ABSTRACT
Among the crops that can potentially be used to provide the raw material needed for liquid biofuel production Jatropha curcas L. is the one that generates most interest in many development projects. The aim of this study is to evaluate the production and development of 3-year-old J. curcas L. seedlings in 2012. The results showed that after three years of planting, subsoiling and provenance have no effect on growth variables and on the major part of foliar functional traits. However, the number of branches in bloom (p = 0.0041) and in fruiting (p = 0.0091) varied significantly under the effect of subsoiling. Branching is significantly higher in the prepared plot than in the control. As a conclusion, tillage has increased branching, while provenance has improved growth in height of J. curcas L. plants.
Key words: Tillage, provenance, branching, growth.
Among the crops that can potentially be used to provide the raw material needed for liquid biofuel production, Jatropha curcas L. is the one that is of most interest to various development organizations in the tropical and subtropical areas. These biofuels are extracts of renewable sources that bring about environmental benefits, and make it possible to create jobs, both in the collection phase and in the treatment.
J. curcas is a shrub, of the Euphorbiaceae family, originating from Central America (Nicolas, 2010). It exists in all the arid and semi-arid tropical regions of the world (Henning, 2002). In Africa, it can be found in the Sudanese and Guinean savannas and extends from Senegal to Cameroon (Assogbadjo et al., 2010).
According to its most convinced promoters, J. curcas can produce large quantities of high quality inedible fuel oil, restore marginal soils, improve the fertility of all types of soil, reforest degraded lands, promote land safety, limit cattle damages on crops, diversify farmers' income and obtain by-products with phytosanitary and therapeutic uses (Mergeai, 2011).
The exploitation of Jatropha is recent and needs to be established over the long term. Therefore, our current knowledge and studies must be deepened. Jatropha and its by-products are promising, yet technical and agronomic knowledge must be strengthened.
A study carried out two years after having planted in Bargny (Senegal) showed that: subsoiling seems to improve the growth of the upright part (neck, crown) of Jatropha (Barro, 2010). Similarly, growth variables, tree architecture, and fruit numbers also vary significantly between Jatropha provenances (Barro, 2010).
This present study is based on studies (Barro, 2010) in order to further them. It aims at evaluating the production (fruit and leaves) and development (growth variables and architecture of air ports) of J. curcas (JCL) plants aged 4 years in 2012. Two hypotheses were tested: (i) provenance and tillage have affected the growth and productivity of J. curcas plants; (ii) some functional features (leaf thickness, leaf stretch, leaf area, specific leaf area (SLA) and leaf dry matter content (LDMC)) of J. curcas vary according to provenance and soil work.
The plant material of J. curcas comes from the industrial plantation of SOCOCIM located in Bargny. The plants are located next to the limestone quarries and consist of two provenances: "Diobass" and "Casamance". The studied plants were 4 years old. They were first studied for the first time 2 years after being planted by Barro (2010) in the same site.
The small tools used to carry out this study consisted of an electronic vernier caliper to determine the thickness of the leaves, a nesting perch measuring the height of the JCL plants, plastic bags for the conservation of samples taken, a pruning shears pruner and a tape measure for measuring lengths (branches), circumferences (collar) and diameters (crowns).
Experimental plan
The planting was carried out in 2008 at the SOCOCIM site located in Bargny, with the provenances of JCL "Diobass" and "Casamance". It was fulfilled in rows at a density of 625 plants/ha (4 m × 4 m). A crossed subsoiling of 1 tooth (dent semat) was made to a depth of 40 cm on a part of the plot before planting while the rest of the plot was not subject to any land preparation.
The plants of J. curcas were irrigated for 12 months after being planted, at the rate of two waterings of 16 L/plant/week. The main plot provenance factor and the soil tillage factor in secondary plots are the only factors studied in this experiment, the experimental device of which was a split-block with three repetitions, the second factor (tillage) being randomized at the block scale and not within each main plot.
