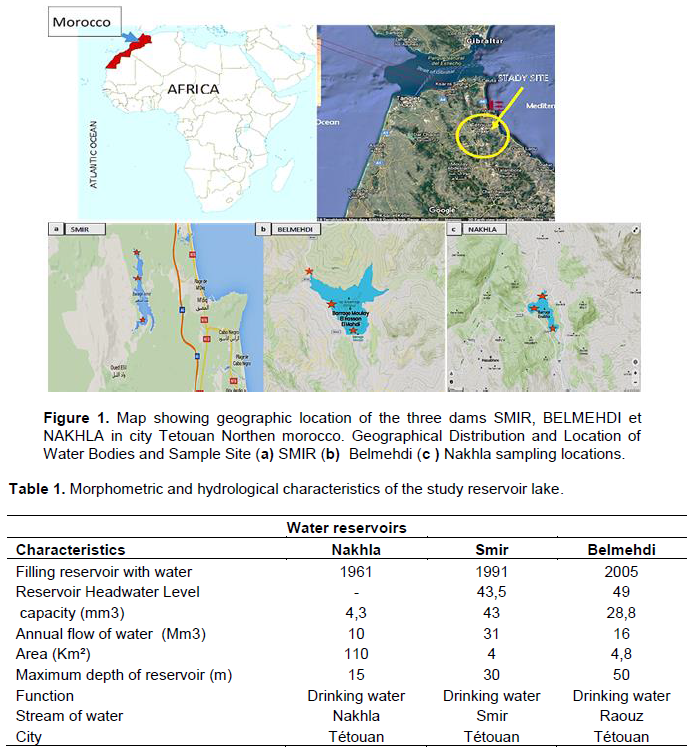
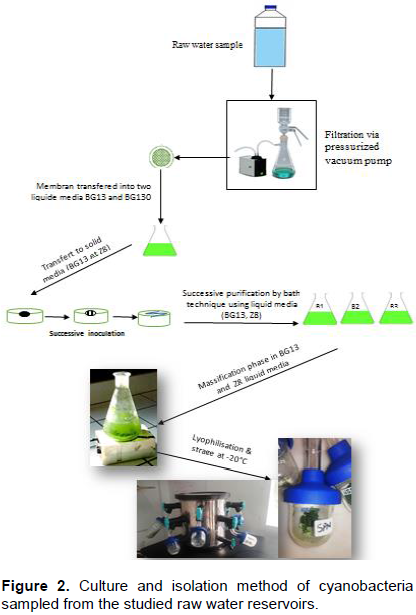
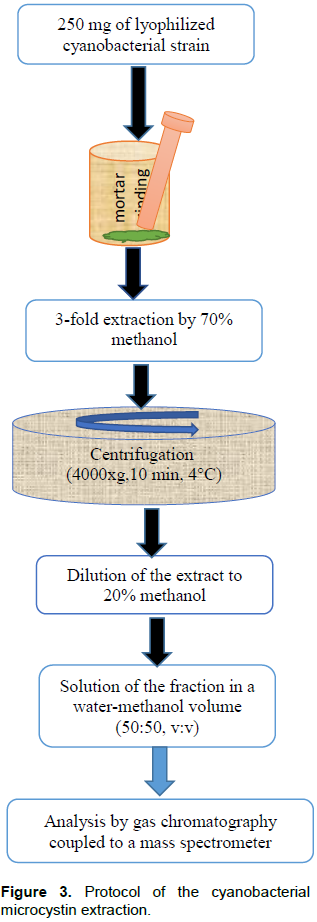
RESULTS

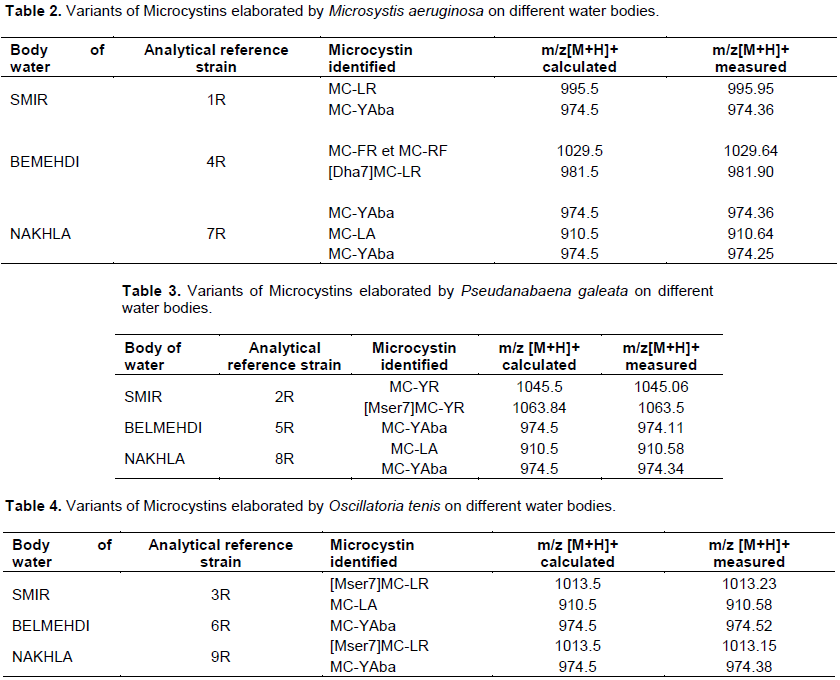
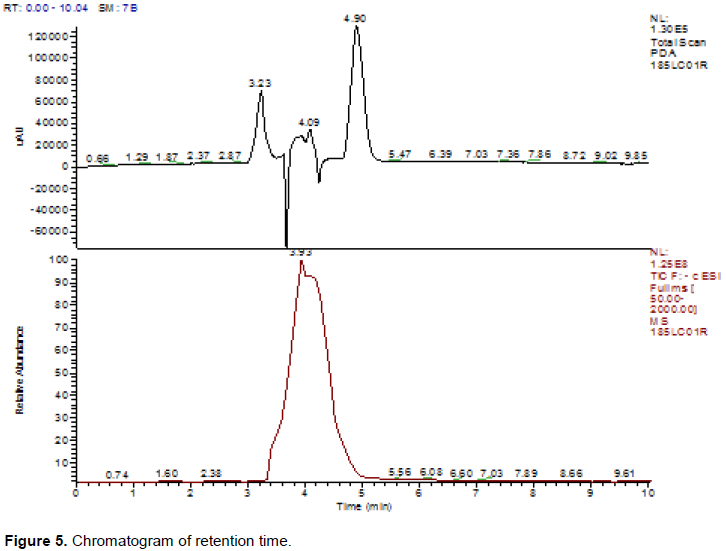
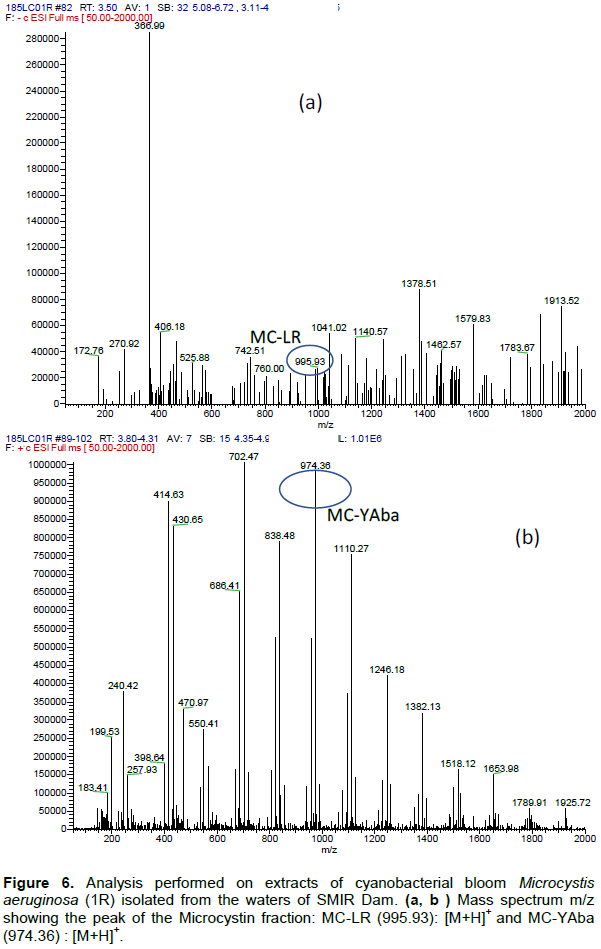
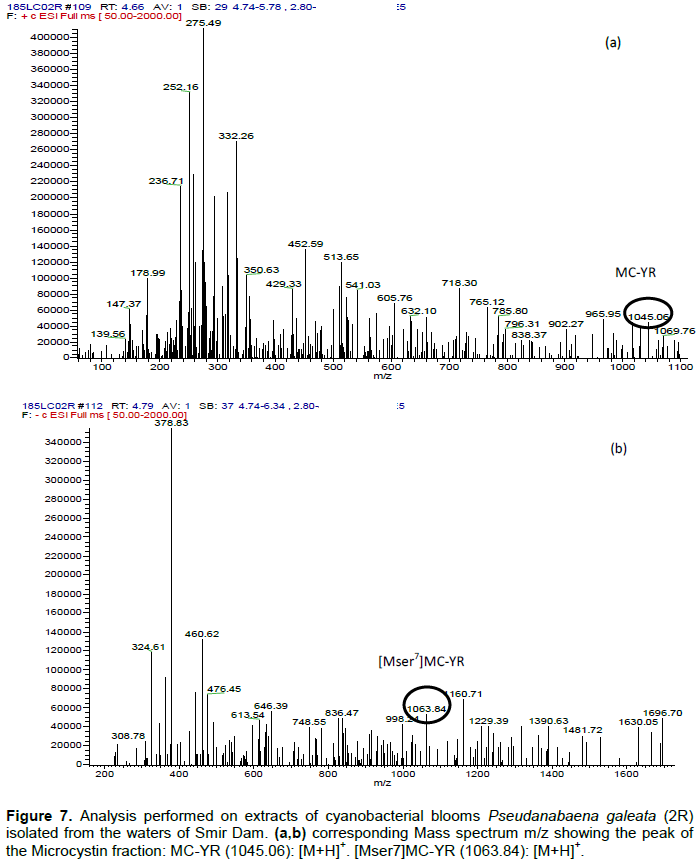
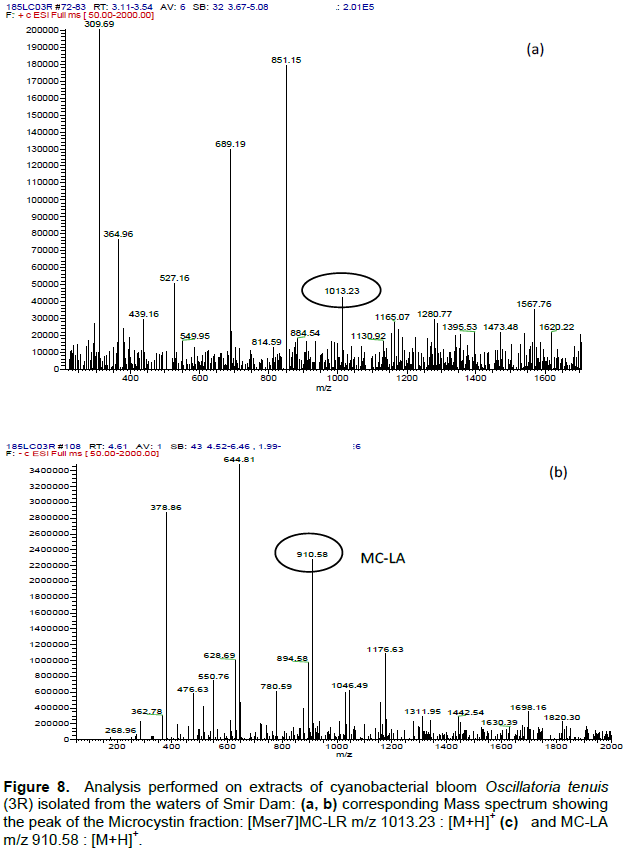
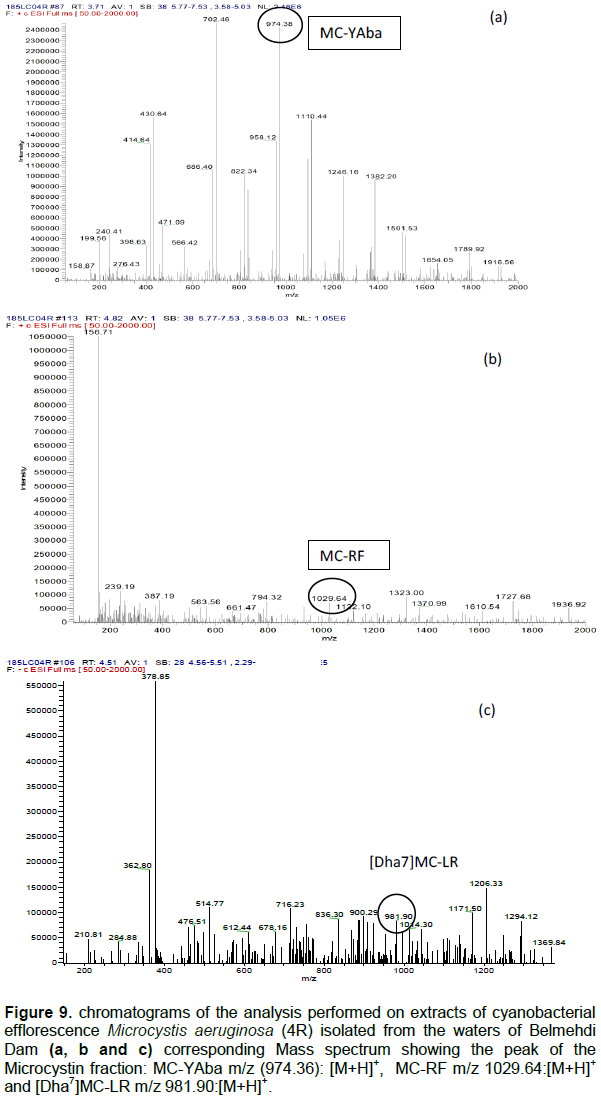
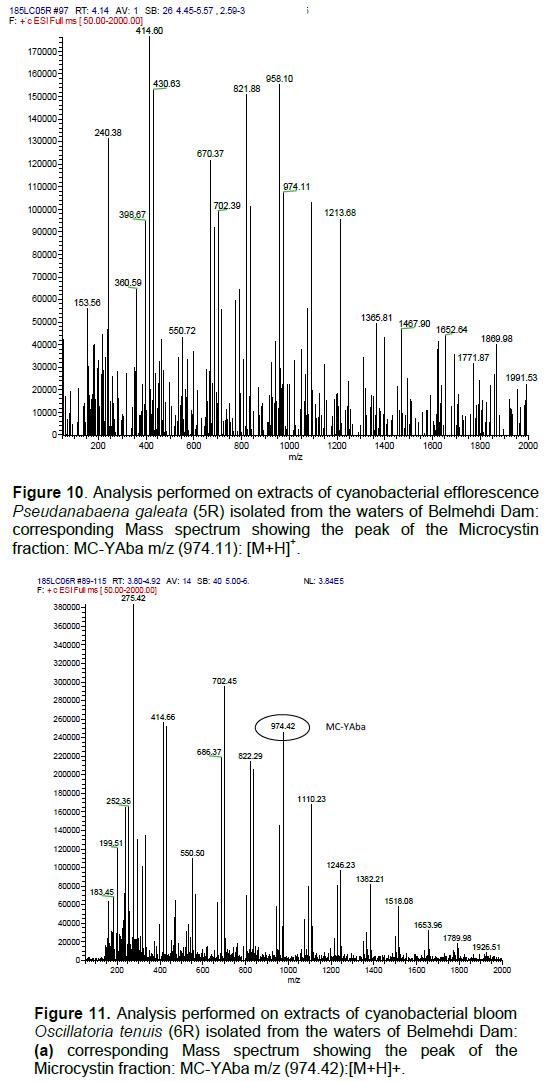
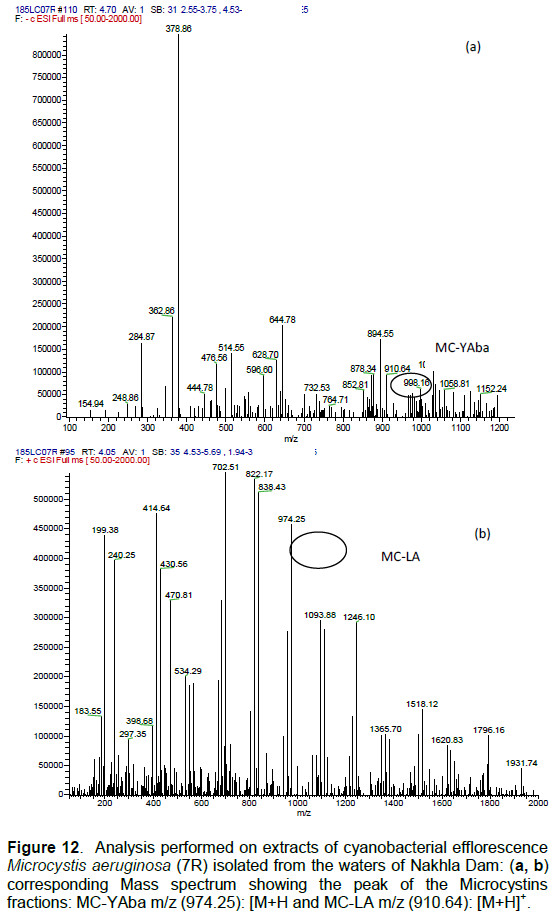
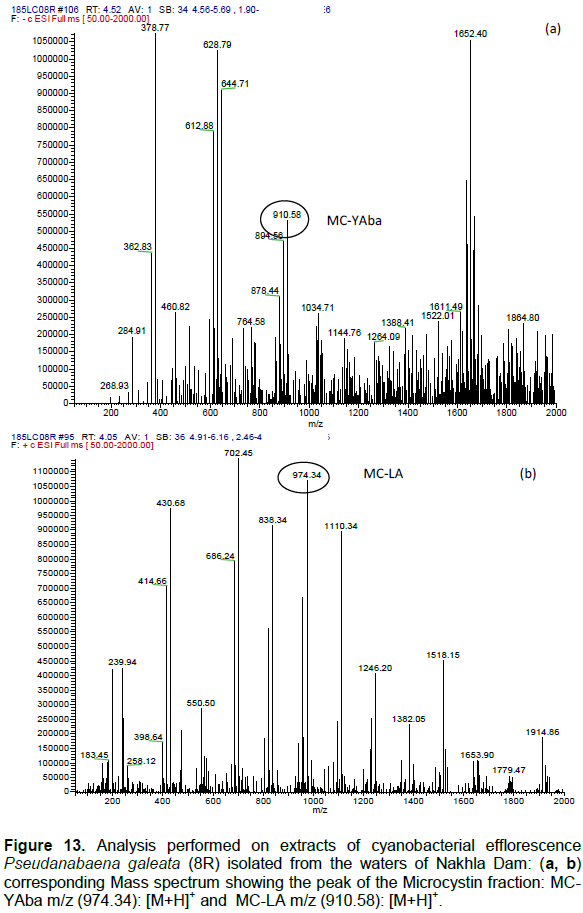
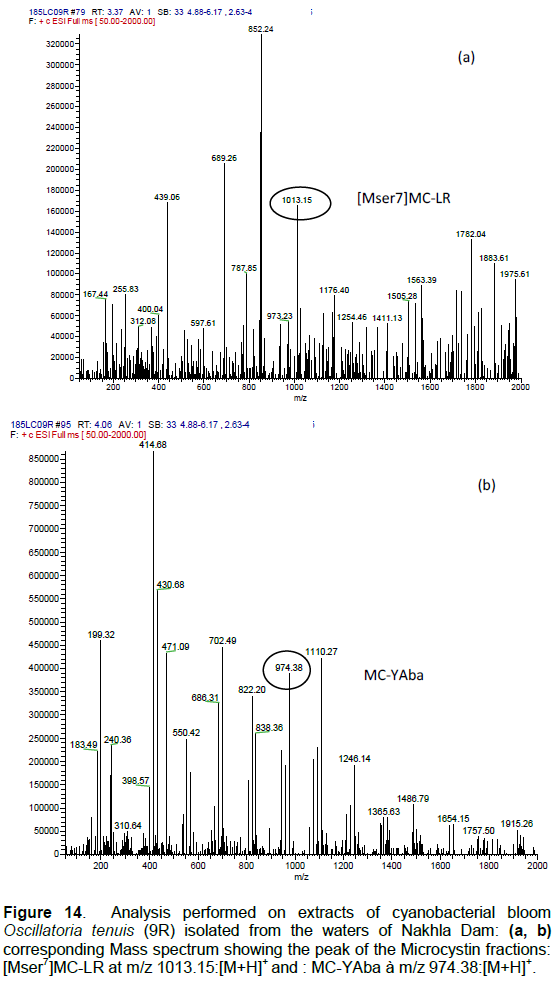
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
ADH (2006). Les grands barrages au Maroc. Publication de l'administration de l'hydraulique, Rabat, Maroc 123 p. |
|
|
Ai Y, Lee S, Lee J (2020). Drinking water treatment residuals from cyanobacteria bloom-affected areas: Investigation of potential impact on agricultural land application. Science of the Total Environment 706:1-11. |
|
|
Bourrelly P (1985). Les algues d'eau douce: Initiation à la systématique. Tome III: Les algues bleues et rouges. Les Eugléniens, Péridiniens et Cryptomonadines. Société Nouvelle des Editions Bouée. |
|
|
Bruno MP, Gucci PMB, Pierdominici E, Sestili P, Ioppolo A, Sechi N, Volterra L (1992). Microcystin-like toxins in different freshwater species of Oscillatoria. Toxicon 30(10):1307-1311. |
|
|
Carmichael WW (1992a). Cyanobacteria secondary metabolites the cyanotoxins. Journal of Applied Bacteriology 72(6):445-459. |
|
|
Carmichael WW (1995). Toxic Microcystis and the environment. Toxic microcystis pp.1-11. |
|
|
Carmichael WW, Azevedo SM, An JS, Jochimsen EM, Lau S, Eaglesham GK (2001). Human fatalities from cyanobacteria: chemical and biological evidence for cyanotoxins. Environmental Health Perspectives 109(7):663-668. |
|
|
Cherifi O, Loudiki M (2002).Variations de la structure du lac-réservoir oligotrophe Bin El Ouidane (Maroc). Revue des Sciences de l'Eau 15(1):193-208. |
|
|
