Full Length Research Paper

ABSTRACT
Anogeissus leiocarpa (DC) Guill. et Perr. belongs to the Combretaceae family and was previously named A. leiocarpa (DC) Guill. Et Perr. It has been widely used in Burkina Faso by traditional medicine for the treatment of hypertension. Previous study showed that the dichloromethanolic fraction from the barks of trunk of A. leiocarpa (ALF) has induced an endothelium-independent and endothelium-dependent vasodilation effect and had the capacity to inhibit in vitro, purified cyclic nucleotide phosphodiesterases (PDEs) activity. The aims of this study were to better underline ALF-induced endothelium-independent vasorelaxation in an organ model. The results showed that ALF significantly reduce the contractile response to U46619 in porcine coronary artery rings without endothelium that were in concentration-dependent manner. In denuded rings, pretreatment by ALF (10, 30 and 100 µg/mL) did not affect relaxation to sodium nitroprusside (SNP), suggesting that relaxation to ALF was not due to its ability to be a nitric oxide donor. Moreover, SNP-induced relaxation had not been affected in the presence of Nimodipine (PDE1 inhibitor), EHNA (PDE2 inhibitor) or DMPPO (PDE5 inhibitor). In addition, the results showed a relaxation effect to isoproterenol in endothelium-denuded artery rings pretreated with ALF (3, 10, and 30 µg/mL) which were significantly affected suggesting a possible membrane hyperpolarization leading to the vasodilation. In the presence of the PDE3 specific inhibitor Cilostamide and ALF (30 and 100 µg/mL), the vasodilation effects of isoproterenol was enhanced and comparable. Moreover, various potassium channels were not involved in ALF-induced relaxation since tetraethylammonium chloride (non-selective K+ channels inhibitor), iberiotoxin (voltage-sensitive potassium channels inhibitor) and Glibenclamide (KATP channels inhibitor) did not notably affect the relaxation effect to ALF in rings without endothelium. Taken together, ALF-induced endothelium-independent relaxation mainly involves a sustained decrease in [Ca2+]i and may be due to PDE1, 3 and 5 inhibitions localized in the vascular smooth muscle cells. While, the involvement of the ions channels have not been clearly revealed in this experiment.
Key words: Anogeissus leiocarpa, U46619, phosphodiesterases (PDEs), calcium, porcine coronary artery, sodium nitroprusside (SNP), isoproterenol.
INTRODUCTION
The traditional medicine has always been used in Burkina Faso for the care of the population and this for several generations. In cardiovascular diseases such as stroke, headache, and hypertension, vessels dilation is fundamental because it directly influences the arteries of the circulatory system. Accordingly, many researchers have investigated the vasorelaxant effects of various herbal medicines (Gan et al., 2016; Kassahun Gebremeskel et al., 2017; Cam et al., 2018; Khan et al., 2018; Vajic et al., 2018; Sanchez-Recillas et al., 2019). It was the case of a recent study which indicated the endothelium-dependent vasodilation effect of the aqueous extract of Anogeissus leiocarpa (Belemnaba et al., 2018).
Anogeissus leiocarpa is commonly named African birch. It has large ecological distribution ranging from the boarders of Sahara up to the out layer humid tropical forests. In West Africa, it expands from Senegal to Cameroon, with extension to Ethiopia and East Africa. It has been grown in dry and gallery forests. The tree was up to 30 m in height but typically, between 15 and 18 m with light green foliage. The base of the trunk is wider and occasionally striped. The colour of the bark is grey and becomes blackished depending on the age. It is fibrous with thin scales. It has a finely publescent stems and alternate to sub-opposite, elliptical to oval leaves which are 2 to 8 cm length and 1.5 to 3.5 cm wide (Arbonnier, 2009).
However, there have been fewer pharmacological studies and clinical data for A. leiocarpa in comparison to other medicinal plants used for their pharmacological effects through their molecules contain (Notopterygii Rhizoma and Radix; Calotropis procera, Kaya senegalensis, Moringa oleifera).
Therefore, more pharmacological and clinical studies are needed to support the continuous use of A. leiocarpa in traditional medicine for the treatment of hypertension.
Nevertheless, studies on A. leiocarpa extracts had already reported various pharmacological activities such as antioxidant, antimicrobial, anthelmintic activity and antihypertensive effect (Mann et al., 2008; Konaté et al., 2011; Soro et al., 2013; Ouedraogo and Kiendrebeogo, 2016). Many second messengers are involved in the regulation of the cardiovascular system with specifications depending on whether it is at the heart or vascular level. At the heart level, regulation involves several systems such as the orthosympathetic and parasympathetic systems in which intracellular Ca2+ as the main regulator of cardiac excitation-contraction coupling. An imbalance in the intracellular calcium concentration inevitably leads to heart dysfunction (Chung et al., 2016; Mora et al., 2017; Montalvo et al., 2018). At the vascular level, regulation in the presence of endothelium involves many second messengers such as nitric oxid, prostacyclin I2, inhibition of phosphodiesterases (PDEs), endothelium-derived hyperpolarizing factors and many others messengers. In the absence of endothelium, this relaxation notably involves the potassium channels, calcium channels, inhibition of PDEs and other vasodilation factors (Alamgeer et al., 2018; Jia et al., 2018; Sanchez-Recillas et al., 2019). Specifically for A. leiocarpa fraction (ALF), previous studies have shown that this extract has induced an endothelium-dependent and endothelium-independent vasorelaxant effect on pig artery by involving mostly NO/sGC/cGMP and Na+/k+-ATPase pathways and in some extent a possible PDEs inhibition (Belemnaba et al., 2013). However, there was no published studies on this extract demonstrating that its vasorelaxation effect implicates the inhibition of PDEs in a vascular system. Indeed, the possible implication of the PDEs inhibition in the mechanism of vasodilation of ALF has been demonstrated but through PDEs purified according to a tube model (Belemnaba et al., 2013). It was therefore necessary to show this implication on a model using vessels which is close to reality. Moreover, it was well known that other ions channels were involved in vascular stone regulation especially in endothelium independent arteries. It was the case of large-conductance calcium-activated potassium channels (BKCa), KATP channels and K+ channels (Gan et al., 2016; Greenberg et al., 2016; Li et al., 2018).