The factors studied had each two levels: "provenances" (Diobass and Casamance) and "tillage" (subsoil and control). Three gross plots of 36 m, that is, 1,296 m2, containing 100 J. curcas, were demarcated. Within each of these plots, a batch of 10 randomly selected and labeled plants constituted the experimental unit. The combination of the different levels of the factors studied gave a total of 4 treatments. Each treatment was repeated 3 times, that means 12 experimental units each one containing 10 JCL plants.
Measured traits
At the scale of the experimental unit (lot of 10 plants), the following variables were determined: the number of low primary branches (0 - 15 cm of the soil), the number of high primary branches (height greater than 15 cm), the number of secondary branches, the number of flowering branches, the number of fruiting branches, the total height (length of the longest strand), the diameter at the neck of the main stem, the diameter of the crown in the direction parallel to the planting line and following the direction perpendicular to the planting line, the state of maturity of the fruit (number of green fruits, number of yellow fruit, number of black fruits or chestnuts, number of total fruits) taking into account the order of branches. At the gross plot scale, survival rates were evaluated.
Measurements also focused on the following functional features: i) leaf thickness, ii) leaf surface, iii) leaf surface area (SLA), iiii) leaf stretching.
According to Violle et al. (2007), a "functional trait" is defined as any morphological, physiological or phenological characteristic, measurable at the individual level without reference to the environment and intervening in the adaptive or fitness value of the species.
To achieve this, three plants from each source were randomly selected, in a well-lit environment, in the prepared plot and in the control. A well-lit branch with a diameter of about 1 cm and at least 1 m long per plant was selected. Six intact fresh leaves of the branch were chosen for the measurements. The selected leaves were kept cool in aluminium foil, placed in a box in the dark, and used throughout the next 24 h.
At first, the leaves were tagged and weighed with a scale to determine their fresh weight (g). Subsequent to these measurements, they were then placed in small aluminium foil bags and dried for at least 48 h at 72°C (up to constant weight) in order to obtain the dry mass of the leaf. It consists of an approximation of the leaf tissue density (when the leaf is not spongy), and is positively correlated with the retention of nutrients in the leaf. By this definition, the leaf dry-matter content (LDMC) was determined by the ratio between the mass of a dry leaf and its weight in the fresh state (g g-1):
LDMC (Leaf dry-matter content) = (Dry mass) / (Fresh mass) (1)
Then for the thickness of the leaf (mm), the measurements were made at 6 points for each leaf in its fresh state, using a digital caliper. The main veins and the midrib have been avoided. Then, each leaf was scanned to obtain a digital image. The leaves have been flattened as much as possible so that the leaf covers the largest possible area, but not crushed to avoid tissue damage.
The cross section of the leaves was supposed to be circular, and an adjustment was made accordingly to calculate the average thickness (Em) by multiplying the diameter by π / 4.
Em = (ΣEi) / 6 x π / 4 (2)
where E represents the thickness of the leaf; i ranging from 1 to 6 leaves.
In the same process, the area of ​​each projected leaf was determined using the Midbmp software. The Foliar Specific Surface (SLA) was obtained by the relation between the surface of a leaf in m2 and its mass in kg.
SLA (Specific leaf area) = (Specific surface area of ​​the leaf) / (Mass of leaf) (3)
Each scanned image (the entire leaf) had a corresponding weight allowing us to calculate the Specific leaf area (SLA). SLA of the branch leaves (expressed in m2.kg-1) was calculated by averaging the specific areas of the six leaves taken from that branch.
(SLA) mean = (ΣSLA) / 6 (4)
For the stretching of the leaves, three plants in each treatment were targeted. Six leaf buds from one branch per plant were selected and labeled and followed until one month of age. The stretching of one-month-old leaves was determined, followed by that of the two-month-old leaves.
The measurements of lengths were carried out with a tape measure in meters except for the height which was measured using a nesting perch of measurement (8 m). Mass measurements were obtained with a precision balance (1/10 000).
Statistical analysis
All the collected data were entered into ExcelTM and analyzed for variance (ANOVA) followed by a mean comparison test (Tukey test) to identify significantly different treatments. The analysis was performed with Statistix 10 software.
Effect of tillage and provenance on growth and survival of J. curcas L.