Chorus I, Bartram J, Editors (1999). Toxic Cyanobacteria in Water-A Guide to Their Public Health Consequences, Monitoring, and Management, E & FN Spon, published on behalf of the World Health Organization, New York. |
|
|
Christophoridis C, Zervou SK, Manolidi K, Katsiapi M, Moustaka-Gouni M, Kaloudis T, Hiskia A (2018). Occurrence and diversity of cyanotoxins in Greek lakes. Scientific Reports 8(1):1-22. |
|
|
Codd GA (2000). Cyanobacterial toxins, the perception of water quality, and the prioritisation of eutrophication control. Ecological Engineering 16(1):51-60. |
|
|
Diehnelt CW, Dugan NR, Peterman SM, Budde WL (2006). Identification of Microcystin Toxins from a Strain of Microcystis aeruginosa by Liquid Chromatography Introduction into a Hybrid Linear Ion Trap-Fourier Transform Ion Cyclotron Resonance Mass Spectrometer. Analytical Chemistry 78(2): 501-512. |
|
|
Diehnelt CW, Peterman SM, Budde WL (2005). Liquid chromatography-tandem mass spectrometry and accurate m/z measurements of cyclic peptide cyanobacteria toxins. Trends in Analytical Chemistry 24(7):622-634. |
|
|
Dittmann E, Börner T (2005). Genetic contributions to the risk assessment of microcystin in the environment. Toxicology and Applied Pharmacology 203(3):192-200. |
|
|
Dittmann E, Neilan BA, Erhard M, Von Döhren H, Börner T (1997). Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Molecular microbiology 26(4):779-787. |
|
|
Douma M, Ouahid Y, del Campo FF, Loudiki M, Mouhri Kh, Oudra B (2010). Identification and quantification of cyanobacterial toxins (microcystins) in two Moroccan drinking-water reservoirs (Mansour Eddahbi, Almassira). Environmental Monitoring and Assessment 160(1-4):439-450. |
|
|
El Ghachtoul Y, Alaoui Mhamidi M, Gabi H (2005). Eutrophisation des eaux des retenues des barrages Smir et Sehla (maroc): causes, conséquences et consignes de gestion. Revue des Sciences de l'Eau 18:75-89. |
|
|
Ferris JM, Hirsch CF (1991). Method for isolation and purification of Cyanobacteria. Applied and Environmental Microbiology 57(5):1448-1452. |
