Full Length Research Paper

ABSTRACT
Bacterial resistance to antibiotics is a serious challenge to human and animal health and all efforts are being put together to resolve the menace. In this study the antibiotic resistance modifying activity of ten plants was established by determination of the minimum inhibitory concentrations (MICs) of the plant extracts, the MICs of the antibiotics alone (amoxicillin, ciprofloxacin, erythromycin and tetracycline) and the MICs of the antibiotics in the presence of sub-inhibitory concentrations of the methanol extracts of some selected medicinal plants including Clerodendron splendens, Cyperus esculentus, Duranta plumieri, Kigelia africana, Kyllinga brevifolia, Momordica charantia, Phyllanthus amarus, Pycnanthus angolensis, Secamone afzelii and Thuja occidentalis against two Gram-positive bacteria (Staphylococcus aureus and Bacillus subtilis) and two Gram-negative bacteria (Escherichia coli and Pseudomonas aeruginosa). The extracts were found to possess varying degree of antimicrobial activity with MICs between 4 and 50 mg/mL. It was observed that 26.9% of the plant extract-antibiotic combinations/interactions resulted in the reduction of activity of antibiotics. Almost 17% of the extract-antibiotic interactions led to the complete loss of activity of the antibiotics and 30.0% of the extract-antibiotic combinations resulted in resistance modulation. Three per cent of the extract-antibiotic combinations/interactions had antibiotics that were not active when used alone but became active in the presence of the extracts and 23.1% extract-antibiotic combinations/interactions had no modifying effect on the individual in vitro activities of the antibiotics. There is need to isolate the bioactive agents from the extracts especially those that potentiated the activity of the antibiotics.
Key words: Bacterial infections, antibiotic resistance, resistance modifying agents; medicinal plants
INTRODUCTION
Antimicrobial resistance by pathogenic microorganisms (bacteria, viruses, fungi, protozoa and helminths) is one of the biggest clinical problems currently facing humanity (Davies and Davies, 2010). Drug combination, as exemplified by the combination of β-lactam antibiotics and β-lactamase inhibitors (for example amoxicillin–clavulanic acid) (Reeves et al., 1978) is one way used to overcome resistance by pathogenic microorganisms. The mechanisms of action of such combinations vary overtly from that of the same drugs acting individually and isolation of a single phyto-constituent thus may alter its importance in exhibiting this effect (Hemaiswarya et al., 2008).
Nature has provided various sources of substances used as medicines and scientists have found medicines for almost all diseases known to man from natural sources (Newman et al., 2000). Mankind has been using antimicrobial agents for the management of infections for a longer period than initially anticipated (Bassett et al., 1980; Cook et al., 1989; Aminov, 2010).
Phylogenetic reconstruction analysis indicates that antibiotic resistance genes have been around long before the antibiotic era (Aminov and Mackie, 2007; Kobayashi et al., 2007). Structure-based phylogeny of serine and metallo-β-lactamases, for example, show that these ancient enzymes have been around for more than two billion years and that some serine β-lactamases have been present on plasmids for thousands of years (Hall and Barlow, 2004; Garau et al., 2005). The development of the β-lactamase and housekeeping genes in Klebsiella oxytoca is highly congruent, meaning that these genes have been evolving for over 100 million years (Fevre, 2005). Thus antibiotic resistance is not a new phenomenon, except that, probably it is now more widespread than before, and it moves round the globe more quickly because of easier and faster movement of mankind.
Antibiotic resistance has become a huge problem in recent times. Patients are dying from infections because the antibiotics used for the treatment of these infections are no more effective because of resistance. Resistance to antibiotics by bacteria and other organisms is now a major public health problem worldwide and antibiotic use has been recognized as the main selective pressure driving the menace.
Plants have been identified as one major source of medicinal agents from nature. Medicinal plants have been used by humans to manage several diseases including infections since time immemorial (Rios et al., 1988). Based on their traditional uses several known medicinal plants have been screened for their antimicrobial activity, phytochemical composition, possible interaction with antibiotics or other medicinal plants, pharmacokinetic, bioavailability and toxicity profiles. However, reports on interactions of antibiotics with plant extracts are scanty and only a few studies have been reported (Rios et al., 1988; Nascimento et al., 2000; Aburjai et al., 2001; Aqil et al., 2005). Natural product scientists have been searching for phytochemicals that could ultimately be developed for use in the treatment of infectious diseases (Cowan, 1999). The antiseptic use of volatile oils has been known to man for centuries and these oils have been obtained from plants using extraction methods such as steam or hydro-distillation. These volatile oils are usually variable mixtures of mostly terpenoids, specifically monoterpenes and sesquiterpenes although diterpenes may also be present (Dorman and Deans, 2000).
Fractionation of Glycyrrhiza glabra L. var. typical, using bioactivity guided methods led to the isolation and characterization of several compounds including 4'-O-methylglabridin, 3'-methoxyglabridin, glabridin and glabrol. Glabrene, hispaglabridin A, hispaglabridin B, 3-hydroxyglabrol formononetin, phaseollinisoflavan, salicylic acid and O-acetyl salicylic acid were also found. Some of these including hispaglabridin A, hispaglabridin B, 4'-O-methylglabridin, glabridin, glabrol and 3-hydroxyglabrol were found to possess significant antimicrobial activity (Mitscher et al., 1980).
Apart from being sources of antimicrobial agents, plants also harbour substances that act to modify the resistance offered by microorganisms to antibiotics (Dapaah et al., 2016; Fankam et al., 2015; Seukep et al., 2016). In a study using the multidrug-resistant Escherichia coli that produces extended-spectrum β-lactamases (ESBLs), such as the CTX-M enzymes, the methanol extracts of Gundelia tournefortii L. and Pimpinella anisum L. enhanced the antibacterial activity of chloramphenicol, neomycin, doxycycline, cephalexin and nalidixic acid (Darwish and Aburjai, 2010). In this study, the antibacterial activity of selected plant extracts as well as their in vitro resistance modulatory effects on selected reference antibiotics were determined.