The analysis of the results showed considerable differences (Table 1) in the total height (p = 0.0202) and in the survival rate (p = 0.0095) of JCL plants under the effect of the interaction between the factors "provenance" and "subsoiling" (Interaction Prov α SS).
The Tukey test enabled us to identify two homogeneous groups: group A composed of Casamance with an average of 1.90 ± 0.01 m and group B composed of Diobass provenance with an average of 2.10 ± 0.01 m (Figure 1). The Tukey test (Table 1) revealed two significantly different groups: group A composed of Casamance provenance with subsoiling (Casa SS) treatment with an average height of 2.07 ± 0.07 m and the group B composed of the treatment "Casamance without subsoiling (control)", noted CasaTem with 1.91 ± 0.07 m. In the "source of cultivation and subsoiling" interaction, the "source Casamance × subsoiling" treatment recorded, on the total height, significantly higher values ​​compared to the "source Casamance × control without subsoiling" treatment.
The same observation was made on the Diobass provenance with two distinct groups revealed by the Tukey test: group A composed of the plot (Diob SS) worked (subsoiling) with an average height of 2.02 ± 0.03 m and group B composed of the unworked plot (DiobTem) (control) with 2.23 ± 0.04 m. The same trend was noted for the survival rate of 98 and 87%, respectively, in the plots worked from Casamance (Casa SS) and Diobass (Diob SS) provenances. The survival rate in the control (non-tillage) plots of Casamance and Diobass provenances was 74.3 and 94%, respectively.
In the "provenance and tillage" interaction, the treatment "Provenance Casamance × subsoiling" recorded, on the survival rate, significantly higher values ​​compared to the treatment "source Casamance × control without subsoiling". The reverse has been registered with the Diobass provenance. Subsoiling was only favorable for the Casamance provenance.
The main factors “provenance and tillage” and their interactions did not affect the crown diameter of the main stem of JCL plants, crown diameter, and survival rate. These variables did not vary significantly under their influence (Table 1).
Due to provenance, Diobass recorded a diameter at the neck of the main stem (11.5 ± 6.2 cm) slightly greater than that of Casamance (11.2 ± 6.2 cm). The same trend is observed for the height, the crown diameter and the survival rate. Similarly, the interaction of factors provenance and subsoiling (Table 1) showed that plots with a diameter at the neck of the stem of the higher JCL plants had higher survival rates, crown diameters and heights. However the plots with small neck diameters of JCL plants have recorded low survival rates, crown height and crown diameter. These results show that the larger the neck diameter of the main stem, the more the plants have better growth and survival rate.
Effect of tillage and provenance on the branching of J. curcas L.
The results showed that the number of high primary branches (NR1h>15 cm from soil) with p = 0.0408, the total number of primary branches (NR1) with p = 0.0381 and the number of primary branches in leaf (NF1) with p = 0.0381 significantly varied under the effect of subsoiling (Table 2). The Tukey test revealed two distinct homogeneous groups: the group A composed of the worked plot (subsoiling) and the group B composed of the control (not worked) respectively produced 8.63 ± 1 and 9.98 ± 1, 0 for NR1h; 11.28 ± 0.60 and 12.47 ± 0.60 for NR1 and 11.28 ± 0.60 and 12.47 ± 0.60 for NF1 (Figure 2).

The source and the interaction between the two factors (source and subsoil) did not have a significant effect on the number of low primary branches (NR1b <15 cm of the soil), the number of high primary branches (>15 cm soil = NR1h), the total number of primary branches (NR1), the number of secondary branches (NR2), the number of primary branches in leaf (NF1) and the number of secondary branches (NF2) in leafing (Table 2).