|
|
Filatova D, Picardo M, Núñez O, Farré M (2020). Analysis, levels and seasonal variation of cyanotoxins in freshwater ecosystems. Trends in Environmental Analytical Chemistry 26:e00091. |
|
|
Gophen M (2021). Climate Change-Enhanced Cyanobacteria Domination in Lake Kinneret: A Retrospective Overview. Water 13(2):163. |
|
|
Hayakawa K, Tsujimura S, Napolitano GE, Nakano SI, Kumagai M, Nakajima T, Jiao C (2002). Fatty acid composition as an indicator of physiological condition of the cyanobacterium miicrocystis aeruginosa. Limnology 3(1):29-35. |
|
|
Hirooka T, Akiyama Y, Tsuji N, Nakamura T, Nagase H, Hirata K, Miyamoto K (2003). Removal of hazardous phenols by microalgae under photoautotrophic conditions. Journal of Bioscience and Bioengineering 95(2):200-203. |
|
|
Hotto AM, Satchwell MF, Boyer GL (2007). Molecular characterization of potential microcystin-producing cyanobacteria in Lake Ontario embayments and nearshore waters. Applied and Environmental Microbiology 73(14):4570-8. |
|
|
Huisman J, Codd GA, Paerl HW, Ibelings BW, Verspagen JM, Visser PM (2018). Cyanobacterial blooms. Nature Reviews Microbiology 16(8):471-83. |
|
|
IPCC (2007). Climate change 2007: the physical sciences basis. Contribution of the Working Group I to the Fourth Assessment report of The IPCC [Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averty KB, Tignor M, Miller HL (eds). Cambridge University Press, Cambridge, United Kingdom and New York, USA. |
|
|
Issaka S, Ashraf MA, (2017). Impact of soil erosion and degradation on water quality: a review. Geology, Ecology and Landscapes 1(1):1-11. |
|
|
Jacoby JM, Kann J (2007). The occurrence and response to toxic cyanobacteria in the Pacific Northwest, North America. Lake and Reservoir Management 23(2):123-143. |
|
|
Kadiri MO, Isagba S, Ogbebor JU, Omoruyi OA, Unusiotame-Owolagba TE, Lorenzi AS, Chia MA (2020). The presence of microcystins in the coastal waters of Nigeria, from the Bights of Bonny and Benin, Gulf of Guinea. Environmental Science and Pollution Research 27(28):35284-35293. |
