Full Length Research Paper

ABSTRACT
INTRODUCTION
Water Hyacinth (Eichhornia crassipes) can be described as a floating macrophyte with thick, glossy, round leaves, inflated leaf stems, and lavender flowers (Toft et al., 2003). It is a free-floating aquatic macrophyte that clusters in large mats that block shorelines (Rocha-Ramirez et al., 2007). It is sometimes found stuck in mud, appearing rooted and it is rarely found as a single plant (Villamagna, 2009). The foliage helps to provide shade and the roots provide filtration, a spawning area and refuge for the aquatic organisms like macroinvertebrates and fish (Schramm et al., 1987). The macrophyte absorbs excess nutrients from the water, helping to reduce algal blooms thus an excellent water clarifier (Rocha-Ramirez et al., 2007). Being a floating macrophyte it moves about with the filtrates thus expected to present a similar microhabitat condition wherever it goes. Studies done in other parts of the world have shown that specific macroinvertebrate taxa inhabit water hyacinth regardless of the region (de Marco et al., 2001; Toft et al., 2003; and Rocha-Ramirez et al., 2007). This study thus set out to confirm this and identify which taxa inhabit water hyacinth in the Kenyan part of Lake Victoria.
MATERIALS AND METHODS
Study Area
The Kenyan portion of Lake Victoria can conveniently be divided into Nyanza Gulf and open waters. The Nyanza Gulf receives water from several rivers and is eutrophic due to nutrients from these rivers while the expansive open waters have less nutrients. A total of 16 stations were sampled during the study between 18 and 26th December, 2013; 8 stations were in the Nyanza gulf while 8 were in the open waters of Lake Victoria (Figure 1). All the stations were geo-referenced using Magellan global positioning system. The study was carried out in the early stages of water hyacinth reinfestation when most mats were found at the shallow littoral areas of the lake.
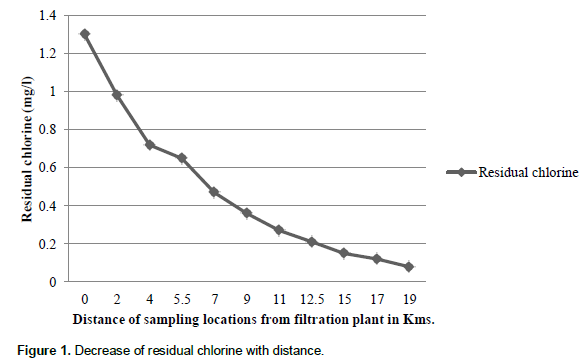
Sampling
Physico-chemical Parameters
Selected physic-chemical parameters were taken in-situ using appropriate meters. The parameters measured include water depth, Dissolved Oxygen (DO), turbidity, conductivity, pH, redox and temperature. At each station, these parameters were taken in triplicates and later averaged.
Macroinvertebrates
Sampling of macroinvertebrates was done in sixteen stations using a Ponar grab and a 500 µm mesh size scoop net. In each sampling station, three grabs and three scoops of macroinvertebrate samples were collected and washed at the site of collection sorted live and then preserved in absolute ethanol. In the laboratory they were identified to genus level. The vertical position of the macroinvertebrates was further examined to show which taxa were found at the benthos below the mat, at the roots and within the stem. This was done by taking samples at the benthos below the mat, examining the sediments trapped on the roots and opening the stems to see if there are any organisms within.
Data analysis
The data on physic-chemical parameters were summarized as mean ± SE and represented in a table. Variation between the stations was tested using one-way ANOVA. The identified benthic macroinvertebrates were then analyzed for taxon diversity (Shannon-Wiener, HI), abundance, evenness, and relative abundance for the lake at each sampling station. The data was represented in frequency tables and bar graphs. Log transformed one way analysis of variance was done to test if there is any variation between the stations sampled.
