Full Length Research Paper

ABSTRACT
Dexamethasone is an anti-inflammatory glucocorticoid that may alter glucose and lipid homeostasis depending on the dose and/or duration of administration. Nigella sativa oil (NSO) can be used as potential modulator of intermediary glucose and lipid metabolism. Herein, the authors evaluated the dose-dependent effects of NSO (0.2 and 0.4 ml/kg body weight) supplementation on plasma glucose, insulin and triglyceride concentration in rats treated with dexamethasone (1 mg/kg body weight) for 5 days. Adult male Wistar rats (150 to 200 g) were housed and treated in the animal holding facility of the Faculty of Basic Medical Sciences, University of Ilorin, Ilorin. Animals were randomly distributed into 4 groups: control (normal saline, 1 ml/kg), DEX (dexamethasone 1 mg/kg), DEX + NSO 0.2 ml/kg and DEX+ NSO 0.4 ml/kg. All drugs were administered intraperitoneally. To assess insulin resistance, an insulin tolerance test was performed in addition to fasting glucose and insulin measurements used in calculating the homeostatic model assessment of insulin resistance index (HOMA-IR). Insulin-induced glucose lowering was improved following NSO supplementation during the insulin tolerance test (p<0.05). Likewise, there was a reduction in fasting hyperinsulinemia (50%; p<0.05) and HOMA-IR index. Dexamethasone induced hypertriglyceridemia which was not significantly reduced following NSO supplementation. Furthermore, NSO had no significant effect on liver alanine aminotransferase enzyme activity in rats treated with dexamethasone. In conclusion, it is shown that 5 days therapy with dexamethasone can be used as a model of insulin resistance and NSO supplementation for 5 consecutive days attenuates hyperglycemia and insulin resistance induced by dexamethasone treatment.
Key word: Dexamethasone, insulin resistance, Nigella sativa oil.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
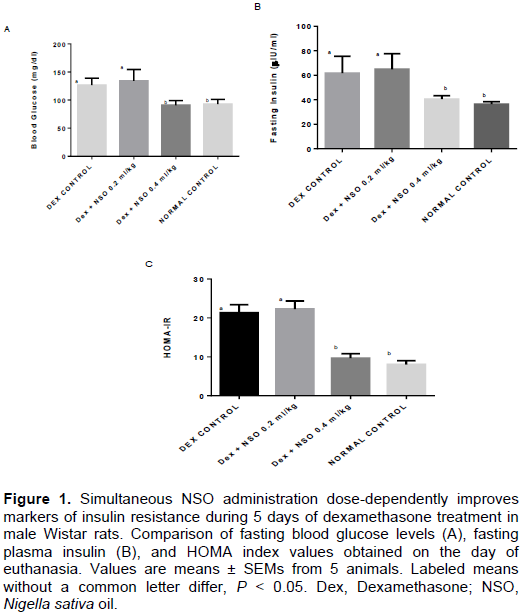


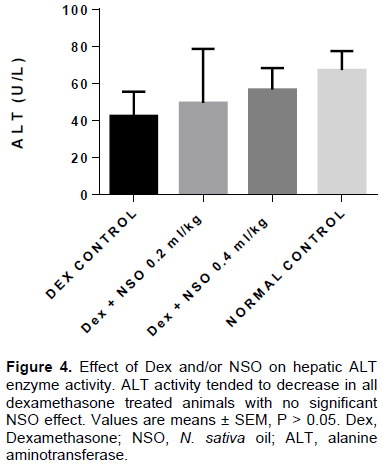
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Ahmad A, Husain A, Mujib M, Khan SA, Najmi AK, Siddique NA, Damanhouri Z, Anwar F (2013). A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. J. Trop. Biomed. 3(5):337-352. |
|
|
Alsaif M (2008). Effect of Nigella sativa oil on impaired glucose tolerance and insulin insensitivity induced by high-fat-diet and turpentine-induced trauma. Pak. J. Biol. Sci. 11(8):1093-1099. |
|
|
American Veterinary Medical Association (AVMA) (2013). AVMA Guidelines For The Euthanasia of Animals. |
|
|
Barbosa AM, Francisco PDC, Motta K, Chagas TR, dos Santos C, Rafacho A, Nunes EA (2015). Fish oil supplementation attenuates changes in plasma lipids caused by dexamethasone treatment in rats. Appl. Physiol. Nutr. Metab. 41(4):382-390. |
|
|
Benhaddou-Andaloussi Ali, Martineau L, Vuong T, Meddah B, Madiraju P, Settaf A, Haddad PS (2011). The In Vivo Antidiabetic Activity of Nigella sativa Is Mediated through Activation of the AMPK Pathway and Increased Muscle Glut4 Content. Evid. Based Complement. Altern. Med. 2011:1-9. |
|
|