|
|
Kenyon CN, Stanier RY (1970). Possible evolutionary significance of Polyunsaturated fatty acids in blue-green algae. Nature 227(5263):1164-1166. |
|
|
Komárek J (2016). Review of the cyanobacterial genera implying planktic species after recent taxonomic revisions according to polyphasic methods: state as of 2014. Hydrobiologia 764 (1):259-270. |
|
|
Komárek J, Anagnostidis K (2005). Cyanoprokaryota, freshwater flora of Central Europe, Gustav Fischer, Jena Stuttgart Lübeck Ulm 548 p. |
|
|
Kotai J (1972). Instructions for preparation of modified nutrient solution Z8 for algae. Norwegian Institute for Water Research, Oslo 11(69):5. |
|
|
Lawton LA, Edwards C, Codd, GA (1994). Extraction and high-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst 119(7):1525-1530. |
|
|
Lindholm T, Eriksson JE, Meriluoto JAO (1989). Toxic cyanobacteria and water quality problems: Examples from a eutrophic lake on Aland, South west Finland. Water Research 23(4):481-486. |
|
|
Los DA, Mironov KS (2015). Modes of fatty acid desaturation in cyanobacteria: an update. Life 5(1):554-567. |
|
|
Loudiki M (1990). Etude limnologique d'un hydrosystème récemment aménagé dans la région de Marrakech (Maroc): apports du bassin versant, hydrochimie, algues et macrophytes (Doctoral dissertation, Aix-Marseille 3) 353 p. |
|
|
Loudiki M, Bouterfas R, Mouhri K (1994). Dynamique et évaluation des apports du bassin versant et de l'état trophique du lac-réservoir Hassan I (Maroc). Revue. Faculté. Sciences. Marrakech 8:45-57. |
|
|
Loudiki M, Oudra B, Sabourd B, Sbiyyaa B, Vasconcelos V, Franca S (2002). Taxonomy and geographic distribution of potentiel toxic cyanobacterial strains in Morocco. Annales de Limnologie 38(2):101-108. |
|
|
Lund HC, Lund JWG (1995). Freshwater algae: their microscopic world explored. Hong Kong: Dah Hua Ltd. ISBN 0-948737-25-5. |
|
|