RESULTS AND DISCUSSION
Physico-chemical parameters
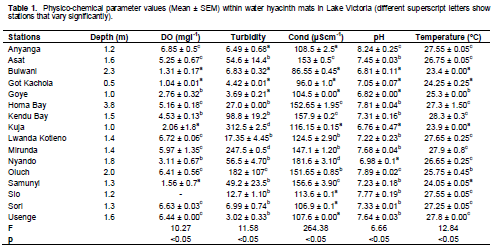
The values of the physic-chemical parameters are shown in Table 1. The DO values ranged between 1.04 ± 0.01mgL-1 and 6.85 ± 0.5 mgL-1 while turbidity ranged between 3.69 ± 0.21 and 312.5 ± 2.5. Conductivity hardly went below 100 µScm-1 except Bulwani and Got Kachola with 86.55 ± 0.45 and 96.0 ± 1.0, respectively while Ph ranged between weak acidic and weakly basic values. Temperature of these stations ranged between 23.4 ± 0.00 and 27.8 ± 0.00. Statistical tests revealed significant variations between the stations for all the parameters but on pairwise comparison, it was discovered that majority of the stations were at one sub set for all the parameters (Table 1). This result shows that there are similarities between the stations.
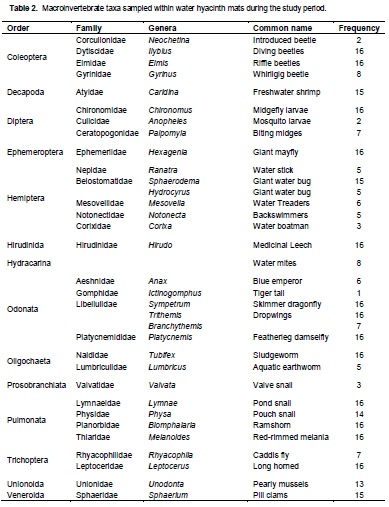
A total of 14 orders, 30 families and 34 genera were found during the sampling period (Table 2). Thirteen of these genera were found at all sampling stations as shown in Table 2.
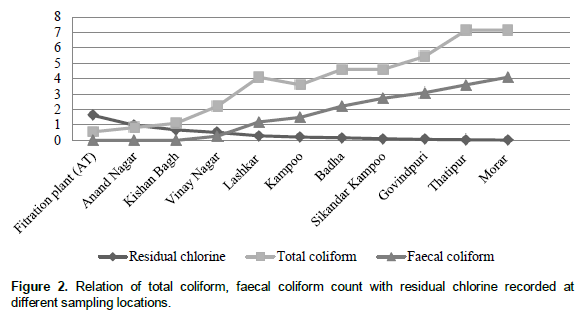
Out of the orders sampled, Hemiptera, Pulmonata and Coleoptera had the highest number of genera with 5, 4 and 4, respectively. In terms of relative abundance, dipterans and Pulmonata were the most abundant while Hydracarina (water mites) were the least abundant (Figure 2). This is an indication that water hyacinth provides a favourable microhabitat for those macroinvertebrates and as a result they associate with the plant. Aquatic organisms such as macroinvertebrates live in selected areas within water bodies depending on the physical, chemical and biological characteristics (Mason 2002; Wang and Lyons, 2003). Due to this fact, they have been used to predict water quality status of different water systems (Masese et al., 2009; Aura et al., 2010). The taxa found represented the sensitive ones such as Trichopterans and the tolerant ones like dipterans and oligochaete worms.
The high diversity obtained can be attributed to favourable water quality conditions and availability of food. Water hyacinth, being a floating macrophyte traps a lot of matter as it floats which could eventually form food for organisms within. Rocha-Ramirez et al. (2007) argues that water hyacinth traps a lot of organic matter as it floats and moves with it from one point to another. Another reason for the high diversity could probably be its ability to protect the invertebrates from direct sunlight, absorb excess nutrients and offer breeding grounds. Aquatic macrophytes like water hyacinth provide shade and the roots provide filtration, a spawning area for the aquatic organisms like macroinvertebrates and fish (Schramm et al., 1987). Water hyacinth could be very effective in nutrient absorption and as a result there were mayflies and caddis flies on the sediments beneath the mat.
