Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS



RESULTS AND DISCUSSION


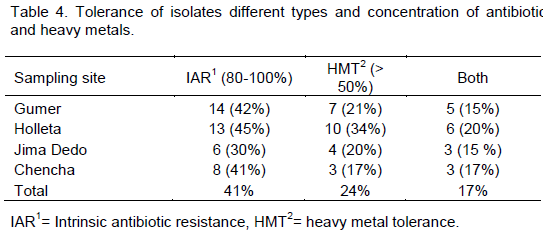
CONCLUSION

CONFLICT OF INTERESTS
The authors do not declare any conflict of interest.
REFERENCES
|
Abere Minalku, Heluf Gebre Kidan, Fasil Assefa (2009). Symbiotic effectiveness and characterization of Rhizobium strains of faba bean (Viciae faba L.) collected from Eastern Western Hararghe highland of Ethiopia. Eth. J. Natural Resources. 11(2): 223-244. |
|
|
|
|
|
Adiguzeli A, Ogutco H, Baris O, Karadayi M, Gulluce M, Sahin F (2010). Isolation and characterization of Rhizobium strains from wild vetch collected from high altitudes in Erzurum-Turkey. RomA. Biotechno. Letter. 15: 5017-5024. |
|
|
|
|
|
Amara DS, Kamara AY, Tucker T (1995). Rhizobium and nodulation assessment on nitrogen fixing trees in Sierra Leone. J. Appl. Science. 4: 41-47. |
|
|
|
|
|
Andrade D, Murphy P, Giller K (2002). The diversity of Phaseolus nodulating rhizobial populations in altered by liming of acid soils planted with Phaseolus vulgaris L. in Brazil. Appl. Environ. Microbiol. 68:4025-4034. |
|
|
|
|
|
Anteneh Argaw (2012). Charcterization of symbiotic effectiveness of rhizobia nodulating faba bean (Vicia faba L.) isolated from central Ethiopia. Resea. J. Microbiol. 7(6): 280-296. |
|
|
|
|
|
Aynabeba Adamu, Fassil Assefa, Asfaw Hailemariam, Endashaw Bekele (2001). Studies of Rhizobium inoculation and fertilizer treatment on growth and production of faba bean (Vicia faba) in some yield depleted and yield sustained regions of Semien Showa. SINET: Ethiop. J. Sci. 24:197-211. |
|
|
|
|
|
Brockwell J, Pilka A, Holliday RA (1991). Soil pH is a major determinant of numbers of naturally occurring Rhizobium meliloti in non-cultivated soils in Cental New South Wales. Austr. J. Exp. Agri. 31:211-219. |
|
|
|
|
|
Devi MJ, Sinclair TR, Vadez V (2010). Genotypic variability among peanut (Arachis hypogea L.) in sensitivity of nitrogen fixation to soil drying. Plant Soil. 330:139-148. |
|
|
|
|
|
Girmaye Kenasa, Mulissa Jida, Fasil Assefa (2014). Charcteriazation of phosphate solublizing faba bean (Vicia faba L.) nodulating rhizobia isolated from acidic soils of Wollega. J. Sci. Technol. Arts Res. 3(3):11-17. |
|
|
|
|
|
Gomez KA, Gomez A (1984). Statistical Procedures for Agricultural Research. John Wiely and Sons. New York. |
|
|
|
|
|
Gorfu A (1998). The role of fertilizers and faba bean (Vicia faba) in sustaining production of cereals in different crop rotations in the South-Eastern Ethiopian Highlands. Doctoral Dissertation, Georg-August-Universität Göttingen, Germany. |
|
|
|
|
|
Graham PH (1992). Stress tolerance in Rhizobium and Bradyrhizobium and nodulation under adverse soil conditions. Can. J. Microbiol. 38:475-484. |
|
|
|
|
|
Hawtin GC, Hebblethwaite PD (1983). Back ground and history of faba bean production. In: The Faba bean (Vicia faba L.), pp. 1-22, (Hebblethwaite, P.D., ed.) Butter Worth's, London. |
|
|
|
|
|
Hubbell DH, Kidder G (2003). Biological nitrogen fixation (http://edis.ifas.ufl.edu/SS180). |