Mahapatra DM, Ramachandra TV (2013). Algal biofuel: bountiful lipid from Chlorococcum sp. proliferating in municipal wastewater. Current Science 105:47-55. |
|
|
Major Y, Kifle D, Spoof L, Meriluoto J (2018). Cyanobacteria and microcystins in Koka reservoir (Ethiopia). Environmental Science and Pollution Research 25(27): 26861-26873. |
|
|
Malki M (1994). Etude de la communauté phytoplanctonique et des caractéristiques physico-chimiques des eaux du lac reservoir Al Massira. Thèse de Doctorat d'état, Université Hassan II, Casablanca 287p. |
|
|
Miles CO, Sandvik M, Nonga HE, Rundberget T, Wilkins AL, Rise F, Ballot A (2013). Identification of microcystins in a Lake Victoria cyanobacterial bloom using LC-MS with thiol derivatization. Toxicon 70:21-31. |
|
|
Moreira C, Gomes C, Vasconcelos V, Antunes A (2020). Cyanotoxins occurrence in Portugal: A new report on their recent multiplication. Toxins 12(3):154. |
|
|
Murata N, Wada H, Gombos Z (1992). Modes of fatty-acid desaturation in cyanobacteria. Plant Cell Physiology 33:933-941. |
|
|
Nagase H, Inthorn D, Oda A, Nishimura J, Kajiwara Y, Park MO, Miyamoto k (2005). Improvement of selective removal of heavy metals in cyanobacteria by NaOH treatment. Journal of Bioscience and Bioengineering 99:372-377. |
|
|
Namikoshi M, Rinehart KL, Sakai R, Stotts RR, Dahlem AM, Beasley VR, Carmichael WW, Evans WR (1992). Identification of 12 hepatotoxins from a Homer Lake bloom of the cyanobacteria Microcystis aeruginosa, Microcystis viridis, and Microcystis wesenbergii: nine new microcystins. The Journal of organic chemistry 57(3):866-872. |
|
|
Namikoshi M, Sun F, Choi BW, Rinehart KL, Carmichael WW, Evans WR, Beasley VR (1995). Seven more microcystins from homer lake cells: Application of the general method for structure assignment of peptides containing. Alpha. Beta-dehydroamino acid unit (s). The Journal of Organic Chemistry 60(12):3671-3679. |
|
|