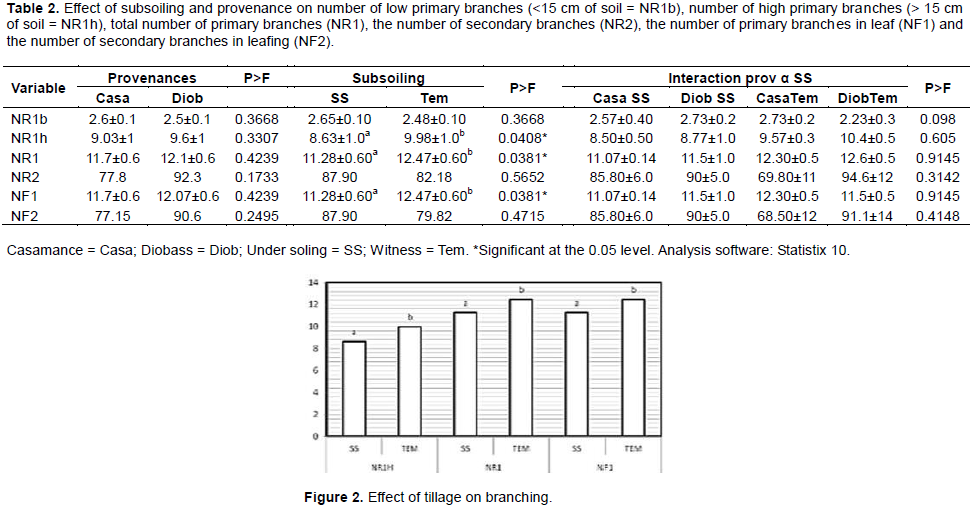
Branching was significantly better for Diobass than for Casamance (Table 2). The total number of primary branches (NR1) and the number of secondary branches (NR2) were 12.1 ± 0.6 and 92.3, respectively on Diobass whereas Casamance provenance had 11.7 ± 0.6 and 77.8, respectively in the same order. Subsoiling had a depressive effect on primary branching (Table 2), the control of which recorded the highest number (12.47 ± 0.6) compared to the worked plot (11.28 ± 0.60). Although subsoiling did not affect the number of secondary branches (NR2), the prepared plot had the highest number.
The interaction of source and subsoiling factors has shown that tillage has a depressive effect on primary and secondary branching for the Diobass. While for the Casamance provenance, soil preparation improved the number of primary branches and was negative for the number of secondary branches. We have the same trend for the number of branches in leaf from each source.
Productivity of J. curcas L. plants as a result of soil preparation and provenance
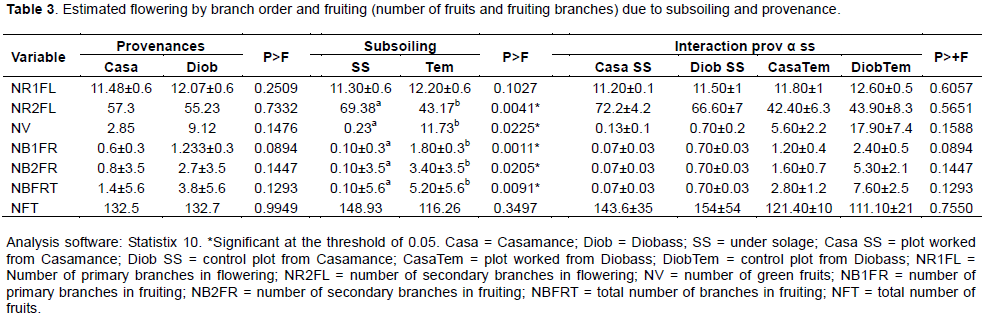
The number of secondary branches in flowering (NR2FL, p = 0.0041), the number of green fruit (NV, p = 0.0225), the number of primary fruiting branches (NB1fr, p = 0.0011), the number of secondary fruiting branches (NB2fr, p = 0.0205), and the total number of fruiting branches (NBfrt, p = 0.0091) significantly varied under the effect of subsoil (Table 3). The Tukey test enabled us to identify two distinct groups under the effect of subsoiling: group A composed of the worked plot and group B represented by the control recorded as 69.38 and 43.17, respectively for NR2FL"; 0.233 and 11.73 for "NV"; 0.1 and 1.8 ± 0.3 for "NB1FR"; 0.1 and 3.4 for "NB2FR"; 0.1 and 5.2 for "NBfrt" (Table 3).
However, in addition to the variables mentioned earlier (Table 3), the source and the interaction of the two factors (source and subsoil) did not have a significant effect on all the supervised variables.