MATERIALS AND METHODS
Collection and identification of plant materials
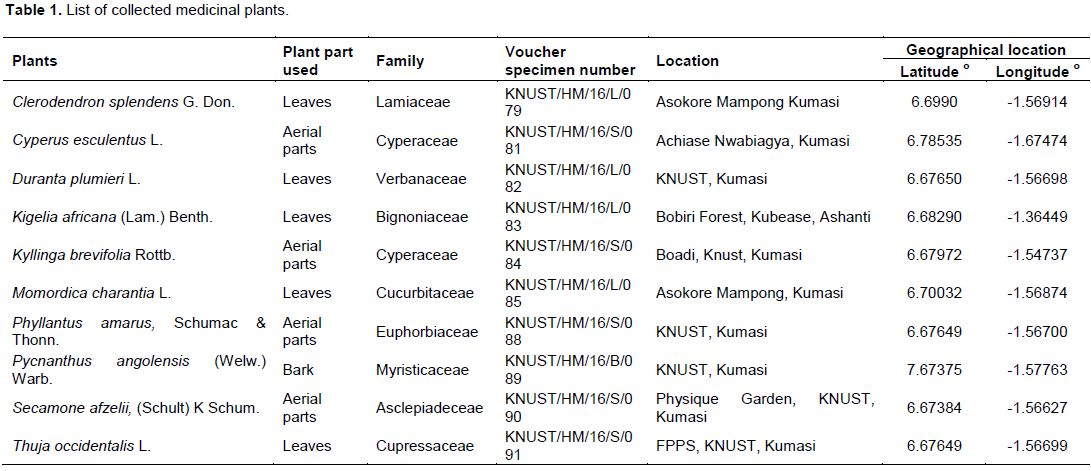
The plants materials were identified, collected and authenticated by Dr. G. H. Sam of the Department of Herbal Medicine, Faculty of Pharmacy and Pharmaceutical Sciences, Kwame Nkrumah University of Science and Technology (KNUST), Kumasi, Ghana with voucher specimens kept at the herbarium of the above mentioned institution (Table 1).
Preparation of plant extracts
The various plant parts were washed under running tap water and air dried individually at room temperature (28 to 32°C) for 7 to 14 days (Meyer and Dilika, 1996; BariÅŸ et al., 2006) to a constant weight. The dried plant materials were milled using a laboratory hammer mill into coarse powder. Each powdered plant material (100 g) was cold macerated with 500 mL of 70% v/v methanol for five (5) days with continuous stirring. They were then filtered using Whatman paper No.1 under reduced pressure. They were then evaporated to dryness using rotary evaporator under reduced pressure at 38°C, lyophilized and stored in a fridge at 4°C.
Antibacterial study
Determination of antibacterial activity of extracts
The antibacterial activities of the crude methanol extracts of the various plant parts were determined using the method as described by Das et al. (2011), Bhalodia and Shukla (2011) and KlanÄnik et al. (2010). One millilitre (1 mL) each of the test organism suspension (S. aureus ATCC 25923, B. subtilis NTCC 4853, E.coli ATCC 25922, P. aeruginosa ATCC 27853) was inoculated into a 10 mL nutrient broth (Oxoid, London UK) and incubated for 24 h and adjusted with normal saline to 0.5 McFarland Standard. Aliquots of 10 μL were applied to the surface of 20 mL of Mueller-Hinton agar (Sigma-Aldrich, Steinheim, Germany) in 10 cm-wide plates. The aliquot was spread over the surface using a sterile cotton wool swab. Three (3) cups were bored in each plate using a cork borer No. 5 (10 mm). Two grams of the extract was added to 1 mL of dimethyl sulfoxide (Sigma-Aldrich, Steinheim, Germany) and diluted to 200 mg/mL with sterile distilled water. It was then serially diluted and the wells filled with 100 µL of 30 and 50 mg/mL of the extract and allowed to diffuse at room temperature (25 to 28°C) for 1 h. A 10 μg/mL solution of tetracycline prepared in water was used as the positive control. The plates were incubated at 37°C for 24 h and zones of growth inhibition recorded. The procedure was performed in triplicates.
Determination of the minimum inhibitory concentration of extracts
The minimum inhibitory concentration (MIC) of all the plant extracts was determined using the macrobroth dilution method (Okeke et al., 2001; Wiegand et al., 2008). Solutions of the extracts were prepared by mixing 4 g of extract with 0.5 mL of dimethylsulfoxide (DMSO) and 9.5 mL of distilled water added to produce a 400 mg/mL stock solution. Specified volumes (5 to 250 μL) were then taken from the stock solution and added to 0.5 mL double strength Muller-Hinton broth (Sigma-Aldrich, Steinheim, Germany). Appropriate volume of sterile distilled water was added and 10 μL of test organism containing 1 x 105 cells per mL was added to the appropriate well to produce concentrations of 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 mg/mL. They were incubated at 37°C for 24 h, after which 10 μL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT 0.1% w/v) was added to each well to detect the presence of growth after 30 min of incubation at 37°C. The MICs were re-determined with a constant difference of 2 mg/mL.
The MICs of reference antibiotics (amoxicillin, erythromycin, tetracycline and ciprofloxacin) were similarly determined against S. aureus, B. subtilis, E. coli and P. aeruginosa using the broth microdilution method (Wiegand et al., 2008; Qaiyumi and Wanger, 2007). The reference antibiotics at 1, 2, 4, 8, 16, 32, 64 and 128 μg/mL were prepared in 96-well plates and the volumes adjusted to 190 μL and 10 μL of test organism suspension containing 105 cells per mL was added to make it 200 μL per well. They were incubated at 37°C for 24 h. The MIC was recorded as the least concentration that showed no visible bacterial growth which was detected by the absence of purple colour after the addition 10 μL of MTT (0.1% w/v) to each well followed by incubation at 37°C for 30 min.
Screening of extracts for antibacterial resistance modifying activities
The antibiotic resistance modifying activity was determined for all the extracts against the test organisms in the presence of the four reference antibiotics (amoxicillin, erythromycin, ciprofloxacin and tetracycline). The MIC of the reference antibiotics was determined against S. aureus, B. subtilis, E. coli and P. aeruginosa (as described above) in the presence of a sub-inhibitory concentration of 2 mg/mL of extracts by incorporating 10 μL of a 200 mg/mL stock solution of the extract into 100 μL of double strength Muller-Hinton broth. The broth was adjusted to 190 μL with sterile distilled water and 10 μL of test organism was added to produce 200 μL. The plates were then incubated at 37°C for 24 h. The MIC was recorded as the least concentration that showed no visible bacterial growth. This was detected by the absence of purple colour after the addition 10 μL of MTT (0.1% w/v) to each well followed by incubation at 37°C for 30 min. The experiment was done in three replicates
RESULTS
Antibacterial activity of extracts
Antibacterial activity showed that the methanol leaf extract of C. splendens, methanol aerial parts extract of P. amarus, methanol leaf extract of T. occidentalis and aerial part methanol extract of C. esculentus had activity against all test organisms at test concentrations with zones of growth inhibition in the range of 11.3±0.33 to 19.6±0.33, 12.3±0.33 to 19.3±0.33, 13.7±0.33 to 19.6±0.33 and 11.6±0.33 to 15.7±0.88 mm, respectively (Table 2). Additionally, the methanol leaf extract of K. africana, methanol aerial part extract of K. brevifolia, methanol leaf extract of M. charantia and aerial part extract of S. afzelii exhibited activity against all the test organisms at 50 mg/mL. The methanol leaf extract of D. plumieri showed no activity against S. aureus and P. aeruginosa at test concentrations. Methanol bark extract of P. angolensis also showed no activity against test Gram-negative organisms (Table 2).