Bönisch C, Irmler M, Brachthäuser L, Neff F, Bamberger M, Marschall S, de Angelis H, Becjkers J (2016). Dexamethasone treatment alters insulin, leptin, and adiponectin levels in male mice as observed in DIO but does not lead to alterations of metabolic phenotypes in the offspring. Mamm. Genome 27:17-28 |
|
|
Bonora E, Kiechl S, Willeit J, Oberhollenzer F, Egger G, Targher G, Alberiche M, Bonadonna R, Muggeo M (1998). Prevalence of Insulin Resistance in Metabolic Disorders The Bruneck Study. Diabetes 47:1643-1649. |
|
|
Bose SK, Hutson I, Harris CA (2016). Hepatic glucocorticoid receptor plays a greater role than adipose GR in metabolic syndrome despite renal compensation. Endocrinology 157(12):4943-4960. |
|
|
Cain DW, Cidlowski JA (2015). Specificity and sensitivity of glucocorticoid signaling in health and disease. Best Pract. Res. Clin. Endocrinol. Metab. 29(4):545-556. |
|
|
Darakhshan S, Pour AB, Colagar AH, Sisakhtnezhad S (2015). Thymoquinone and its therapeutic potentials. Pharm. Res. 95:138-158. |
|
|
Fararh KM, Atoji Y, Shimizu Y, Shiina T, Nikami H, Takewaki T (2004). Mechanisms of the hypoglycaemic and immunopotentiating effects of Nigella sativa L. oil in streptozotocin-induced diabetic hamsters. Res. Vet. Sci. 77(2):123-129. |
|
|
Fararh KM, Shimizu Y, Shiina T, Nikami H, Ghanem MM, Takewaki T (2005). Thymoquinone reduces hepatic glucose production in diabetic hamsters. Res. Vet. Sci. 79(3):219-223. |
|
|
Ferris H, Kahn RC (2012). New mechanisms of glucocorticoid-induced insulin resistance: make no bones about it. J. Clin. Invest. 122(11):3854-3857. |
|
|
Gao X, Li B, Jiang H, Liu F, Xu D, Liu Z (2007). Dioscorea opposita reverses dexamethasone induced insulin resistance. Fitoterapia 78:12-15. |
|
|
Harad S, Smith RM, Hu DQ, Jarett L (1996). Dexamethasone inhibits insulin binding to insulin-degrading enzyme and cytosolic insulin-binding protein p82. Biochem. Biophys. Res. Commun. 218(1):154-158. |
|
|
Imam A, Ajao MS, Ajibola MI, Amin A, Abdulmajeed WI, Lawal AZ, Alli-Oluwafuyi A, Akinola OB, Oyewopo AO, Olajide OJ, Adana MY (2016). Black seed oil ameliorated scopolamine-induced memory dysfunction and cortico-hippocampal neural alterations in male Wistar rats. Bull. Fac. Pharm. Cairo Univ. 54(1):49-57. |
|
|
Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, Tobe K (2006). Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Invest. 116(7):1784-1792. |
|
|
Kanter M, Coskun O, Korkmaz A, Oter S (2004). Effects of Nigella sativa on oxidative Stress and beta-Cell Damage in Streptozotocin-Induced Diabetic Rats. Anat. Rec. 279A:685-691. |
|
|
King G, Park K, Li, Q (2015). Selective Insulin Resistance and the Development of Cardiovascular Diseases in Diabetes: The 2015 Edwin Bierman Award Lecture. Diabetes 65:1462-71. |
|
|
Levy JC., Mathews RR, Hermans MP (1998). Correct homeostasis model assessment (HOMA) evaluation uses the computer program. Diabetes care 21(12):2191-2192. |
|
|
Martin BC, Warram JH, Krowleski AS, Soeldner JS, Kahn CR, Martin BC, Bergman RNB (1992). Role of glucose and insulin resistance in development of type 2 diabetes mellitus: results of a 25-year follow-up study. Lancet 2(8825):925-929. |
|
|
