For sorghum hybrid development, assessment of parental lines is a pre-requisite. However, information on heterotic performance and combing ability of Ethiopian elite sorghum lines is inadequate. ANOVA revealed mean squares had signifying substantial amount of variability amongst genotypes for most traits. Males 72, 81 and 99 were the best performing parents for yield and related traits. Hybrids, 106 x 94, 106 x 90, 106 x 102, 107 x 99 and 107 x 105 were found maximum heterotic hybrids for yield as compare to check. The estimates of variance of combining ability revealed that σ2gca was found inferior to σ2sca for all traits except plant height and number of heads. The σ2gca/σ2sca ratio revealed preponderance of supremacy gene action for most traits. The degree of dominance was found greater than unity for entire traits except plant height. The estimations of parental GCA effects showed that female 106 and males 79, 96, 94 and 81 were good general combiners for yield and related traits. Based on perse performance, heterotic response, combining ability and nature of gene action for yield and related traits, female parent 106 and male parents 94, 102 and 90 were found most performed. Those parental lines were grouped into dualistic heterotic groups based on their SCA and GCA results.
Genetic variability and genotype performance across sites
The recorded data on different agronomic traits were subjected to analysis of variance to assure the differences among the experimental genotypes. The combined analysis over locations was done for all traits based on their homogeneity test across locations. The combined analysis of variance for phenological and grain yield and yield-related parameters is presented in Table 1.
The mean square value of combined analysis of variance for grain yield was 2.3 which was highly significantly different (p<0.01). Similarly, the mean square value of inbred parents was 2.4 indicating highly significant difference at p<0.01 between them (Table 1). This shows that the hybrids and the inbred lines have an inherent genetic variability which could be useful to make selection and genetic advancement. The mean squares due to total genotypes for combined analysis were highly significant different at p <0.01 for all the studied traits (Table 1). The analysis of variance revealed significant genotypic effect for all the traits. This provides evidence of the presence of enough genetic variability among females, males, and hybrids and allows further assessment of combining ability analysis.
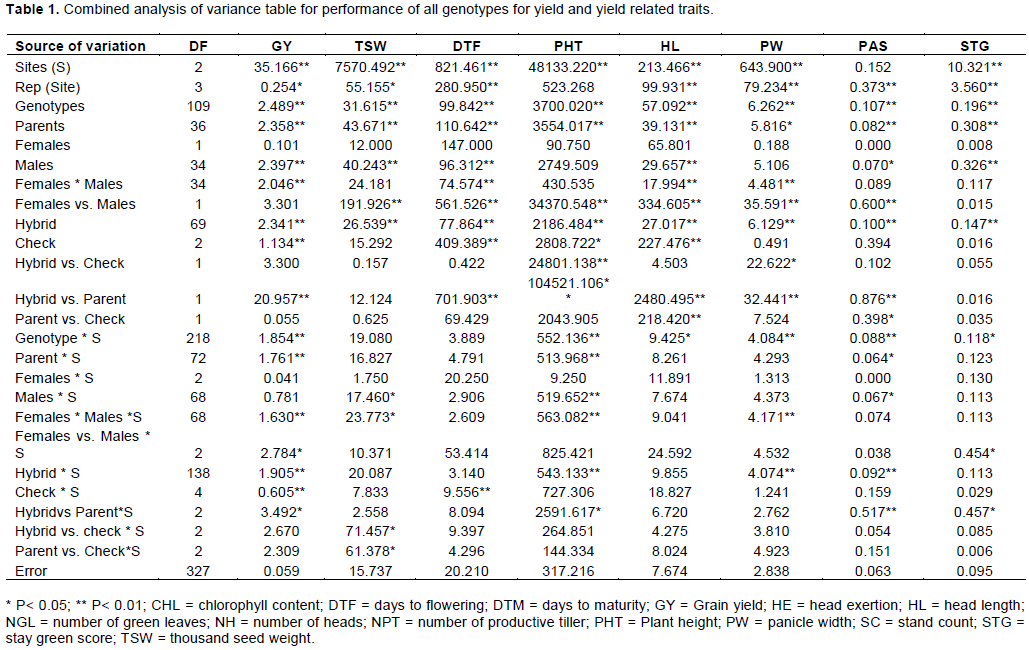
The genotype x environment interaction showed significant difference for eight traits but there was no significant variation for TSW, DTF and HE (Table 1). The combined analysis also revealed significant differences among parents for all the traits studied. The interaction between test hybrids with environment exhibited significant difference for all measured traits except head exertion. The interaction between parents and the test sites was found to be significant for GY, PHT, DTM, NH, PAS and HE. This suggests that the inbred lines used for the cross development responded differently in different environments. However, the female parents were not significantly different for all the traits measured except for plant height and panicle width. The significant difference in phenological traits (plant height and panicle width) agreed with the established fact that drought hardy crops like sorghum can change their phenological growth in response to the dominant climatic conditions (Tadesse et al., 2008).