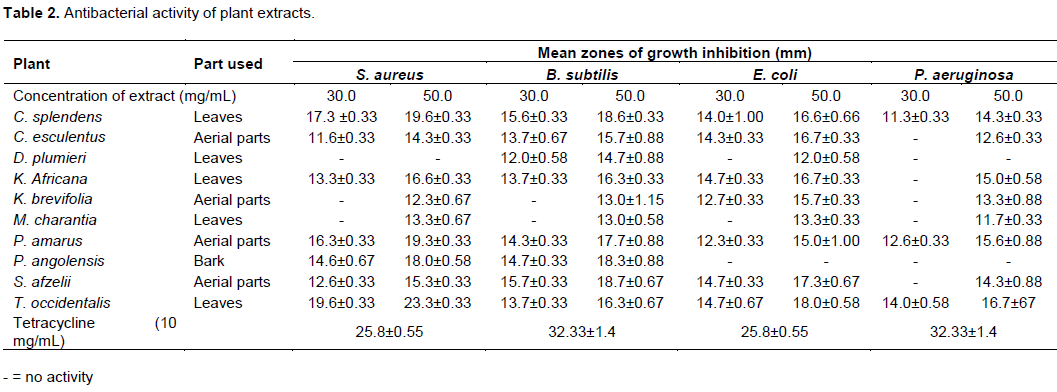
MIC of extracts
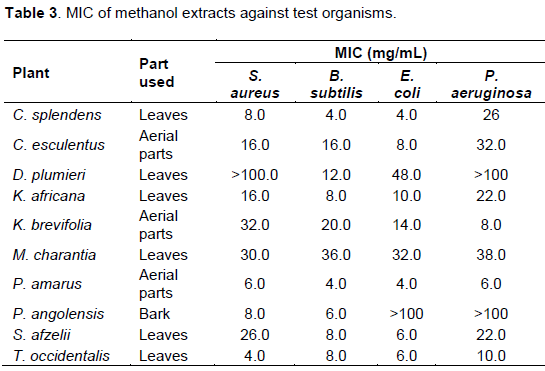
The Gram-positive bacteria were more susceptible to the extracts with lower MICs compared to the Gram-negative bacteria (Table 3). P. amarus and C. splendens were also very active against B. subtilis with MICs of 4 mg/mL. Methanol extracts of M. charantia and K. brevifolia, exhibited weak activity against B. subtilis with MICs of 36 and 20 mg/mL, respectively (Table 3). Methanol leaf extract of T. occidentalis had MIC of 4 mg/mL against S. aureus. S. afzelii, M. charantia and K. brevifolia extracts exhibited weak activity against S. aureus with MICs of 26, 30 and 32 mg/mL, respectively. C. esculentus showed moderate activity against S. aureus and B. subtilis with MIC of 16 mg/mL (Table 3).
E. coli was susceptible to the leaf extract of P. amarus and C. splendens with MICs of 4 mg/mL. The MIC of C. esculentus against E. coli was 8 mg/mL. Additionally, susceptibility of E. coli to extracts of M arantia and D. plumieri was quite low with MICs of 32 and 48 mg/mL, respectively. P. aeruginosa was comparably, the most resistant organism to all the extracts but was most sensitive to P. amarus with MIC of 6 mg/mL. It showed very little susceptibility to C. esculentus, S. afzelii, M. charantia, K. africana and C. splendens with MICs of 32, 22, 38, 22, 32 and 26 mg/mL, respectively (Table 3).
Some of the extracts did not show activity at all against the test organisms within the range of concentrations used. The methanol extract of D. plumieri was not active against S. aureus and P. aeruginosa while P. angolensis showed no activity against the Gram-negative bacteria (Table 3).
Antibiotic resistance modifying/modulation activity of extracts
Influence of C. splendens extract on activity of selected antibiotics
Sub-inhibitory concentrations of methanol leaf extract of C. splendens had minimal effects on the activities of the antibiotics (Table 4). The extract enhanced the activity of tetracycline against P. aeruginosa by reducing the MIC by 8 folds (Table 14). Most of the interactions between the organisms and the antibiotics in the presence of the sub-inhibitory concentration of methanol extracts of C. splendens resulted in marginal changes in their MICs (Tables 4 and 14).

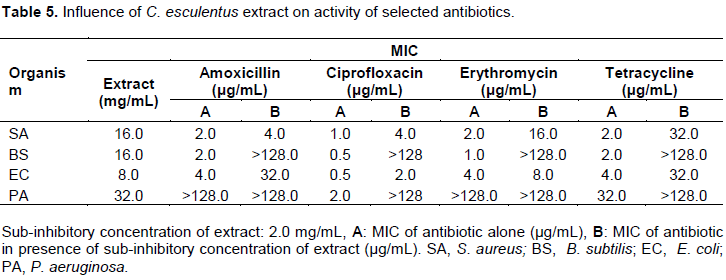
Influence of C. esculentus extract on activity of some selected antibiotics
The MICs of the reference antibiotics were determined with and without the sub-inhibitory concentration (2.0 mg/mL) of methanol aerial parts extract of C. esculentus. It was established that the antibacterial activities of all the reference antibiotics against all the organisms were reduced (Tables 5 and 14).
Influence of D. plumieri extract on activity of selected antibiotics
The presence of sub-inhibitory concentration of methanol leaf extract D. plumieri (2 mg/mL) reduced the activity of the antibiotics against most of the test organisms (Table 6). With the exception of E. coli, S. aureus and P. aeruginosa that saw increased sensitivity to ciprofloxacin, erythromycin and tetracycline producing 2.5, 2 and 8-folds reduction in resistance respectively almost all the interactions produced resistance enhancement (Table 14).