The number of secondary productive branches (number of secondary branches in flowering and fruiting) was much higher than the number of primary productive branches (number of primary branches in fruiting and flowering) for the Casamance provenance (respectively 58.1 and 18, 08) as for Diobass (respectively 57.93 and 13.3). During the period of data collecting, the fruits were all green; that is why there are no results on the number of yellow and black fruits.
Effect of tillage and provenance on some functional traits of J. curcas L.
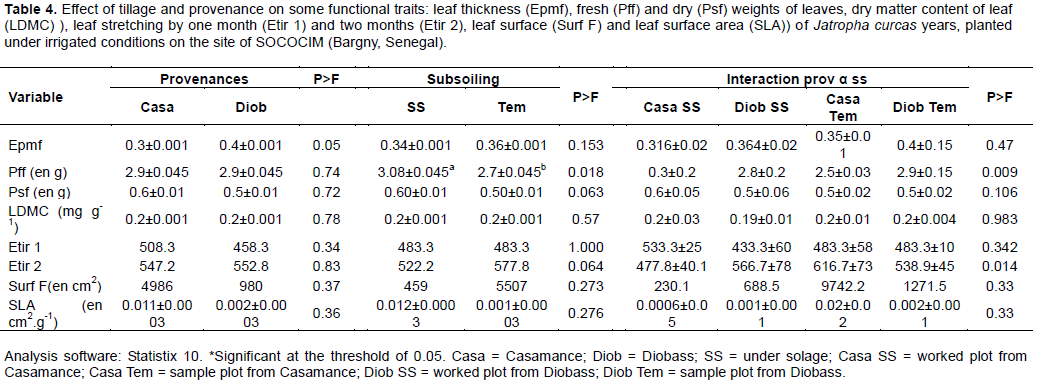
After planting, the measures taken on the average leaf thickness (Epmf) taken from the 4-year-old J. curcas varied significantly (Casamance: 0.334 mm, Diobass: 0.371 mm) depending on the source (p = 0.05) (Table 4). On the contrary, subsoiling and factors interaction did not have a significant effect on this variable (Epmf).
However, leaf fresh weight (p = 0.009) (Table 4) and two-month-old leaf stretch (p = 0.014) (Table 4) significantly varied due to the interaction between the two factors (source and subsoiling).
The diameter at the neck of the main stem and the diameter of the crown did not significantly vary under the effect of tillage and provenance factors after 3 years of planting. On the other hand, the height significantly varied under the effect of the interaction of the two factors; although Casa Tem plots gave the lowest heights. The total height of 3-year-old J. curcas was 1.90 ± 0.01 m for the Casamance provenance and 2.12 ± 0.01 for Diobass. It reached 2.04 m in the worked plot and 2.07 m in the control plot. These results are substantially similar to other studies (Chehaibi et al., 2008), where evidence is presented that tillage has a favorable effect on the plants’ development. It improves in particular the speed of growth.
This same plantation was studied by Barro (2010) with a total height of 1.8 ± 0.004 m for the provenance Casamance, 1.5 ± 0.004 m for Diobass, 1.7 m in the worked plot and in the control. The new results show a clear increase in height growth of J. curcas plants. Huwe (2003) noted that tillage influences the biotic and abiotic processes, including the modification of soil structural properties such as cracks, aggregates and pore continuity, as well as soil aeration, temperature levels and humidity, what should promote the growth of plants. In this study, tillage has improved height growth for the Casamance provenance and was depressive for Diobass. Similarly, the preparation of the soil has positively influenced the survival rate for the Casamance provenance and has negatively impacted on the Diobass provenance. Overall, the worked plot had the highest survival rate. The present results are better than those of Kobilke (1989) in Cape Verde and Heller (1992) in Senegal who reported 5 to 50% survival rates with direct seeding.