For this purpose, we have used pig isolated arteries rings to assess the implication of PDEs inhibitions in the ALF endothelium independent vasodilation effects by using various pharmacologicals agents [bradykinin, sodium nitroprusside (SNP, a NO donor), isoproterenol (Adenylate cylase activator)] and specifc inhibitors [Nimodipine (PDE1 inhibitor), EHNA (PDE2 inhibitor), Cilostamide (PDE3 inhibitor), DMPPO (PDE5 inhibitor), tetraethylammonium chloride (non-selective K+ channels inhibitor), iberiotoxin (voltage-sensitive potassium channels inhibitor) and Glibenclamide (KATP channels inhibitor)].
MATERIALS AND METHODS
Plant and extraction
The plant materials consist of the barks of the trunk of A. leiocarpa.
The collection method of A. leiocarpa has been previously described (Belemnaba et al., 2013).
Briefly, the barks of trunk of A. leiocarpa were collected in May 2006 in the East of Ouagadougou (zone of savana). A voucher specimen (Number 1544) was deposited at the Department of Forest Production of the National Centre for Scientific and Technological Research (CNRST) after identification by a botanist of this centre. Plant material freshly collected was dried in a greenhouse with air circulation. It was powdered in a pulverizer mil and a decoction was made (120 g in 2100 mL distilled water). After the decoction, ALF extract was obtained by an exhaustion into a methylene chloride solution, then concentrated into a rotavapor system and finally dried in an oven at 35°C.
Drugs and chemicals
Bradykinin, isoproterenol (Isop), indomethacin, sodium nitroprusside (SNP), glibenclamide (GLIB), iberiotoxin (IBTX), tetraetylammonium chloride (TEA), bradykinin and the cyclic nucleotides PDE inhibitors (EHNA, Cilostamide) were from Sigma-Aldrich (Saint Quentin Fallavier, France). Nimodipine and DMPPO were given by Bayer (Berlin). U46619 (9,11-dideoxy-11α,9α-epoxymethanoprostaglandin F2α) was from Cayman Chemical (Ann Arbor, MI, United States of America).
Vascular reactivity studies
The vascular reactivity of ALF was assessed as indicated previously (Belemnaba et al., 2018). Pig hearts were collected from the local slaughterhouse (Copvial, Holtzheim) and left circumflex coronary arteries were excised and then cleaned to remove all connectives tissues. Porcine coronary arteries were cut into rings of 3 to 4 mm and suspended in organ bath contained appropriated krebs bicarbonate solution (Composition in mM: NaCl 119, CaCl2 1.25, NaHCO3 25, MgSO4 1.18, KH2PO4 1.18, KCl 4.7, D-glucose 11, pH 7.4, 37°C) and oxygened with carbogen gas (mixture of oxygen 95% and carbon dioxide 5%). Rings were put into an initial isometric tension of 5 g for 1 h.
The presence of endothelium was confirmed pharmacologically by testing the response to bradykinin at 3×10-7 M under pre-contraction conditions with U46619. A relaxation about 90% of the contracted rings by bradykinin (3×10-7 M) was considered with endothelium. When required, the endothelium was removed by gently rubbing the intimal space with a stainless steel rod with a diameter equivalent to that of the arterial lumen. A relaxation about 10% of the maximal contracted rings by bradykinin (3×10-7 M) was considered without endothelium. For the assessment of ALF effect on rings, rings were contracted with U46619 to about 80% of the maximal contraction before the construction of the concentration-response curve to ALF. In some experiments, rings were pre-incubated with specific inhibitors or ALF (3, 10, 30 or 100 µg/mL) before contraction to U46619 and the subsequent construction of a concentration-response curve to an antagonist. Indeed, to determine the effect of the extract on U46619-induced vasoconstriction, rings were first incubated with ALF (3, 10, 30, 100 and 300 μg/mL) for 5 min before a cumulative of U46619. In other experiments, rings were pre-incubated with different concentrations of ALF (3, 10, 30, 100 and 300 μg/mL) for 5 min followed by U46619 to the maximum contraction and subsequent cumulative with SNP or isoproterenol. Other rings were first pre-incubated with PDEs inhibitors (Nimodipine, EHNA, Cilostamide, DMPPO) or with TEA, Glibenclamide or Iberiotoxin for 30 min followed by U46619 contraction and cumulative with ALF (3, 10, 30, 100 and 300 μg/mL).