Okada T, Liew CW, Hu Jiang, Hinault C, Micheal DM, Crutzfeldt J, Yin C, Holzenberger M, Stoffel M, Kulkarni RN (2007). Insulin receptors in beta-cells are critical for islet compensatory growth response to insulin resistance. Proc. Natl. Acad. Sci. 104(21):8977-8982. |
|
|
Pari L, Sankaranarayanan C (2009). Beneficial effects of thymoquinone on hepatic key enzymes in streptozotocin–nicotinamide induced diabetic rats. Life Sci. 85(23):830-834. |
|
|
Pasieka AM, Rafacho A (2016). Impact of Glucocorticoid Excess on Glucose Tolerance: Clinical and Preclinical Evidence. Metabolites 6(3):24. |
|
|
Patel R, Patel M, Tsai R, Lin V, Bookout AL, Zhang Y, Magomedova L, Li T, Chan JF, Budd C, Mangelsdorf DJ (2011). LXRβ is required for glucocorticoid-induced hyperglycemia and hepatosteatosis in mice. J. Clin. Invest. 121(1):431-41. |
|
|
Phanse MA, Manohar J, Abbulu PK (2014). Effect of Thespesia populnea Linn on dexamethasone induced insulin resistance in mice. Afr. J. Pharm. Pharmacol. 8(16):429-432. |
|
|
Protzek A, Costa-Júnior JM, Rezende LF, Santos GJ, Araujo TG, Vettorazzi JF, Ortis F, Carneiro EM, Rafacho M, Boschero AC (2016). Hyperinsulinemia caused by dexamethasone treatment is associated with reduced insulin clearance and lower hepatic activity of insulin-degrading enzyme. J. Steroid Biochem. Mol. Biol. 155 (Pt A):1-8. |
|
|
Rafacho A, Roma LP, Taboga AC, Boschero A, Bosquiero JR (2007). Dexamethasone-induced insulin resistance is associated with increased connexin 36 mRNA and protein expression in pancreatic rat islets. Can. J. Physiol. Pharmacol. 85(5):536-545. |
|
|
Rafacho A, Giozzet V, Boschero A, Bosquiero J (2008). Functional Alterations in Endocrine Pancreas of Rats With Different Degrees of Dexamethasone-Induced Insulin Resistance. Pancreas 36:284-293. |
|
|
Rafacho A, Goncalves-Neto LM, Santos-Silva JC, Alonso-Magdalena P, Beatriz M, Taboga SR, Carneiro E, Boschero AC, Nadal A, Quesada I (2014). Pancreatic Alpha-Cell Dysfunction Contributes to the Disruption of Glucose Homeostasis and Compensatory Insulin Hypersecretion in Glucocorticoid-Treated Rats. PLoS One 9(4):e93531. |
|
|
Reitman S, Frankel S (1957). A colorimetric method for determination of serum glutamate oxaloacetate and glutamic pyruvate transaminase. Am. J. Clin. Pathol. 28:56-58. |
|
|
Sarwar A, Latif Z (2015). GC-MS characterization and antibacterial activity evaluation of Nigella sativa oil against diverse strains of Salmonella. Nat. Prod. Res. 29(5):447-451. |
|
|
Wajs A, Bonikowski R, Kalemba D (2008). Composition of essential oil from seeds of Nigella sativa L. cultivated in Poland. Flavour Fragr. J. 23:126-132. |
|
|
Woo SL, Xu H, Li H, Zhao Y, Hu X, Zhao J, Guo X, Guo T, Botchlett R, Qi T, Pei Y (2014). Metformin ameliorates hepatic steatosis and inflammation without altering adipose phenotype in diet-induced obesity. PloS one 9(3):e91111. |
|
|
Yoon CJ, Puigserver P, Chen G, Donovan J, Wu Z, Rhee James, Adelmant G, Stafford J, Kahn RC, Graner DK, Newgard CB, Spiegelman BM (2001). Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1. Nature 413:131-138. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