For the case building invertebrates, the water hyacinth stem provided a soft material that is convenient to them. Some even burrowed through the soft stem and hatched their eggs inside. It can thus be summarized that a mat of water hyacinth provides food, shelter and breeding ground to macroinvertebrates as such a microhabitat on its own.
Macroinvertebrate abundance
Macroinvertebrate abundance was highest at Bulwani (1639 individuals) and lowest at Lwanda Kotieno (61). The beach stations (Asat and Anyanga) recorded higher abundances compared to those away from the shore. Despite this large variation in total abundance, box and whisker plots showed that the most of the whiskers intercept indicative of lack of significant differences between the stations (Figure 3).
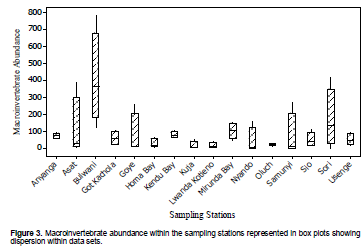
Kruskal-Wallis test showed that the variation in abundance was not significant at 99% confidence level though significant at 95% (H = 22.11, p = 0.033). When the data was log transformed {Log(X+1)} then subjected to one way ANOVA, the variation was still not significant at 99% (F =2.02, p = 0.026). Considering the abundance of the twelve common taxa, the variation was not significant at both 95% and 99% when subjected to one way ANOVA (F = 1.98, p = 0.061).
On pairwise comparison, DMRT revealed the stations responsible for the variation. Over 80% of the stations were at subset b. The stations that differed from the rest were Lwanda Kotieno, Kuja RM, and Oluch RM at subset a and Sori, Asat beach, and Mirunda Bay at subset c.
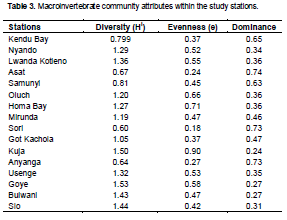
Table 3 shows the values for Shannon-Wiener, evenness and dominance indices using the genera values for the stations sampled during the study period. Shannon-Wiener diversity index was between 0.6 and 1.53. Evenness index was highest at Kuja RM (0.9) and lowest at Sori (0.18). Apart from recording higher evenness index, Kuja RM also had the lowest dominance index which indicates that the organisms at the station coexisted well.
The common taxa were found at specific position in relation to the water hyacinth plant. On the benthic fauna below the plant mainly consisting of algae and sunken water hyacinth plant there were the Hexagenia spp, Lymnae spp and Leptocerus spp together with the cases. On the roots were Tubifex worms, Chironomus spp, Hirudo spp, Melanoids spp, Lymnae spp, Biomphalaria spp, Platycnemis spp, Sympetrum spp and Trithemis spp while on the stem there were the larvae of Elmis and Ilybius spp inside the stem of the plant.
Over 80% of the macroinvertebrates sampled beneath the water hyacinth mat were predators mainly feeding on mosquito larvae (Figure 4). The other feeding guilds found according to literature were collector gatherers (2.6%), parasites (1.4%), filter feeder (6.5), and scrapers (1.9%).
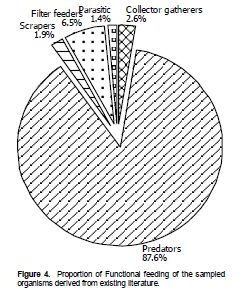
Over 80% of the invertebrates were predators whose main preys are insect larvae. The Hemiptera taxa sampled (Notonectidae, Belostomatidae and Nepidae); Coleoptera (Dytiscidae and Gyrinidae); and Odonata (Gomphidae, Platycnemididae and Libellulidae) are believed to feed mostly on mosquito larvae. This could partly explain the absence of mosquito larvae in most stations despite common belief that mosquitos breed more in areas covered by water hyacinth mats. This corroborates the earlier findings of Ofulla et al (2010). The presence of filter feeders and gathering collectors such as the dipterans (Culicidae, Chironomidae); and some coleopterans (Elmidae) on the root of the plant indicates presence of algae and detritus that are trapped during their float. Research has shown that water hyacinth traps phytoplankton and detritus while floating in a water body or sedentary and lake shores (Rommens et al., 2003; Greenfield et al., 2007).