Statistical analysis
All results were expressed as the mean ± standard error of mean (SEM) and n represents the number of rings from different pigs used in the experiments. The relaxation response was expressed as percentage of decreases in tension from the contracting level induced by U46619. The concentrations of substances induced 50% of maximal relaxation (EC50) and their maximal relaxation (Emax) was determined with GraphPad Prism 5.00.288. Two way or one way ANOVAs (with post hoc Bonferroni’s test) determined significant differences, if any, between concentration-relaxation curves and EC50, respectively in different treatment groups. P < 0.05 was considered as the significant threshold.
RESULTS
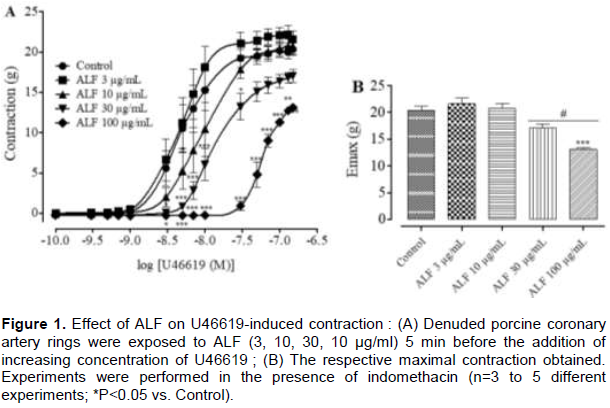
Effect of ALF (3-100 µg/mL) on U46619-induced contraction in rings with endothelium
The thromoxane A2 mimetic U46619 (10-10 M to 3×10-7 M) has induced a concentration-dependent contraction of pig coronary rings (Figure 1). The magnitude of U46619-induced isometric tension development on rings without endothelium was in a concentration-dependent manner. The maximum contraction-response (Emax) to U46619 in rings pre-incubated with ALF 3, 10, 30 and 100 μg/mL was of 21.57±1.88, 20.73±1.81, 17.04±1.56 and 13.11±0.51 g, respectively (Figure 1B) while the control was 20.34±1.69 g. The pre-incubation by ALF 30 and 100 μg/mL causes a significant reduction in the contraction-induced by U46619 as compared to the control. Moreover, we noted a slight but not significant shift to the left and to the right contraction-curve when rings were preincubated with ALF 3 and 10 µg/mL, respectively compared to the control.
Effects of ALF (3-100 µg/mL) on SNP-induced vasodilation in porcine arteries rings
The results showed that the endothelium-independent vasodilator sodium nitroprusside (SNP, 10-11 to 10-5 M) was dose-dependently relaxed U46619-contracted pig coronary arteries rings without endothelium. In pre-incubated denuded artery rings with ALF (3, 10, 30 or 100 µg/mL), the vasodilator effect of SNP (a NO donnor) was significantly improved in a concentration-dependent manner compared to the control (Figure 2). The half-maximal effective concentration (EC50) values were of 0.171±0.076 µM for the control, 0.054±0.019, 0.050±0.034, 0.043±0.017 and 0.005±0.002 µM in the presence of ALF 3, 10, 30 and 100 µg/mL, respectively. No significant difference was notified in the Emax values that were 99.51±1.72, 100.27±0.37, 101.62±1.78, 100.2±0.35, and 100±0, respectively for the control, ALF 3, 10, 30 and 100 µg/mL.

Effects of Nimodipine (1 µM), EHNA (10 µM), and DMPPO (0.1 µM) on SNP-induced vasodilatation in porcine arteries rings
The concentration-relaxation curves to SNP in denuded rings contracted with U46619 were significantly shifted to the left when pre-incubated with Nimodipine (PDE1 specific inhibitor),EHNA (PDE2 specific inhibitor)or DMPPO(PDE5 specific inhibitor) compared to the control (Figure 3).

The EC50 values of SNP-induced endothelium-independent vasodilation on rings were of 0.063±0.040, 0.059±0.015 and 0.022±0.007 µM, respectively in the presence of Nimodipine (1 µM), EHNA (10 µM), and DMPPO (0.1 µM). When comparing the EC50 values, only the presence of DMPPO showed a significant difference compared to the control (EC50 = 0.171±0.076 µM) but their Emax was not affected.
Effects of ALF (3-100 µg/mL) on isoproterenol-induced vasorelaxation in porcine arteries rings
Isoproterenol caused concentration-dependent relaxation of U46619 pre-contracted in pig coronary artery without endothelium (EC50 = 0.148±0.027 µM). Pre-incubation with ALF (3, 10, 30 and 100 µg/mL) has been potentiated significantly in the relaxation effect of isoproterenol in a concetration-dependent manner compared to the control. The EC50 was of 0.08±0.03, 0.04±0.009, 0.02±0.01 and 0.01±0.002 µM in the presence of ALF 3, 10, 30 and 100 µg/mL, respectively. However, the Emax values were not significantly modified compared to the control (Figure 4).