Tillage (subsoiling) has an important outcome on the number of primary branches, the number of high primary branches, the number of primary branches in leafing, the number of secondary branches in bloom and the number of branches in fruiting. These results corroborate with the observations of Domergue and Pirot (2008) who noted that the preparation of the soil leads to a faster tree development of the plant, with a broad crown. Similarly, Kouyaté et al. (2008) reported that tillage directly influences crop development. In the same vein, Eléonore et al. (2012) report that the more soil is worked in depth with a high frequency, the greater the amount of mineralized organic matter that can lead to a good development of Jatropha plants.
However, tillage does not have a significant effect on the low primary branching and secondary branching. Nevertheless, the prepared plots had the highest number of branches. The results have shown an increase in the growth and branching of J. curcas under the effect of provenance and tillage between 2 (Barro, 2010) and 3 years (Barro, 2013) at the SOCOCIM site. The number of branches correlates well with production, indicating that an early branching of JCL would have a decisive effect on the yield because the inflorescences only develop at the end of branches (Domergue and Pirot, 2008; Legendre, 2008). So the more branches the tree has, the more fruit it produces, thus increasing the yield in this SOCOCIM plantation.
The choice of more productive provenance is a very delicate parameter, since production can vary from one year to the next and is even dependent on several factors. Comparative tests of 13 varieties made in Keur Samba Gueye and Toubacouta (Heller, 1992) have shown that the best results were obtained with the local variety (collected at Santhie Ram).
Legendre (2008) reveals that the results obtained at Beude Dieng by the Italian-Senegalese SBE Senegal project tend to indicate that, out of a dozen origins from all the continents, the plants coming from the Sokone zone are the most homogeneous and present the best vegetative development. Similarly, seed size (length, width and thickness), weight (100 seeds, hull and kernel), and germination rate of two provenances (Kaffrine and Nioro) were followed by Ly et al. (2015).
Growth in height, neck diameter, and number of leaves were measured after 60 days. The results obtained showed that the Kaffrine origin had significantly heavier seeds (weight 100 seeds = 71.50 ± 1.22 g, weight = 14.50 ± 0.41 g and weight almond = 22.75 ± 0, 50 g), longer (18.82 ± 0.78 mm) and wider (11.41 ± 0.42 mm) with germination rates (90%) higher than those of Nioro (Ly et al., 2015).
Most of the functional traits (Epmf, Psf, LDMC, Etir1, Etir2, Surf f, SLA) did not vary between provenances or even under the effect of subsoiling. A dense, thick-leafed leaf has a strong LDMC, which is not the case for a leaf rich in photosynthetic tissues. A high LDMC is often associated with a long leaf life because it gives better mechanical resistance to damage caused by drought and friction (wind) (Bumb, 2012).
However, the dry substance content (LDMC) and specific leaf area values ​​obtained in this study are very low and the JCL plants showed more or less slow growth. These results confirm the previous studies (Westoby, 1998) showing that high SLA values ​​characterize fast-growing plants with large leaves but low longevity; whereas high LDMC values ​​characterize plants leaf organs of which accumulate nutrients and constitute important storage pools with low tissue turnover.
According to Westoby et al. (2002), high ALS species have strategies for rapid leaf material production and frequent leaf turnover. They are efficient in acquiring resources, but poor in the conservation of nutrients in their tissues. These assertions show that J. curcas plants do not grow rapidly and have a low frequency for leaf renewal at the Bargny site.
The purpose of this study was to evaluate the effect of tillage and provenance on the growth and the productivity of J. curcas plants. The study has revealed that subsoiling and provenance did not affect growth variables and most foliar traits. However, the number of blooming and fruiting branches varied significantly under the effect of subsoiling. Branching is significantly higher in the prepared plot than in the control plot. Likewise, the survival rate is better in the worked field compared to the control.
The interaction of provenance and subsoiling factors had an effect on height growth and survival rate of JCL plants. On the other hand, it has no effect on the branching of JCL plants.
The production of a plant of J. curcas L. would, therefore, depend on the branching. Based on these current findings, additional questions arise: which method to apply to Jatropha plants to increase the number of branches? Is the size of the branches a good strategy to stimulate branching? These questions have justified the next studies to be pursued in our research.
The authors have not declared any conflict of interests.