CONCLUSION
Based on the results and the existing literature, it is concluded that water hyacinth mat provides a microhabitat that favours the survival of several macroinvertebrate taxa. The plant absorbs excess nutrients to make water quality favourable and traps phytoplankton and detritus which in turn becomes food for aquatic life.
RECOMMENDATIONS
We hereby recommend the following based on the findings of this study:
1. Stomach contents of all predators within water hyacinth mats should be examined to check for presence of mosquito larvae.
2. The colonization of water hyacinth by the various macroinvertebrates should be studied to determine the trend and sequence of habitation.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
REFERENCES
|
Aura CM, Raburu PO, Herrmann J (2010). A Preliminary Macroinvertebrate IBI for biossesment of the Kipkaren and Sosiani Rivers, Nzoia River Basin, Kenya. Lakes and Reservoirs. 15(2):119-128. Crossref |
||||
|
de Marco P, Araujo MAR, Barcelos MK, dos Santos MBL (2001). Aquatic invertebrates associated with the water-hyacinth (Eichhornia crassipes) in an eutrophic reservoir in tropical Brazil. Studies on Neotropical Fauna and Environment. 36:73-80. Crossref |
||||
|
Masese FO, Muchiri M, Raburu PO (2009). Macroinvertebrate assemblages as biological indicators of water quality in the Moiben River, Kenya. Afr. J. Aqua. Sci. 34:15-26. Crossref |
||||
| Mason CF (2002). Biology of freshwater pollution. 4th Ed. Pearson Education Ltd. Essex. London. 38pp. | ||||
|
Ofulla AV, Karanja OD, Omondi R, Okurut T, Matano A, Jembe T, Abila R, Boera P, Gichuki J (2010). Relative abundance of mosquitos and snails associated with water hyacinth and hippo grass in the Nyanza gulf of Lake Victoria. Lakes and Reservoirs: Research and Management 15:255-271. Crossref |
||||
|
Rocha-Ramirez A, Ramirez-Rojas A, Chavez-Lopez R, Alcocer J (2007). Invertebrate assemblages associated with root masses of Eichhornia crassipes (Mart.) Solms-Laubach 1883 in the Alvarado Lagoon al System, Veracruz, Mexico. Aqua. Ecol. 41(2):319-333. Crossref |
||||
|
Rommens W, Maes J, Dekeza N, Inghelbrecht P, Nhiwatiwa T, Holsters E, Ollevier F, Marshall B, Brendonck L (2003). The impact of water hyacinth (Eichhornia crassipes) in a eutrophic subtropical impoundment (Lake Chivero, Zimbabwe). I. Water quality. Archiv Fur Hydrobiologie 158:373-388. Crossref |
||||
|
Schramm HL, Jirka KJ Jr, Hoyer MV (1987). Epiphytic macroinvertebrates on dominant macrophytes in two central Florida Lakes. J. Freshwater Ecol. 4 (2). Crossref |
||||
|
Toft JD, Simenstad CA, Cordell JR, Grimaldo LF (2003). The effects of introduced water hyacinth on habitat structure, invertebrate assemblages, and fish diets. Estuaries 26:746-758. Crossref |
||||
| Villamagna, A. M. (2009). Ecological effects ofwater hyacinth (Eichhornia crassipes) on Lake Chapala, Mexico. D. Phil thesis, Virginia Polytechnic Institute and State University. | ||||
| Wang L, Lyons J (2003). Fish and Benthic Macroinvertebrate Assemblage as Indicators of Stream Degradation in Urbanizing Watersheds. In Biological Response Signatures: Indicator Patterns Using Aquatic Communities (Simon, T.P., ed.), CRC Press, Boca Raton: pp. 113-120. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