Effects of ALF (3 µg/mL), nimodipine (10 µM) and DMPPO (0.1 µM) on isoproterenol-induced vasodilatation in porcine arteries rings
The results shown in Figure 5 indicated that the vasodilation effect to isoproterenol was not notably improved when rings were preincubated either with nimodipine (PDE1 specific inhibitor) or with DMPPO (PDE5 specific inhibitor). The shape of their relaxation curves was similar to that following the pre-incubation with ALF 3 µg/mL. The EC50 values of isoproterenol were of 0.148±0.027 µM for the control and of 0.076±0.030, 0.123 ±0.091 and 0.112±0.076 µM in the presence of ALF 3 µg/mL, nimodipine and DMPPO, respectively but without significant difference to the control.
Effects of ALF (30 and 100 µg/mL) and cilostamide (10 µM) on isoproterenol-induced vasodilation in porcine arteries rings

The results showed that the vasodilation effect to isoproterenol was higly and significantly improved when rings were pre-incubated with cilostamide (a PDE3 specific inhibitor, 10 µM; Figure 6). This effect was relatively similar to those obtained in the presence of ALF30 and 100 µg/mL. The EC50 of isoproterenol was of 0.030±0.007, 0.023±0.007 and 0.014±0.002 µM in the presence of cilostamide, ALF 30 and 100 µg/mL, respectively. Thus, the rank orders for relaxant effect of isoproterenol when pre-incubated with ALF and cilostamide were ALF 100 µg/mL > ALF 30 µg/mL > Cilostamide.

Effects of TEA (1 µM), IBTX (10 µM) and GLIB (0.44 µM) on ALF-induced vasodilation in porcine arteries rings
In endothelium denuded artery rings, results showed that the non-selective blockade of potassium channels with TEA was not affected. ALF (0.1 to 30 µg/mL)-induced vasodilation effect that was superposable to those of the control (Figure 7). When pre-incubated with IBTX or Glibenclamide, the relaxation of ALF in rings were slighly shifted but not significantly different to the control. The Emax values were 97.41±6.84% for the control and 100% in the presence of the tested inhibitors.

DISCUSSION
In previous study, it was demonstrated that ALF has been induced as an endothelium-dependent and endothelium-independent vasodilation effect in porcine coronary arteries rings that was mediated via NO/sGC/cGMP and cAMP pathways, potassium (K+) and partly Na+/K+-ATPase channels opening (Belemnaba et al., 2013). Moreover, these findings indicated that ALF was a powerful inhibitor of PDE1, 2, 4 and PDE5 with a less activity on PDE 3. While these tests were conducted on tubes assay and on purified PDEs.
In the present study, experiments were focus on the endothelium-independent vasodilaion effect of ALF in order to better characterized its underline mechanism of vasodilation and also to show the implication of PDEs inhibiton activity by this fraction of A. leiocarpa in an organ model.
Thus, in endothelium-denuded coronary artery ring, vasoconstriction was induced by the thromboxane A2 analogue U46619. The thromboxane A2 (TXA2) was a pharmacological agonist known to be a potent constrictor of artery smooth muscle as well as an inducer of platelet aggregation but also a major cyclooxygenase-dependent endothelium-derived contracting factor (Fu et al., 2017; Minuz et al., 2018; Xu et al., 2018). Because thromboxane A2 was an unstable component, the stable U46619 was used to contract the coronary arteries rings in the presence or absence of active substances. Indeed, the U46619 was a full TXA2 receptor agonist that induced a susbtained contraction in porcine coronary arteries rings by an influx of Ca2+ transmembrane leading to the increase of intracellular Ca2+ (Hanasaki et al., 1988; Han et al., 1995; Cogolludo et al., 2003; Hahnenkamp et al., 2004; Sinharoy et al., 2017). Other studies have been also indicated that the important mechanism by which TXA2 elicits its role in pulmonary vasoconstriction was through the involvement of an increase in [Ca2+]i in the smooth muscle cells (Chakraborti et al., 2009).
The present findings showed that ALF had significantly reduced the U46619-induced vasoconstriction in porcine coronary artery rings without endothelium and interestingly, this effect was in a concentration-dependent manner (Figure 1). For that, ALF effect could be explained by a possible decrease in [Ca2+]i release in the vascular smooth muscles cells (VSMC) since U46619 was known to induce vasoconstriction in vessels by activation of PLC via Gaq, followed by an increase in intracellular free calcium concentration through an inositol-3-phosphate stimulation sensitive receptors (Hirata et al., 1991; Somlyo and Somlyo, 2000; Bhattacharya et al., 2005).
In order to determine the possible implication of Ca2+ decrease in ALF-induced vasodilation effect in vascular system, the effect of SNP was investigated in denuded artery rings pre-incubated with different concentrations of ALF and contracted with U46619 (Figure 2). In this study, results showed that in the functional vascular study using wire myograph and SNP which acts directly on VSMC, that ALF acutely displays an increase in endothelium-independent relaxation in a concentration-dependent manner compared to the control.
In the literature, the SNP (a NO donor) was a well-known arterial and venous vasodilator used in clinical practice to lower blood pressure and usually used to increase the endothelium-independent vasodilatation effect by a cGMP independent mechanism (Otsuka et al., 1988; Hottinger et al., 2014; Basrali et al., 2015). Indeed, several studies demontrated that in vascular smooth muscle, the NO has activated the soluble guanylate cyclase (sGC) that stimulate the production of cyclic GMP, which induced a reduction in the concentration of cytosolic Ca2+ with consequent vascular relaxation (Rapoport and Murad, 1983; Hottinger et al., 2014; Zhao et al., 2015; Montfort et al., 2017). Because ALF has enhanced SNP effect suggested that ALF and SNP may have possible synergetic actions to promote cGMP accumulation and Ca2+ decreased in VSMC.