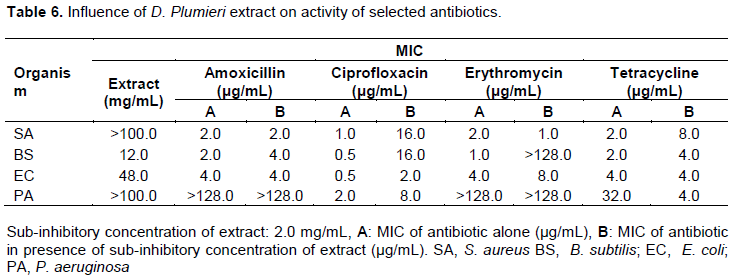
Influence of K. africana extract on activity of selected antibiotics
Methanol leaf extracts of K. africana modulated the susceptibility of the test organisms to the antibiotics. The resistance of B. subtilis reduced against all the antibiotics with amoxicillin showing the greatest increase in activity producing a 15-fold decrease in MIC. In the presence of the extract, E. coli lost sensitivity to all the antibiotics. The sensitivity of P. aeruginosa to tetracycline increased by 8-fold (Tables 7 and 14).
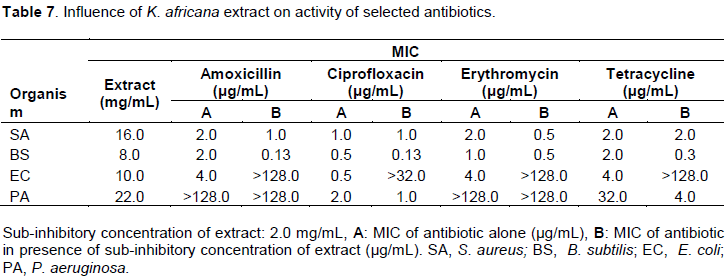
Influence of K. brevifolia extract on activity of selected antibiotics
The methanol aerial parts of extract of K. brevifolia nullified the activity of amoxicillin against S. aureus and B. subtilis and also tetracycline against B. subtilis. The extract made P. aeruginosa sensitive to amoxicillin with an MIC of 32 µg/mL, while producing 2 and 4 fold enhancement in the activities of erythromycin and tetracycline against E. coli, respectively Tables 8 and 14).
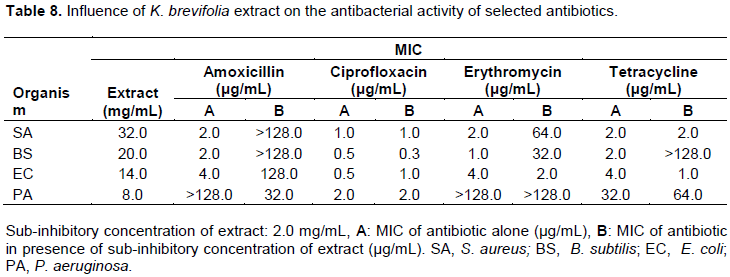
Influence of M. charantia extract on activity of selected antibiotics
In the presence of sub-inhibitory concentration (2 mg/mL) of methanol leaf of M. charantia, S. aureus and B. subtilis lost their sensitivity to tetracycline completely (Table 9). The resistance of S. aureus was reduced 66 folds to amoxicillin while that of P. aeruginosa and E. coli were reduced 15 and 13 folds with MICs of 0.13 and 0.3 µg/mL, respectively (Table 14).
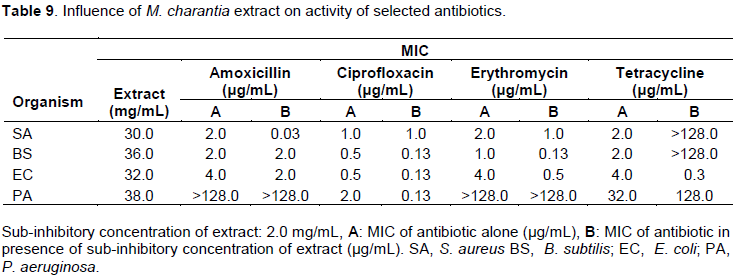
Influence of P. amarus extract on activity of selected antibiotics
In the presence of sub-inhibitory concentration (2 mg/mL) of P. amarus, the organisms were more resistant to amoxicillin with higher MICs than theamoxicillin alone. With the exception of S. aureus, all the organisms were more sensitive to ciprofloxacin in the presence of the extract. Erythromycin lost activity completely against S. aureus rather had lower MIC of 0.13 µg/mL against P. aeruginosa (Table 10). With the exception of S. aureus, the rest of the organisms including B. subtilis, E. coli and P. aeruginosa became more sensitive to tetracycline with 4, 8 and 16 folds reduction in MICs respectively (Table 14).

Influence of P. angolensis extract on activity of selected antibiotics
Methanol bark extract of P. angolensis had marginal effects on the activities of the antibiotics (Table 11). Amoxicillin, ciprofloxacin and erythromycin were more active against S. aureus with 2-fold reduction of their MICs. In the presence of the extract, amoxicillin and ciprofloxacin were less active against B. subtilis (Table 14).
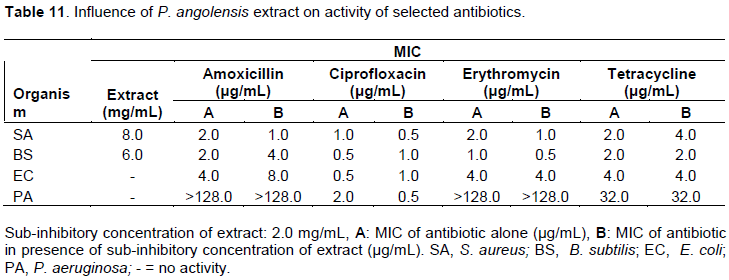
Influence of S. afzelii extract on activity of selected antibiotics
P. aeruginosa was very sensitive to amoxicillin when combined with methanol leaf extract of S. afzelii with an MIC of 16 µg/mL while amoxicillin alone at a concentration of 128 µg/mL had no effect on P. aeruginosa (Table 12). S. aureus, B. subtilis and P. aeruginosa also became less resistant to ciprofloxacin in the presence of the extract. The extract made all the organisms more resistant to erythromycin except P. aeruginosa that became very sensitive to the antibiotic in the presence of the extract with MIC of 8 µg/mL while the antibiotic alone has no activity within the concentrations used. Methanol leaf extract of S. afzelii demonstrated resistance modulation activity on amoxicillin against all the test organisms with 2-fold potentiation of amoxicillin against S. aureus and B. subtilis and an 8-fold potentiation against E. coli. In the case of tetracycline, S. aureus and E. coli lost their sensitivity completely while B. subtilis and P. eruginosa became more susceptible in the presence of the extract (Table 14).