The interactions between the male and female parents were found to be significantly different for eight of the measured traits (GY, DTF, NGL, CHL, DTM, HL, PW and NPT) but showed insignificant difference for the rest of traits. This interaction indicated that male and female parents differed significantly in respect to the main and majority of traits studied in the present investigation. This significant difference of male and female interaction revealed the presence of genetic recombination among the inbred lines which will give rise to better specific combining ability that gives the chance to identify superior hybrids for the desired trait.
The analysis of variance further revealed that hybrids differed significantly for all the traits as their mean square values were highly significant; this point out the existence of considerable genetic variability among the hybrids for all the traits that were studied. Mean squares due to hybrid vs. parent were significant for all traits except TSW and STG. This suggests the presence of heterosis for these traits as parents and hybrids were found to be significantly different (Table 1).
From the combined ANOVA, it was observed that mean squares due to male parents was significant and higher than that of the female parents for all the traits except PHT and PW. This shows the existence of better diversity among the male parents than the female parents for most of the traits measured. The interaction of females with their respected testing sites was insignificant for all the traits. This reveals that the female parents did not respond to the environments differently (Table 1). The interaction of females * males was significantly different for all traits except TSW, PHT, NH, PAS, HE and STG. Similarly, the interaction with the sites was significantly different for GY, TSW, PHT, DTM, PW and NH. Among all the genotypes, checks were significantly different only for GY and DTF.
In general, the significant mean squares of female and male component revealed the presence of additive variance, whereas the non-significant means square revealed the presence of non-additive or dominance variance (Table 1).
Hybrid means performance and magnitude of heterosis across sites
Mean performance of the hybrids
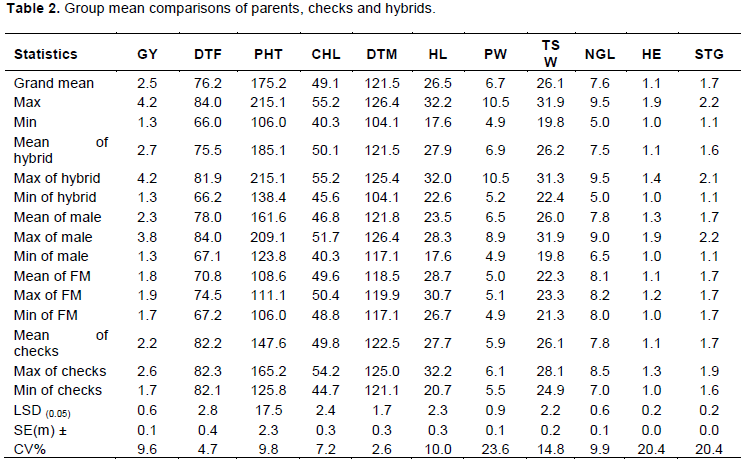
Hybrid as compared to their parents and checks had comparable advantages in mean performance. The mean of grain yield for hybrids ranged from 1.3 to 4.2 t/ha. Among the grand mean of all the genotypes, the maximum GY was attained by a hybrid cross of 106 x 24 (4.2 t/ha) followed by hybrid combination of 106x32 (4.1 t/ha), 106x20 (4.1 t/ha), 107x29 (4.0 t/ha) and 107x35 (4.0 t/ha) with an average value of 2.7 t/ha which had high mean value than the grand mean, mean of checks and mean of parents. Similarly, inbred line parents had mean value of ranging from 1.3 to 3.8 t/ha (Table 2). From the top better genotypes, genotype 24 and genotype 44 were statistically different while genotype 24, 20, 32, 64, 70, 22 and 25 were not statistically different for their GY mean values. But the bottom worst and top better genotypes were statistically different for the mean value.