Moreover, the endothelium-independent relaxation to isoproterenol (a β-adrenoceptor agonist) was enhanced by the presence of ALF which was in a concentration-dependent manner (Figure 4). Because isoproterenol was known to reduce VSMC vasoconstriction state by stimulating intracellular cAMP level leading to vasodilation suggested a possible synergetic action between ALF and isoproterenol vasorelaxant pathway (Mokkapatti et al., 1998; Xu et al., 2006; Townsend et al., 2012; Ruiz-Medina et al., 2018). The present findings are in accordance with those of other authors which have shown that extract of Thymus linearis Benht significantly reduced the contraction effect of U46619 in endothelium-denuded porcine coronary rings in concentration dependent (Alamgeer et al., 2018). In addition, the extract of this plant has significantly improved the effect of isoproterenol, which indicated a certain implication of the cAMP relaxation pathway that was in line of those of ALF. Likewise, the ALF effect might also be due to other mechanisms of actions such as the PDEs inhibition pathway and confirm our previous study (Belemnaba et al., 2013). In fact, the previous study indicated that ALF endothelium-independent effect might be explained by a stimulation of the K+, Na+/K+-ATPase channels but also via inhibition of PDEs activity.
The inhibition of PDEs fosters the accumulation of second messengers according to their family. Under normal physiological conditions, PDE1 was activated by the Ca2+-calmodulin complex. On the one hand, PDE2 and PDE1 have used cAMP and cGMP as substrates for their activity following a decrease in the two second messengers normally responsible for the vasodilatation of VSM (Lugnier, 2006, 2011; Keravis and Lugnier, 2012; Bobin et al., 2016). On the other hand, the PDE5 have used cGMP as substrate and their inhibition leads specifically to the accumulation of cGMP at the smooth muscle level (Lugnier, 2006). In the present findings, the results have shown that the effects of SNP in the presence of nimodipine (PDE1 specific inhibitor) and EHNA (PDE2 specific inhibitor) were slightly shifted to the left but not significantly different as compared to the control.
These results suggest that, in the presence of EHNA, cGMP accumulated by the action of SNP on sGC reactivates PDE2 present in VSM which degrades this substrate thus reducing the effect of sGC activator. However, nimodipine did not significantly improve the effect of SNP compared to the control and its effect in the presence of EHNA. These results indicate a possible involvement of PDE5 which degrade the cGMP produced by the SNP in the case of PDE2 inhibition by EHNA; insofar as in the presence of DMPPO, this effect was significantly different from the control (Figure 3). Thereby, these results corroborate those of others studies in which, in rat lungs precontracted with U46619, the DMPPO amplifies the vasodilator effects of sodium nitroprusside (Eddahibi et al., 1998).
Furthermore, in the present study, the results showed that the effect of isoproterenol on rings pre-incubated with ALF 3 μg/mL is comparable to those obtained in the presence of nimodipine and DMPPO (Figure 5). On the other hand, this effect in the presence of ALF (10, 30 or 100 μg/mL) is improved and comparable to that obtained with rings pre-incubated with cilostamide (PDE3 inhibitor, Figure 6). Thus, at high concentrations, ALF would also inhibit PDE3 to ensure its vasodilator effect.
Because PDE3 was well known to have a high affinity for cAMP and hydrolyzes cAMP with a rate 10-fold greater than for cGMP hydrolysis despite its capacity to hydrolyze both cAMP and cGMP, this study suggests a possible increase of cAMP and cGMP in VSM after ALF treatment (Beavo, 1995). These results are in line with other studies performed in denuded rat aorta which led to vasodilation and this effect was enhanced by the cilostamide (PDE3 inhibitor) and by the combinaison of SNP plus DMPPO (Delpy et al., 1996; Cui and Green, 2003).
Moreover, the large-conductance calcium-activated potassium channels (BKCa) were an important potassium ion channels in the VSMC membrane. A depolarization of VSMC membrane potential was followed by an increase of BKCa, leading to the increase of the intracellular potassium efflux responsible of membrane hyperpolarization. Since then, L-type-calcium channel opening was reduced and was followed by the intracellular calcium concentration which decreased, responsible to vasodilation (Qian et al., 2017). In this study, results showed that ALF has no significant effect on denuded rings pre-incubated with IBTX indicating that BKCa has not been involved in ALF endothelium independent vasodilation effect.
Likewise, investigation of ALF effect with the KATP channels inhibitor (Glibenclamide), showed that this effect was slightly but not significantly enhanced compared to the control. That result suggests that KATP pathway appears not to be involved in the ALF vasodilation process (Figure 7). The same observation was obtained with the non-selective K+ channel blocker (TEA) claiming that ALF would induce its effect by preferentially inhibited by the PDE1, 3, and 5 activity than K+, KATP and BKCa channels involved
CONCLUSION
The present findings demonstrated that ALF obtained from barks of trunk of A. leiocarpa was able to induce endothelium-independent vasodilation effect in denuded porcine coronary arteries rings pre-contracted with U46619. This effect might be due to a possible decrease in [Ca2+]i and the reduction of the membrane hyperpolarization. In addition, ALF endothelium-independent effect was more due to its capacity to inhibit PDE1, 3 and PDE5 in the VSMC but not the inhibition of the PDE2. Moreover, it also appears that the implication of openned channels (BKCa, K+ and KATP) was too lessier than those of the inhibition of PDEs and the Ca2+-decrease in vessels. Then, the efficacy for acute and chronic usage of ALF needs to be investigated in in vivo model.