REFERENCES
|
Assogbadjo AE, Sinsin B (2010). Current state of plant diversity in Benin. In Atlas of the Biodiversity of West Africa (Volume 1), Sinsin B, Kanpman D (eds). pp. 222-227.
|
|
|
|
Barro L (2010). Effect of tillage on the agronomic performance of Jatropha curcas in Bargny. Master's thesis, Agroforestry, Ecology, Adaptation. University Cheikh Anta Diop of Dakar; Department of Plant Biology.
|
|
|
|
|
Barro L, Samba NAS, Diatta M, Akpo EL (2013). Effect of tillage on the productivity of different provenances of Jatropha curcas. OCL 20(3):165-170.
Crossref
|
|
|
|
|
Bumb I (2012). Effects of the functional composition of subalpine grassland communities on primary production and forage quality. Montpellier II University, Place Eugene Battalion, CC 065, 34095 MONTPELLIER Cedex 05.
|
|
|
|
|
Chehaibi S, Hannachi C, Pieters JG, Verschoore RA (2008). Keywords, Effects of plowing tools on soil structural condition and yield of a potato crop Re-plowing- Resistance penetration- Voluminous mass- Plant biomass- Tubers yield-Tunisia.
|
|
|
|
|
Domergue M, Pirot R (2008). Jatropha curcas L, bibliographic synthesis report. CIRAD, Agropolis Avenue 34398 Montpellier Cedex 5 AGROgeneration, 45-47 rue de Monceau 75008 Paris.
|
|
|
|
|
Eléonore B, Gérard G, Didier J, Blaise L, Yves N (2012). Adapting Organic Inputs on the Ground.
|
|
|
|
|
Heller J (1992). Untersuchugen über Gentopische Eigenschaften und Vermehrungsung und An-bauVerfähren bei des Pürgiernuss (Jatropha curcas L.). 1992: Hamburg.
|
|
|
|
|
Henning RK (2002). Jatropha curcas in Africa, The Global Facilitation Unit for Underutilized Species (GFUUS), Weissensberg, Germany, P 49.
|
|
|
|
|
Huwe B (2003). The role of soil for soil structure. In: El Titi A (ed) Soil tillage in agroecosys-tems. CRC, Boca Raton, Florida pp. 27-49.
Crossref
|
|
|
|
|
Kobilke H (1989). Untersuchungen zur Bestandesbegrundung von Purgiernuß (Jatropha curcas L.). in University. 1989, Hohenheim: Stuttgart.
|
|
|
|
|
Kouyaté AM, Van Damme P, Goyens S, De Neve S, Hofman G (2008). Evaluation of soil fertility at Detarium microcarpum Guill. & Perr. Detarium microcarpum- Soil fertility- Physicochem-ical properties- C / N- Mali.
|
|
|
|
|
Legendre B (2008). Jatropha curcas (Tabanani), Agronomic Note. Performance Company RC 96B1982 - Dakar Ninea 190263 2B2.
|
|
|
|
|
Ly MO, Kumar D, Diouf M, Nautiyal S, Diop T (2015). Morphological traits of seed and vigor of seedlings of two provenances of Jatropha curcas L. in Senegal. Journal of Applied Biosciences 88:8249-8255. ISSN 1997-5902.
Crossref
|
|
|
|
|
Mergeai Guy (2011). Jatropha curcas L: miraculous plant or devil tree? Tropicultura 29:1-2.
|
|
|
|
|
Nicolas C (2010). Jatropha curcas L: a review. Progress of Botanical Research 2(50): 39-86.
Crossref
|
|
|
|
|
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I & Garnier E (2007). Let the concept of be functional, Oikos 116(5):882-892.
Crossref
|
|
|
|
|
Westoby M (1998). A leaf height seed (LHS), Plant ecology strategy scheme. Plant Soil 199:213-227.
Crossref
|
|
|
|
|
Westoby M, Falster DS, Moles AT, Vesk PA & Wright IJ (2002). Plant ecological strategies: Some leading dimensions of variation between species. Annual Review of Ecology, Evolution, and Systematics 33:125-159.
Crossref
|
|