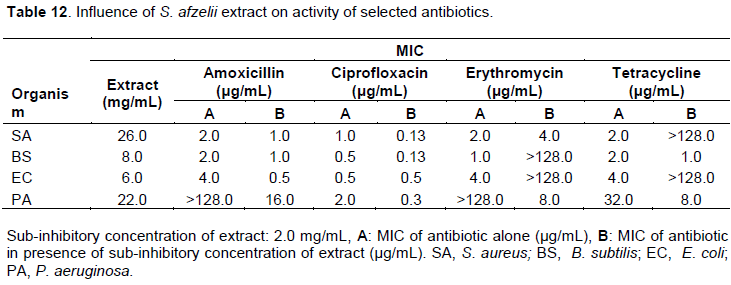
Influence of T. occidentalis extract on activity of selected antibiotics
The methanol leaf extract of T. occidentalis suppressed the activity of all the antibiotics against all the test organisms except in some few cases. S. aureus and B. subtilis lost sensitivity completely to amoxicillin while S. aureus, E. coli and P. aeruginosa lost their sensitivity completely to tetracycline in the presence of the extract. B. subtilis showed a 33 folds reduction in resistance to erythromycin and tetracycline while E. coli also showed an 8 folds reduction in resistance to erythromycin. P. aeruginosa became more sensitivity to erythromycin in the presence of the extract with an MIC of 1 µg/mL (Tables 13 and 14).

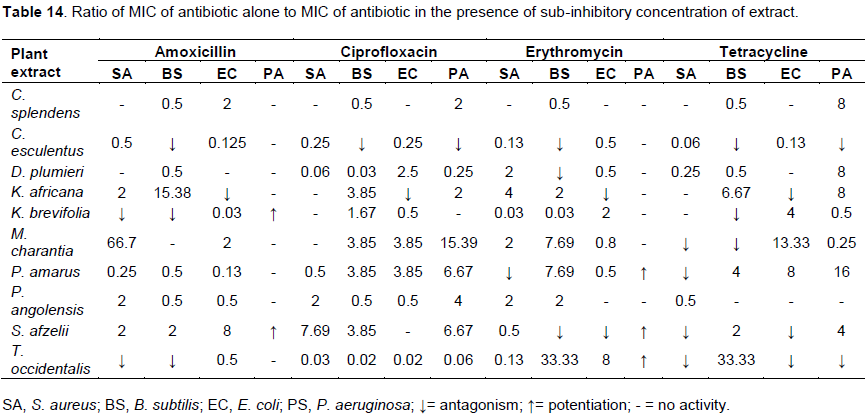
Ratio of activity of antibiotic to activity of antibiotic in the presence of extract
The antibacterial activity of the extracts and reference antibiotics were expressed as MIC, the smaller the MIC the more active the substance. Thus the effect of the sub-inhibitory concentration of the extracts on the activity of the test antibiotics was determined by comparing the ration of MIC of the antibiotic to the MIC of the antibiotic in the presence of sub-inhibitory concentrations of the extract (Table 14).
DISCUSSION
Some plant extracts have been shown over the years to exhibit antimicrobial activity and hence the agar diffusion method was employed to determine the activity of the selected plant extracts against test Gram-positive and negative bacteria. The study revealed that the methanol leaf extract of C. splendens (11.3±0.33 - 19.6±0.33 mm), methanol aerial part extract of P. amarus (12.3±0.33 - 19.3±0.33 mm), methanol leaf extract of T. occidentalis (13.7±0.33 to 19.6±0.33 mm) and aerial part methanol extract of C. esculentus (11.6±0.33 to 15.7±0.88 mm) had activity against all test organisms at test concentrations (Table 2). Methanol leaf extract of C. splendens have been reported to have antibacterial and antiprotozoan activity, the former is in agreement with our studies (Gbedema et al., 2010; Abouzid et al., 2013). MIC of C. splendens methanol extract was determined to be 4.0 to 26 mg/mL against test Gram-positive and negative organisms (Table 3). However, Gbedema et al. (2010) reported a MIC of 128 to 516 µg/mL for extract of C. splendens, indicating a better antimicrobial activity than our findings. The difference may be due to method of extraction, the part of the plant used, site of collection and variation in storage conditions may contribute to the variations in activity (Inbathamizh and Padmini, 2013; Khattak, 2015). The extract of C. splendens enhanced the activity of tetracycline against P. aeruginosa by reducing the MIC by 8 folds (Table 14). This reduction may due to blockade of tetracycline efflux pump activity or modification of the binding site which have been identified as the main mechanism of bacteria resistance to antibiotics including the tetracyclines (John, 2005; Lin et al., 2015).
Aerial parts extract of C. esculentus showed antibacterial activity with zones of inhibition and MIC determined to be between 11.6±0.33 to 16.7±0.33 mmand 8 to 32 mg/mL, respectively for test organisms. This is agreement with the findings of Adeniyi et al. (2014) in which the zone of inhibition was reported to be within the range of 7.5±0.41 to 25.0±0.16 mm for aqueous and ethanol whole plant extracts of C. esculentus. It was also established that the antibacterial activities of all the antibiotics against all the organisms were reduced (Tables 5 and 14). This may probably be due to the extracts ability to reduce efflux pump activity, increase drug uptake and accumulation (Lin et al., 2015; Munita and Arias, 2016).
In the case of D. plumieri leaf extract, the zones of inhibition were determined to be 12.0±0.58 and 14.7±0.88 mm for B. subtilis. No activity was observed for S. aureus and P. aeruginosa but for E. coli (12.0±0.58 mm) at a concentration of 50 mg/mL. This indicates that the extract has weak antibacterial activity which is in agreement with the findings of Adu et al. (2011). The presence of sub-inhibitory concentration of D. plumieri leaf extract reduced the activity of the antibiotics against most of the test organisms (Tables 6 and 14). The reduced or nullified activity of the antibiotics may be as a result of interactions between the phytochemicals in the extract and the antibiotics or the microorganisms. The phyto-constituents may react chemically with the antibiotics leading to loss of activity (Dapaah et al., 2016; Adu et al., 2009).
The methanol leaf extract of K. africana showed activity against test organisms which is consistent with report by Agyare et al. (2013). The ethanolic extracts of K. africana stem bark and fruit extracts have also been reported to possess activity against B. subtilis, E. coli, K. pneumoniae and S. aureus with MICs of 0.63 to 2.5 mg/mL (Grace et al., 2002). The resistance of B. subtilis was reduced against all the antibiotics with amoxicillin showing the greatest increase in activity producing a 15-fold reduction in the MIC. Sensitivity of P. aeruginosa to tetracycline increased by 8-folds. This observed potentiation in activities of the antibiotics may be due to increased drug uptake and its accumulation in bacteria (Lin et al., 2015; Munita and Arias, 2016).