ACKNOWLEDGEMENTS
This work was supported by grant from Service de Coopération et d’Action Culturelle de l’Ambassade de France to Belemnaba Lazare. The author thanks Dr Fadima BOCOUM for the technical support.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Alamgeer AC, Chabert P, Lugnier C, Mushtaq MN, Schini-Kerth VB (2018). Mechanisms underlying vasorelaxation induced in the porcine coronary arteries by Thymus linearis, Benth.Journal of Ethnopharmacology 225:211-219. |
|
|
Arbonnier M (2009). Arbres arbustes et lianes des zones sèches d 'Afrique de l'Ouest. Paris, MNHN, Quae 576 p. |
|
|
Basrali F, Kocer G, Ulker KP, Nasircilar US, Sati L, Ozen N, Ozyurt D, Senturk UK (2015). Effect of magnesium supplementation on blood pressure and vascular reactivity in nitric oxide synthase inhibition-induced hypertension model. Clinical and experimental hypertension 37:633-642. |
|
|
Beavo JA (1995). Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiological Reviews 75:725-748. |
|
|
Belemnaba L, Ouedraogo S, Auger C, Chataigneau T, Traore A, Guissou IP, Lugnier C, Schini-Kerth VB, Bucher B (2013). Endothelium-independent and endothelium-dependent vasorelaxation by a dichloromethane fraction from Anogeissus Leiocarpus (DC) Guill. Et Perr. (Combretaceae): possible involvement of cyclic nucleotide phosphodiesterase inhibition. African journal of traditional, complementary, and alternative medicines 10:173-179. |
|
|
Belemnaba L, Ouedraogo S, Nitiema M, Chataigneau T, Guissou IP, Schini-Kerth VB, Bucher B, Auger C (2018). An aqueous extract of the Anogeissus leiocarpus bark (AEAL) induces the endothelium-dependent relaxation of porcine coronary artery rings involving predominantly nitric oxide. Journal of basic and clinical physiology and pharmacology 29:599-608. |
|
|
Bhattacharya B, Williams R, Latif ML, Roberts RE (2005). Role of cytosolic phospholipase A2 in the enhancement of alpha2-adrenoceptor-mediated vasoconstriction by the thromboxane-mimetic U46619 in the porcine isolated ear artery: comparison with vasopressin-enhanced responses. Biochemical Pharmacology 70:1200-1210. |
|
|
Bobin P, Belacel-Ouari M, Bedioune I, Zhang L, Leroy J, Leblais V, Fischmeister R, Vandecasteele G (2016). Cyclic nucleotide phosphodiesterases in heart and vessels: A therapeutic perspective. Archives of cardiovascular diseases 109:431-443. |
|
|
Cam ME, Hazar-Yavuz AN, Yildiz S, Ertas B, Ayaz Adakul B, Taskin T, Alan S, Kabasakal L (2018). The methanolic extract of Thymus praecox subsp. skorpilii var. skorpilii restores glucose homeostasis, ameliorates insulin resistance and improves pancreatic beta-cell function on streptozotocin/nicotinamide-induced type 2 diabetic rats. Journal of Ethnopharmacology 231:29-38. |
|
|
Chakraborti S, Chowdhury A, Kar P, Das P, Shaikh S, Roy S, Chakraborti T (2009). Role of protein kinase C in NADPH oxidase derived O2*(-)-mediated regulation of KV-LVOCC axis under U46619 induced increase in [Ca2+]i in pulmonary smooth muscle cells. Archives of biochemistry and biophysics 487:123-130. |
|
|
Chung JH, Biesiadecki BJ, Ziolo MT, Davis JP, Janssen PM (2016). Myofilament Calcium Sensitivity: Role in Regulation of In vivo Cardiac Contraction and Relaxation. Frontiers in Physiology 7:562. |
|
|
Cogolludo A, Moreno L, Bosca L, Tamargo J, Perez-Vizcaino F (2003). Thromboxane A2-induced inhibition of voltage-gated K+ channels and pulmonary vasoconstriction: role of protein kinase Czeta. Circulation Research 93:656-663. |
|
|
Cui H, Green RD (2003). Regulation of the cAMP-elevating effects of isoproterenol and forskolin in cardiac myocytes by treatments that cause increases in cAMP. Biochemical and Biophysical Research Communications 307:119-126. |
|
|
Delpy E, Coste H, Gouville AC (1996). Effects of cyclic GMP elevation on isoprenaline-induced increase in cyclic AMP and relaxation in rat aortic smooth muscle: role of phosphodiesterase 3. British Journal of Pharmacology 119:471-478. |
|
|
Eddahibi S, Raffestin B, Launay JM, Sitbon M, Adnot S (1998). Effect of dexfenfluramine treatment in rats exposed to acute and chronic hypoxia. American Journal of Respiratory and Critical Care Medicine 157:1111-1119 |
|
|