Oudra B, Loudiki M, Sabour B, Sbiyyaa B, Vasconcelos V (2001b). Étude des blooms toxiques à cyanobactéries dans trois lacs réservoirs du Maroc: Résultats préliminaires. Revue des sciences de l'eau 15:301-313. |
|
|
Oudra B, Loudiki M, Sbiyyaa B, Martins R, Vasconcelos V, Namikoshi N (2001a). Isolation, characterisation and quantification of microcystins (heptapeptides hepatotoxins) in Microcystis aeruginosa dominated bloom of Lalla Takerkoust lake-reservoir (Morocco). Toxicon 39:1375-1381. |
|
|
Oudra B, Loudiki M, Sbiyyaa B, Vasconcelos V, Zerrouk H, El-Andaloussi-Dadi M, Darley J (1998). Occurrence of hepatotoxic Microcystis aeruginosa blooms in eutrophic Moroccan lake-reservoir. Harmful Algae, Xunta de Galicia and Intergovernamental Oceanographic Comisión (IOC) of UNESCO pp. 29-31. |
|
|
Oudra B, Loudiki M, Vasconcelos V, Sabour B, Sbiyyaa B, Oufdou K, Mezrioul N (2002a). Detection and quantification of microcystins from cyanobacteria strains isolated from reservoirs and ponds in Morocco. Environmental Toxicology 17(1):32-39. |
|
|
Ouhamdouch S, Bahir M, Ouazar D, Carreira P M, Zouari K (2019). Evaluation of climate change impact on groundwater from semi-arid environment (Essaouira Basin, Morocco) using integrated approaches. Environmental Earth Sciences 78(15):449. |
|
|
Ouhsassi M, Khay EL, Bouyahya A, Abrini J (2017). Isolation and characterization of cyanobacteria strains based on the compositional approach of fatty acids: case of drinking water reservoirs in the region of Tetouan (Northern Morocco). Journal of Algal Biomass Utilization 9(1):57-67. |
|
|
Passaquet C, Zabulon G, Lemoine Y, Dubacq JP, Thomas JC (1989). Lipides et pigments des fractions sub-membranaires obtenues par traitement à la digitonine chez la cyanobactérie Pseudanabaena sp. Souche M2. Bulletin de La Société Botanique de France. Actualités Botaniques 136(1):169-173. |
|
|
Paerl HW (2016). Impacts of climate change on cyanobacteria in aquatic environments. Climate change and microbial ecology: Current research and future trends pp. 5-22. |