Extracts of K. brevifolia nullified the antibacterial activity of amoxicillin against S. aureus and B. subtilis and also tetracycline against B. subtilis which may be due to the phyto-constituents reacting chemically with the antibiotics leading to loss of activity (Dapaah et al., 2016; Adu et al., 2009). The extract enhanced the sensitivity of P. aeruginosa to amoxicillin with an MIC of 32 µg/mL, while producing 2 and 4 fold potentiation of the activities of erythromycin and tetracycline respectively, against E. coli (Tables 8 and 14). This indicates that the extract possesses phyto-constituents that make the organisms more susceptible to the antibiotics which may be due to increased drug uptake and efflux pump activity inhibition as well as β lactamase inhibition in the case of penicillins (Munita and Arias, 2016; Adu et al., 2009).
For M. charantia, the MIC and zones of growth inhibition was respectively determined to be in the range of 30 to 38 mg/mL and 11.7±0.33 to 13.3±0.33 mm against test organisms. These findings are in agreement with the reported antimicrobial activity of leaf, seed and fruit extracts of M. charantia (Costa et al., 2010; Ozusaglam et al., 2013; de Lucena et al., 2015). In the presence of sub-inhibitory concentration of M. charantia extract, S. aureus and B. subtilis lost their sensitivity to tetracycline completely (Table 9). The resistance of S. aureus was reduced 66 folds to amoxicillin while that of P. aeruginosa and E. coli were reduced 15 and 13 folds with MICs of 0.13 and 0.3 µg/mL, respectively (Table 14). This indicates that the extract contains phytochemicals that have either synergistic or inhibitory effect when combined with the test antibiotics (Adu et al., 2009; Costa et al., 2010).
The aerial part methanol extract of P. amarus was determined to possess antibacterial activity with MIC in the range of 4 to 6 mg/mL which is in agreement with the findings of Babatunde et al. (2014) and Oluwafemi and Debiri (2008). With the exception of S. aureus, the test organisms were more sensitive to ciprofloxacin in the presence of the extract indicating a lowering of efflux pump activity and reduced drug uptake which are the major players in bacterial resistance to the flouroquinolones (Munita and Arias, 2016; Fabrega et al., 2009). With the exception of S. aureus that lost activity completely in the presence of the extract B. subtilis, E. coli and P. aeruginosa became more sensitive to tetracycline with 4, 8 and 16-folds reduction in MICs, respectively (Tables 10 and 14). The observed effect may due to blockade of tetracycline efflux pump activity or modification of the binding site which have been identified as the main mechanism of bacteria resistance to tetracycline (John, 2005; Munita and Arias, 2016).
The methanol bark extract of P. angolensis was active against only S. aureus and B subtilis but not E. coli and P. aeruginosa. The findings of Olademeji et al. (2006) which give credence to our studies indicate that the stem, leaves and root bark extracts of P. angolensis have activity against Gram-positive bacteria but not Gram-negative bacteria. However, Kuete et al. (2011) reported that the root bark extract has antimicrobial activity. In the presence of the extract B. subtilis became more resistant to amoxicillin and ciprofloxacin with the resistance increasing 2 folds (Tables 11 and 14). However, Lambert (2002) reported that some phyto-constituents may act as protein activators or co-enzymes which bind to and activate enzymes or genes responsible for resistance in an organism.
Methanol aerial part extract of S. afzelii was observed have antibacterial activity which is in agreement with previous studies (Lagnika et al., 2011). The observance of both resistance and synergistic effects from the methanol aerial part extract of S. afzelii (Tables 12 and 14) may be due the presence of myriad compounds. These compounds may either potentiate or inhibit the activity of the test antibiotics by activating or inhibiting certain enzymes or genes responsible for their resistance (Dapaah et al., 2016; Adu et al., 2009).
Methanol leaf extract of T. occidentalis had antibacterial activity with MIC of 4 to 10 mg/mL which is in agreement with the findings of Eltayeb and Hamid (2017). However, Sah et al. (2017) reported better antibacterial activity for T. occidentalis extract which may be due to the solvent system (a mixture of ethylacetate chloroform and ethanol in a ratio of 30:30:40) used in their extraction protocol. Additionally, Khubeiz et al. (2016) reported a poorer antibacterial activity of T. occidentalis compared to our findings which may be due to method of extraction (aqueous extraction) and also the geographical as well as the season of collection of plant material which have been reported to influence the levels of the various secondary metabolites in the plants (Inbathamizh and Padmini, 2013; Khattak, 2015). Additionally, B. subtilis showed a 33-fold reduction in resistance to erythromycin and tetracycline while E. coli also showed an 8 folds reduction in resistance to erythromycin. P. aeruginosa gained sensitivity to erythromycin in the presence of the extract with an MIC of 1 µg/mL (Tables 13 and 14). The increased sensitivity of the test organisms to erythromycin and tetracycline may be due to decreased efflux pump activity, target site modification and reduce drug uptake (John, 2005; Munita and Arias, 2016; Leclercq and Courvalin, 2002).
In general, there was almost 30% reduction in in vitro activities of the antibiotics when they were combined with the plant extracts. This means that the sub-inhibitory concentrations of the extracts inhibited some mechanisms of action of the antibiotics. In some situations, the antibiotic alone had activity against the organism but in the presence of the extract, the activity of the antibiotic was lost completely thus making the organism very resistant to the antibiotic. This may mean that, the extract might have prevented the antibiotic from entering the organism, prevented the antibiotic from reaching its target site after entering the organism, reacted with the antibiotic to produce a non-active compound(s) or, inhibited one or more enzymes that were involved in the mechanisms of antimicrobial action (Dapaah et al., 2016; Adu et al., 2009; Adu et al., 2014).
The study clearly shows that when medicinal plants are combined with antibiotics, microbial resistance which is a global problem can be reduced. Synergistic combinations of medicinal plants and antibiotics may help to reduce emergence of multidrug resistant mutants, toxicity, exhibit more antimicrobial activity and more effective against mixed infections (Bhardwaji et al., 2016; Abascal and Yarnell, 2004). The above findings show the need to exercise caution when it comes to the indiscriminate combination of herbal medicines with antibiotics.
CONCLUSION
Potentiation and antagonistic activities were observed among the plant extract-antibiotic combinations. Synergistic activity observed from combinations of plant extract and antibiotics could help reduce the emergence and effect of multidrug resistant strains responsible for some bacterial infections and these extracts possess phytochemicals which could be exploited for their synergistic or antagonistic effect when combined with antibiotics. It is recommended that further studies be carried out to identify the phytocompounds responsible for the observed potentiation or antagonism when these plant extracts are combined with the selected reference antibiotics.
ACKNOWLEDGEMENT
We are grateful to Mr. Francis Amankwah of Department of Pharmaceutics, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana for his technical assistance.
CONFLICT OF INTERESTS
The authors declare that they have no conflict of interest.