Fu J, Zhu X, Wang W, Lu H, Zhang Z, Liu T, Xu H, Fu H, Ma S, Luo Y (2017). 1, 6-di-O-caffeoyl-beta-D-glucopyranoside, a natural compound from Callicarpa nudiflora Hook impairs P2Y12 and thromboxane A2 receptor-mediated amplification of platelet activation and aggregation. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology 36:273-282. |
|
|
Gan L, Wang ZH, Zhang H, Zhou X, Zhou H, Sun C, Si J, Zhou R, Ma CJ, Li J (2016). Endothelium-independent vasorelaxant effect of 20(S)-protopanaxadiol on isolated rat thoracic aorta. Acta pharmacologica Sinica 37:1555-1562. |
|
|
Greenberg HZ, Shi J, Jahan KS, Martinucci MC, Gilbert SJ, Vanessa Ho WS, Albert AP (2016). Stimulation of calcium-sensing receptors induces endothelium-dependent vasorelaxations via nitric oxide production and activation of IKCa channels. Vascular pharmacology 80:75-84. |
|
|
Hahnenkamp K, Nollet J, Strumper D, Halene T, Rathman P, Mortier E, Van Aken H, Knapp J, Durieux ME, Hoenemann CW (2004). Bupivacaine inhibits thromboxane A2-induced vasoconstriction in rat thoracic aorta. Anesthesia and analgesia 99:97-102. 10.1213/01.ANE.0000118107.62304.25 |
|
|
Han SZ, Karaki H, Ouchi Y, Akishita M, Orimo H (1995). 17 beta-Estradiol inhibits Ca2+ influx and Ca2+ release induced by thromboxane A2 in porcine coronary artery. Circulation 91:2619-2626. |
|
|
Hanasaki K, Nakano K, Kasai H, Arita H, Ohtani K, Doteuchi M (1988). Specific receptors for thromboxane A2 in cultured vascular smooth muscle cells of rat aorta. Biochemical and Biophysical Research Communications 150:1170-1175. |
|
|
Hirata M, Hayashi Y, Ushikubi F, Yokota Y, Kageyama R, Nakanishi S, Narumiya S (1991). Cloning and expression of cDNA for a human thromboxane A2 receptor. Nature 349:617-620. |
|
|
Hottinger DG, Beebe DS, Kozhimannil T, Prielipp RC, Belani KG (2014). Sodium nitroprusside in 2014: A clinical concepts review. Journal of anaesthesiology and clinical pharmacology 30:462-471. |
|
|
Jia M, Zhou XX, Qin Q, Wang F, Li J, Xu CB, Wang S (2018). Tetrahydroxystilbene glucoside-induced relaxation of the superior mesenteric artery via both endothelium-dependent and endothelium-independent mechanisms. Microvascular research 123:42-49. |
|
|
Kassahun GA, Wijerathne TD, Kim JH, Kim MJ, Seo CS, Shin HK, Lee KP (2017). Psoralea corylifolia extract induces vasodilation in rat arteries through both endothelium-dependent and -independent mechanisms involving inhibition of TRPC3 channel activity and elaboration of prostaglandin. Pharmaceutical biology 55:2136-2144. |
|
|
Keravis T, Lugnier C (2012). Cyclic nucleotide phosphodiesterase (PDE) isozymes as targets of the intracellular signalling network: benefits of PDE inhibitors in various diseases and perspectives for future therapeutic developments. British Journal of Pharmacology 165:1288-1305. |
|
|
Khan S, Khan T, Shah AJ (2018). Total phenolic and flavonoid contents and antihypertensive effect of the crude extract and fractions of Calamintha vulgaris. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology 47:174-183. |
|
|
Konaté K, Kiendrébéogo M, Ouattara M B, Souza A, Lamine-Meda A, Nongasida Y, Barro N, Millogo-Rasolodimby J, Nacoulma OG (2011). Antibacterial Potential of Aqueous Acetone Extracts From Five Medicinal Plants used Traditionally to Treat Infectious Diseases in Burkina Faso. . Current Research Journal of Biological Sciences 3:435-442. |
|
|
Li H, Kim HW, Shin SE, Seo MS, An JR, Jung WK, Ha KS, Han ET, Hong SH, Bang H, Choi IW, Na SH, Park WS (2018). The vasorelaxant effect of antidiabetic drug nateglinide via activation of voltage-dependent K(+) channels in aortic smooth muscle. Cardiovascular therapeutics 36. |
|
|
Lugnier C (2006).Cyclic nucleotide phosphodiesterase (PDE) superfamily: a new target for the development of specific therapeutic agents. Pharmacology and therapeutics 109:366-398. |
|
|
Lugnier C (2011). PDE inhibitors: a new approach to treat metabolic syndrome? Current opinion in pharmacology 11:698-706. |
|
|
Mann A, Banso A, Clifford LC (2008). An antifungal property of crude plant extracts from Anogeissus leiocarpus and Terminalia avicennioides. Tanzania Journal of Health Research 10:34-38. |
|
|
Minuz P, Meneguzzi A, Fumagalli L, Degan M, Calabria S, Ferraro R, Ricci M, Veneri D, Berton G (2018). Calcium-Dependent Src Phosphorylation and Reactive Oxygen Species Generation Are Implicated in the Activation of Human Platelet Induced by Thromboxane A2 Analogs. Frontiers in pharmacology 9:1081. |