|
|
|
|
|
Hungaria M, Vargas MAT (2000). Environmental factors affecting nitrogen fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Rese. 65:151-164. |
|
|
|
|
|
ICARDA (International Center for Agricultural Research in the Dry Areas) (2006). Screening techniques for disease resistance in faba bean. Aleppo, Syria. |
|
|
|
|
|
Jordan DC (2005). Familly III. Rhizobiaceae conn 1938, 321. IN: Bergey's manual of systematic Bacteriology, Brenner, J.D., N.R.Krieg and J.T. Staley (Eds.) springer, New York, USA. pp. 324-358. |
|
|
|
|
|
Maatallah JEB, Berrah J. Sanjuan C, Lunch C (2002). Phenotypic characterization of rhizobia isolated from chickpea (Cicer arietinum) growing in Moroccan soils. Agronomy 22:321-329. |
|
|
|
|
|
Montanez A (2000). Overview and case studies on biological nitrogen fixation perspectives and limitations. Prepared for FAO. (http://www.fao.org). |
|
|
|
|
|
Purcino HMA, Festin PM, Elkan GH (2000). Identification of effective strains of Bradyrhizobium for Archis. Pintoi. Trop. Agri. 77: 226-231. |
|
|
|
|
|
Rengel Z (2002). Breeding for better symbiosis. Plant Soil 245:147-162 |
|
|
|
|
|
Roughley RJ, Wahab FA, Sundram J (1992). Internisic resistance to sterptomycin and spectinomycin among root-nodule bacteria from Malysian soils. Soil Biolo. and Biochem. 24:715-716. |
|
|
|
|
|
SAS (Statistical Analysis System) (2002). SAS/STAT User's Guide. Propritery software version 9.0. SAS Inst., Inc., Cary, NC. |
|
|
|
|
|
Schlede H (1989). Distribution of acid soils and liming materials in Ethiopia. Ethiopian Institute of Geological Survey. Report Note 326. |
|
|
|
|
|
Somasegaran P, Hoben HJ (1994). HandBook for Rhizobia; methods in legume-Rhizobium technology. Springer, BerlinHeidelberg New York. 2p. |
|
|
|
|
|
Stowers MD (1985). Carbon metabolism in Rhizobium species. Ann. Rev. Microbiol. 39:89-108. |
|
|
|
|
|
Tekalign Mamo, Haque I, Kamara CS (1988) Phosphorus status of some Ethiopian Vertisols. In: Management of Vertisols in Sub-Saharan Africa. pp. 232-252, Jutzi, S.C., Haque, I., McIntire, J. and Stares, J.E.S., eds). Proceedings of a Conference, 31 August-4 September 1987, ILCA, Addis Ababa, Ethiopia. |
|
|
|
|
|
Tsegaye T (1992). Vertisols of the Centeral highlands, Ethiopia – characterization, classification and evaluation of the phosphorus status. M.Sc Thesis, Alemaya University of Agriculture, Ethiopia. |
|
|
|
|
|
Vijila K, Jebaraj S (2008). Studies on the improvement of Rhizobium- green gram (Vigna radiata (l.) wilczek) symbiosis in low nutrient, acid stress soils. Legu. Rese. 31(2):126-129. |
|
|
|
|
|
Yanmei Y, Huang W, Ge Y (2007). Exopolysaccharide: a novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microbiol. Biotechnol. 24:1059-1065. |
|
|
|
|
|
Zerihun Belay, Fasil Assefa (2011). Symbiotic and phenotypic diversity of Rhizobium leguminosarum bv. viciae isolates (Vicia faba) from Northern Gondar. Afr. J. Biotechnol. 10(21):4372-4379. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