REFERENCES
|
Abascal K, Yarnell E (2004). Herbs and drug resistance: Potential of botanical in drug-resistant microbes. Alternative and Complementary Therapies 1:237-241. |
|
|
Abouzid SF, Wahba HM, Elshamy A, Cos P, Maes L, Apers S, Shahat AA (2013). Antimicrobial activity of some Clerodendrum species from Egypt. Natural Products Research 27(11):1032-1036. |
|
|
Aburjai T, Darwish RM, Al-Khalil S, Mahafzah A, Al-Abbadi A (2001). Screening of antibiotic resistant inhibitors from local plant materials against two different strains of Staphylococcus aureus. Journal of Ethnopharmacology 76(1):359-364. |
|
|
Adeniyi TA, Adeonipekun PA, Omotayo EA (2014). Investigating the phytochemicals and antimicrobial properties of three sedge (Cyperaceae) species. Notulae Scientia Biologicae 6(3):276-281. |
|
|
Adu F, Gbedema S, Annan K (2009). Antimicrobial and resistance modulatory activities of Corynanthe pachyceras. Pharmacognosy Research 1(5):280. |
|
|
Adu F, Gbedema SY, Brown P, Annan K, Boamah VE (2011). Antibacterial and Free Radical Scavenging activity of Duranta plumieri, Linn. International Journal of Pharmaceutical Science Research 2(2):282-287 |
|
|
Adu F, Sam GH, Agyare C, Apenteng JA, Boamah VE (2014). Influence of methanol fruit and leaf extracts of Myristica fragrans (Myristicaceae) on the activity of some antibiotics. African Journal of Microbiological Research 8(19):1982-1986. |
|
|
Agyare C, Dwobeng AS, Agyepong N, Boakye YD, Mensah KB, Ayande PG, Adarkwa-Yiadom M (2013). Antimicrobial, antioxidant, and wound healing properties of Kigelia africana (Lam.) Beneth. and Strophanthus hispidus DC. Advanced Pharmaceutical Sciences 2013. |
|
|
Aminov RI (2010). A brief history of the antibiotic era: Lessons learned and challenges for the future. Frontiers in Microbiology 1(134):1-7. |
|
|
Aminov RI, Mackie RI (2007). Evolution and ecology of antibiotic resistance genes. 271:147–161. |
|
|
Aqil F, Khan MSA, Owais M, Ahmad I (2005). Effect of certain bioactive plant extracts on clinical isolates of β-lactamase producing methicillin resistant Staphylococcus aureus. Journal of Basic Microbiology 45(2):106-114. |
|
|
Babatunde SK, Abubakare AA, Abdulraheem YJ, Ajiboye EA (2014). Antimicrobial activity of Phyllanthus amarus on some humanintestinal facultatively anaerobic flora. International Journal of Medical and Biomedical Research 3(1):52-57. |
|
|
BariÅŸ Ö, Güllüce M, Åžahin F, Özer H, Kiliç H, Özkan H, Sökmen M, Özbek T (2006). Biological activities of the essential oil and methanol extract of Achillea biebersteinii Afan. (Asteraceae). Turkish Journal of Biology 30(2):65-73. |
|
|
Bassett EJ, Keith MS, Armelagos GJ, Martin DL, Villanueva AR (1980). Tetracycline-labeled human bone from ancient Sudanese Nubia (A.D. 350). Science 209(4464):1532-1534. |
|
|
Bhalodia NR, Shukla VJ (2011). Antibacterial and antifungal activities from leaf extracts of Cassia fistula L. Journal of Advanced Pharmaceutical Technology and Reseach 2(1):104-109. |
|
|
Bhardwaji M, Singh BR, Sinha DK, Kumar V, Prasanna Vadhana OR, Varan Singh S (2016). Potential of herbal drug and antibiotic combination therapy: A review approach to treat multidrug resistant bacteria. Pharmaceutica Analytica Acta 7(11). |
|
|
Cook M, Molto EL, Anderson C (1989). Fluorochrome Labelling in Roman Period Skeletons From Dakhleh Oasis, Egypt. American Journal of Physical Anthropology 80(2):137-143. |
|
|
Costa JGM, Nascimento EM, Campos AR, Rodrigues FF (2010). Antibacterial activity of Momordica charantia (Curcubitaceae) extracts and fractions. Journal of Basic and Clinical Pharmacology 2(1):45. |
|
|
Cowan MM (1999). Plant Products as Antimicrobial Agents. Clinical Microbiology Reviews 12(4): 564-582. |
|
|
Dapaah SO, Agyare C, Boakye YD, Appiah T (2016). Modulatory effects of Hilleria latifolia and Laportea ovalifolia on activity of selected antibiotics. Journal of Medicinal Plant Research 10(25):360-366 |
|
|
Darwish RM, Aburjai TA (2010). Effect of ethnomedicinal plants used in folklore medicine in Jordan as antibiotic resistant inhibitors on Escherichia coli. BMC Complementary and Alternative Medicine 10(9):9. |
|
|
Das S, Das K, Dubey V (2011). Inhibitory activity and phytochemical assessment of ethno-medicinal plants against some human pathogenic bacteria. Journal of Medicinal Plants Research 5(29):6536-6543. |
|
|
Davies J, Davies D (2010). Origins and Evolution of Antibiotic Resistance. Microbioliology and Molecular Biology Reviews 74(3):417-433. |
|
|
de Lucena Filho JHS, de Freitas Lima R, de Medeiros, ACD, Pereira JV, Granville-Garcia AF, de Brito Costa EMM (2015). Antimicrobial potential of Momordica charantia L. against multi-resistant standard species and clinical isolates. Journal of Contemporary Dental Practice 16(11):854-858. |
|
|
Dorman HJ, Deans SG (2000). Antimicrobial agents from plants: antibacterial activity of plant volatile oils. Journal of Applied Microbiology 88(2):308-316. |
|
|
Eltayeb I, Hamid A (2017). Phytochemical screening and antimicrobial activity of Thuja occidentalis seeds extracts against the isolated common skin infecting microorganisms. International Journal of Pharmacy and Pharmaceutical Sciences 9(10):131-134. |
|
|
Fabrega A, Madurga S, Giralt E, Vila J (2009). Mechanism of action of and resistance to quinolones. Microbiology and Biotechnology 2(1):40-61. |
|
|
Fankam AG, Kuiate JR, Kuete V (2015). Antibacterial and antibiotic resistance modifying activity of the extracts from Allanblackia gabonensis, Combretum molle and Gladiolus quartinianus against Gram-negative bacteria including multi-drug resistant phenotypes. BMC Complementary and Alternative Medicine 15(1):206. |
|
|
Fevre C, Jbel M, Passet V, Weill FX, Grimont PA, Brisse S (2005). Six groups of the OXY β-lactamase evolved over millions of years in Klebsiella oxytoca. Antimicrobial Agents and Chemotherapy 49(8):3453-3462. |