|
|
Mokkapatti R, Vyas SJ, Romero GG, Mi Z, Inoue T, Dubey RK, Gillespie DG, Stout AK, Jackson EK (1998).Modulation by angiotensin II of isoproterenol-induced cAMP production in preglomerular microvascular smooth muscle cells from normotensive and genetically hypertensive rats. The Journal of pharmacology and experimental therapeutics 287:223-231. |
|
|
Montalvo D, Perez-Trevino P, Madrazo-Aguirre K, Gonzalez-Mondellini FA, Miranda-Roblero HO, Ramonfaur-Gracia D, Jacobo-Antonio M, Mayorga-Luna M, Gomez-Viquez NL, Garcia N, Altamirano J (2018). Underlying mechanism of the contractile dysfunction in atrophied ventricular myocytes from a murine model of hypothyroidism. Cell calcium 72:26-38. |
|
|
Montfort WR, Wales JA, Weichsel A (2017). Structure and Activation of Soluble Guanylyl Cyclase, the Nitric Oxide Sensor. Antioxidants and redox signaling 26:107-121. |
|
|
Mora MT, Ferrero JM, Romero L, Trenor B (2017). Sensitivity analysis revealing the effect of modulating ionic mechanisms on calcium dynamics in simulated human heart failure. PloS one 12, e0187739. |
|
|
Otsuka AJ, Buoncristiani MR, Howard PK, Flamm J, Johnson C Yamamoto R, Uchida K, Cook C, Ruppert J, Matsuzaki J (1988). The Escherichia coli biotin biosynthetic enzyme sequences predicted from the nucleotide sequence of the bio operon. Journal of Biological Chemistry 263:19577-19585. |
|
|
Ouedraogo V, Kiendrebeogo M (2016). Methanol Extract from Anogeissus leiocarpus (DC) Guill. et Perr. (Combretaceae) Stem Bark Quenches the Quorum Sensing of Pseudomonas aeruginosa PAO1. Medicines 3 p. |
|
|
Qian YF, Wang Y, Tian WW, Wang S, Zhao L, Li L, Ma KT, Si JQ (2017). Effects of RMF on BKCa and Kv channels in basilar arterial smoothmuscle cells of SHR. Molecular medicine reports 16:2620-2626. |
|
|
Rapoport RM, Murad F (1983). Agonist-induced endothelium-dependent relaxation in rat thoracic aorta may be mediated through cGMP. Circulation Research 52:352-357. |
|
|
Ruiz-Medina BE, Cadena-Medina DA, Esparza E, Arrieta AJ, Kirken RA (2018). Isoproterenol-induced beta-2 adrenergic receptor activation negatively regulates interleukin-2 signaling. The Biochemical Journal 475:2907-2923. |
|
|
Sanchez-Recillas A, Rivero-Medina L, Ortiz-Andrade R, Araujo-Leon JA, Flores-Guido JS (2019). Airway smooth muscle relaxant activity of Cordia dodecandra A. DC. mainly by cAMP increase and calcium channel blockade. Journal of Ethnopharmacology 229:280-287. |
|
|
Sinharoy P, Bratz IN, Sinha , Showalter LE, Andrei SR, Damron DS (2017).TRPA1 and TRPV1 contribute to propofol-mediated antagonism of U46619-induced constriction in murine coronary arteries. PloS one 12, e0180106. |
|
|
Somlyo AP, Somlyo AV (2000). Signal transduction by G-proteins, rho-kinase and protein phosphatase to smooth muscle and non-muscle myosin II. The Journal of Physiology 522(2):177-185. |
|
|
Soro D, Kone WM, Bonfoh B, Dro B, Toily KB, Kamanzi K (2013). In vivo anthelmintic activity of Anogeissus leiocarpus Guill & Perr (Combretaceae) against nematodes in naturally infected sheep. Parasitology Research 112:2681-2688. |
|
|
Townsend EA, Sathish V, Thompson MA, Pabelick CM, Prakash YS (2012). Estrogen effects on human airway smooth muscle involve cAMP and protein kinase A. American journal of physiology. Lung cellular and molecular physiology 303:923-928. |
|
|
Vajic UJ, Grujic-Milanovic J, Miloradovic Z, Jovovic D, Ivanov M, Karanovic D, Savikin K, Bugarski B, Mihailovic-Stanojevic N (2018). Urtica dioica L. leaf extract modulates blood pressure and oxidative stress in spontaneously hypertensive rats. Phytomedicine : International Journal of Phytotherapy and Phytopharmacology 46:39-45. |
|
|
Xu H, Lu H, Zhu X, Wang W, Zhang Z, Fu H, Ma S, Luo Y, Fu J (2018). Inhibitory effects of luteolin4'ObetaDglucopyranoside on P2Y12 and thromboxane A2 receptormediated amplification of platelet activation in vitro. International Journal of Molecular Medicine 42:615-624. |
|
|
Xu YC, Yeung DK, Man RY, Leung SW (2006). Kaempferol enhances endothelium-independent and dependent relaxation in the porcine coronary artery. Molecular and Cellular Biochemistry 287:61-67. |
|
|
Zhao Y, Vanhoutte PM, Leung SW (2015). Vascular nitric oxide: Beyond eNOS. Journal of Pharmacological Sciences 129:83-94. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