|
|
Perrin JL, Raïs N, Chahinian N, Moulin P, Ijjaali M (2014). Water quality assessment of highly polluted rivers in a semi-arid Mediterranean zone Oued Fez and Sebou River (Morocco). Journal of Hydrology 14(510):26-34. |
|
|
Plaas HE, Paerl HW (2021). Toxic cyanobacteria: a growing threat to water and air quality. Environmental Science and Technology 55(1):44-64. |
|
|
Poon GK, Griggs LJ, Edwards C, Beattie KA, Codd GA (1993). Liquid chromatography-electrospray ionization-mass spectrometry of cyanobacterial toxins. Journal of Chromatography A 628(2):215-233. |
|
|
Quesada A, Sanchis D, Carrasco D (2004).Cyanobacteria in Spanish reservoirs. How frequently are they toxic? |
|
|
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY (1979). Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 111(1):1-61. |
|
|
Rouhiainen L, Sivonen K, Buikema WJ, Haselkorn R (1995). Characterization of toxin-producing cyanobacteria by using an oligonucleotide probe containing a tandemly repeated heptamer. Journal of Bacteriology 177(20):6021-6026. |
|
|
Rose E, Sabir M, Laouina A (2010). Gestion durable des eaux et des sols au Maroc: valorisation des techniques traditionnelles méditerranéennes. IRD Editions Marseille. |
|
|
Sabour B (2002). Etude des blooms d'algues toxiques à Cyanobactéries et Haptophycées dans le lac de barrage Oued Mellah (Maroc): ecologie, écophysiologie et toxicologie. PhD thesis, Mohamedia, Morocco. |
|
|
Saoudi A (2008). Isolement, culture et evaluation de la toxicite des afflorescences à microcystis sp; du barrage mexa (el-tarf) (doctoral dissertation, université de annaba-badji mokhtar). |
|
|
Saoudi A, Brient L, Boucetta S, Ouzrout R, Bormans M, Bensouilah M (2017). Management of toxic cyanobacteria for drinking water production of Ain Zada Dam. Environmental Monitoring and Assessment 189(7):1-11. |
|
|