|
|
Garau G, Di Guilmi AM, Hall BG (2005). Structure-Based Phylogeny of the Metallo-Lactamases. Antimicrobial Agents and Chemotherapy 49(7):2778-2784. |
|
|
Gbedema SY, Kissieh E, Adu F, Annan K, Woode E (2010). Wound healing properties and kill kinetics of Clerodendron splendens G. Don, a Ghanaian wound healing plant. Pharmacognosy Research 2(2):63-68. |
|
|
Grace OM, Light ME, Lindsey KL, Mulholland DA, Van Staden J, Jager AK, Eloff JN (2002). Antibacterial activity and isolation of active compounds from fruit of the traditional African medicinal tree Kigelia africana. South African Journal of Botany 68(2):220-222. |
|
|
Hall BG, Barlow M (2004). Evolution of the serine β-lactamases: Past, present and future. Drug Resistance Updates 7(2):111-123. |
|
|
Hemaiswarya S, Kruthiventi AK, Doble M (2008). Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 15(8):639-652. |
|
|
Inbathamizh L, Padmini E (2013). Effect of geographical properties on the phytochemical composition and antioxidant potential of Moringa oleifera flowers. Journal of Pharmacy Research solutions BioMedRx 1(3):239-247. |
|
|
John MC (2005). Roberts update on acquired tetracycline resistance genes. FEMS Microbiology Letters 245(12):195-203. |
|
|
Khattak KF (2015). Effect of geographical distributions on the nutrient composition, phytochemical profile and antioxidant activity of Morus nigra. Pakistan Journal of Pharmaceutical Sciences 28(5):1671-1678. |
|
|
Khubeiz MJ, Mansour G, Zahraa B (2016). Antibacterial and phytochemical investigation of Thuja orientalis (L.) leaves essential oil from Syria. Int. Journal of Current Pharmaceutical Reviews and Research 7(5):243-247. |
|
|
KlanÄnik A, Piskernik S, Jeršek B, and Možina SS (2010). Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. Journal of Microbiological Methods 81(2):121-126. |
|
|
Kobayashi T, Nonaka L, Maruyama F, Suzuki S (2007). Molecular evidence for the ancient origin of the ribosomal protection protein that mediates tetracycline resistance in bacteria. Journal of Molecular Evolution 65(3):228-235. |
|
|
Kuete V, Nono EC, Mkounga P, Marat K, Hultin PG, Nkengfack AE (2011). Antimicrobial activities of the CH2Cl2–CH3OH (1: 1) extracts and compounds from the roots and fruits of Pycnanthus angolensis (Myristicaceae). Nature Products Research 25(4):432-443. |
|
|
Lagnika L, Anago E., Sanni A (2011). Screening for antibacterial, antioxidant activity and toxicity of some medicinal plants used in Benin folkloric medicine. Journal of Medicinal Plants Research 5(5):773-777. |
|
|
Lambert PA (2002). Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. Journal of Royal Society of Medicine 95(41):22-26. |
|
|
Leclercq R, Courvalin P (2002). Resistance to macrolides and related antibiotics in Streptococcus pneumoniae. Antimicrobial Agents and Chemotherapy 46(9):2727-2734. |
|
|
Lin J, Nishino K, Roberts MC, Tolmasky M, Aminov RI, Zhang L. (2015). Mechanisms of antibiotic resistance. Frontiers in Microbiology 6:34. |
|
|
Meyer JJM, Dilika F (1996). Antibacterial activity of Helichrysum pedunculatum used in circumcision rites. Journal of Ethnopharmacology 53(1):51-54. |
|
|
Mitscher L, Park Y, Clark D (1980). Antimicrobial agents from higher plants. Antimicrobial isoflavanoids and related substances from Glycyrrhiza glabra L. var. typica. Journal of Natural Products 43(2): 259-269. |
|
|
Munita JM, Arias CA (2016). Mechanisms of antibiotic resistance. Microbiol spectr 4(2). |
|
|
Nascimento GG, Locatelli J, Freitas PC, Silva GL (2000). Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Brazilian Journal of Microbiology 31(4):247-256. |
|
|
Newman DJ, Cragg GM, Snader KM (2000). The influence of natural products upon drug discovery. Natural Products Reports 17(3):215-234. |
|
|
Okeke MI, Iroegbu CU, Eze EN, Okoli AS, Esimone CO (2001). Evaluation of extracts of the root of Landolphia owerrience for antibacterial activity. Journal of Ethnopharmacology 78(2):119-127. |
|
|
Oladimeji OH, Ubulom PE, Igboasoiyi AC, Ndukwe K, Nia R (2006). Some biological activities of Pycnanthus angolensis (Welw.) Warb. Journal of Pharmaceutical Bioresources 3(1):49-55. |
|
|
Oluwafemi F, Debiri F (2008). Antimicrobial effect of Phyllanthus amarus and Parquetina nigrescens on Salmonella typhi. African Journal of Biomedical Research 11(2). |
|
|
Ozusaglam MA, Karakoca K (2013). Antimicrobial and antioxidant activities of Momordica charantia from Turkey. African Journal of Biotechnology 12(13):1548-1558 |
|
|
Qaiyumi S, Wanger A (2007). Antimicrobial Susceptibility Testing Protocols R. Schwalbe, L. Steele-Moore, and A. C. Goodwin, eds., London, New York: CRC Press, Taylor and Francis Group. |
|
|
Reeves DS, Bywater MJ, Holt HA (1978). Antibacterial synergism between beta-lactam antibiotics: Results using clavulanic acid (BRL. 14151) with amoxycillin, carbenicillin or cephaloridine. Infection 6(1):9-16. |
|
|
Rios JJ, Recio MC, Villar A (1988). Screening methods for natural products with antimicrobial activity: A review of the literature. Journal of Ethnopharmacology 23(3):127-149. |
|
|
Sah SN, Regmi S, Tamang MK (2017). Antibacterial Effects of Thuja Leaves Extract. International Journal of Applied Science and Biotechnology 5(2):256-260. |
|
|
Seukep JA, Sandjo LP, Ngadjui BT, Kuete V (2016). Antibacterial and antibiotic-resistance modifying activity of the extracts and compounds from Nauclea pobeguinii against Gram-negative multi-drug resistant phenotypes. BMC Complementary and Alternative Medicine 16(1):193. |
|
|
Wiegand I, Hilpert K, Hancock REW (2008). Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nature Protocols 3(2):163-175. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