Sarsekeyeva FK, Usserbaeva AA, Zayadan BK, Mironov KS, Sidorov RA, Kozlova AY, Los DA (2014). Isolation and characterization of a new cyanobacterial strain with a unique fatty acid composition. Advances in Microbiology 4(15):1033. |
|
|
Sas H (1989). Lake restoration by reduction of nutrient loading: expectations, experiences, extrapolations. Academia Verlag Richarz, St. Augustin, Germany. |
|
|
Sbiyyaa B (1997). Déterminisme, écophysiologie, toxicité et implications écologiques du bloom toxique à Microcystis aeruginosa Kütz. (Cyanobactérie) dans le lac de barrage Lalla Takerkoust (Maroc). Thèse. University. Cadi Ayyad, Marrakech 219 p. |
|
|
Sharathchandra K, Rajashekhar M (2011). Total lipid and fatty acid composition in some freshwater cyanobacteria. Journal of Algal Biomass Utilization 2(2):83-97. |
|
|
Shimizu K, Sano T, Kubota R, Kobayashi N, Tahara M, Obama T, Sugimoto N, Nishimura T, Ikarashi Y (2014). Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes Toxins 6:168-179. |
|
|
Thajuddin N, Subramanian G (2005). Cyanobacterial biodiversity and potential applications in biotechnology. Current Science 89(1):47-57. |
|
|
Vasconcelos VM, Sivonen K, Evans WR, Carmichael WW, Namikoshi M (1996). Hepatotoxic microcystin diversity in cyanobacterial blooms collected in Portuguese freshwaters. Water Research 30(10):2377-84. |
|
|
Wiegand C, Pflugmacher S (2005). Ecotoxicological effects of selected cyanobacterial secondary metabolites a short review. Toxicology and applied pharmacology 203(3):201-218. |
|
|
Yoon JH, Shin JH, Kim MS, Sim SJ, Park TH (2006). Evaluation of conversion efficiency of light to hydrogen energy by Anabaena variabilis. International Journal of Hydrogen Energy 31:721-727. |
|
|
Yu SZ (1995). Primary prevention of hepatocellular carcinoma. Journal of gastroenterology and Hepatology 10(6):674-682. |
|
|
Zastepa A, Watson SB, Kling H, Kotak B (2017). Spatial and temporal patterns in microcystin toxins in Lake of the Woods surface waters. Lake and Reservoir Management 33(4):433-443. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