Magnitude of heterosis for hybrids
The phenomenon of heterosis has provided the most important genetic tool in improving yield of crop plants. Identification of specific parental combination capable of producing the highest level of heterotic effects in F1 is of immense value for commercial exploitation of heterosis. Percent heterosis for yield and yield related traits was computed over better parents, mid parent and better check. The magnitude of heterosis varied from trait to trait and even from cross to cross. Heterosis over the mid parent, better parent and better check of the hybrids among 35 inbred lines and two female lines are summarized in Table 3. There was significant variation for levels of heterosis among the parental genotypes. Mean performance of the hybrids and heterosis for grain yield ranged from -43.5 to 160.0 (%) across sites. The best parent heterosis (BPH) for grain yield ranged from -55.2 to 129.4(%) with the mean value of 32.3%. The magnitude of heterosis of the hybrids in relation to the better check hybrid (BCH) ranged from -50.0 to 61.5% with the mean value of 3.0% in combination of all testing sites. In general, 84.3% of the crosses exhibited positive MPH and the rest negative heterosis whereas 25.7% of crosses shows >50% of MPH (Table 3). The BPH analysis for grain yield also was done based on the higher performed parent of Hybrids. 68.6% of hybrids attain positive heterosis for BPH and there also 31.4% of hybrids show negative heterosis from high parent of among the corresponding male and female parents. In general, 10 hybrids get greater than 51% of BPH.
The magnitude of heterosis varied from cross to cross and trait to trait. For a specific trait considerable high heterotic effects were observed in certain crosses and low in others, which revealed that nature of gene action varied with the genetic makeup of parents. The results indicated that both positive and negative heterosis was observed for these studied traits. For days to heterosis was desirable but for rest of the traits positive heterosis is desirable. In some cases, for the lowland areas negative heterosis might be desirable for plant height in order to shorten days to flowering and physiological maturity as well as to get lodging free flowering, days to maturity days to emergence etc. negative different. Contrariwise, the comparison of parents vs. hybrids. In other way, positive heterosis for plant height might be desirable for those areas having long maturity period that is in highland and also intermediate sorghum growing areas so as to increase its biomass.
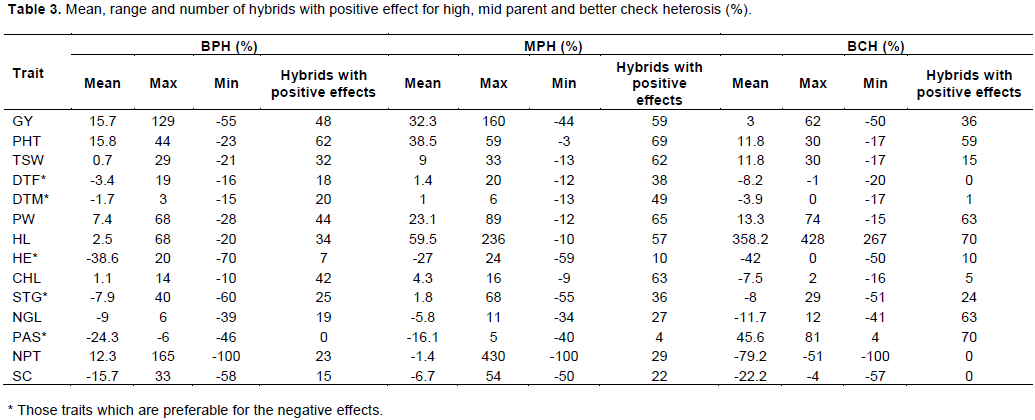
From the resultant 70 crosses, 37 potential hybrids exhibited significant standard heterosis performance over three checks (ESH-1, ESH-4 and Argiti) in desirable direction for grain yield and among these hybrids 31 hybrids shows significant superiority from the better check. There are also differences for standard heterosis (SH) for yield which was calculated based on relative mean (%), better check hybrid means (%). Most of the crosses displayed positive SH of better check heterosis (BCH) for relative to the highest performing check (better check) where the rest crosses exhibited negative BCH (Table 3).
Similarly, for PHT, there was a significant variability level of hybrid vigour among the studied hybrid parents. In this case MPH ranged from 59.4 to -2.5%. Among the hybrids, 69 of them exhibited positive heterosis while the rest 1 hybrid showed negative heterosis. For the case of BPH heterosis ranged from -23.1 to 44.1% and only 8 hybrids show negative heterosis while the rest are positive heterosis and all positive heterosis ranged between 3 and 44.1%. Like this, there also significant difference of standard heterosis which was done based on mean of better check and this ranged from -16.6 to 29.9% where 11 hybrids exhibited negative BCH and the rest 69 hybrids showed positive BCH. The detailed analysis for heterosis regarding MPH, BPH and BCH is listed out in Table 3.
Significantly, negative BCH for DTF was observed in all the crosses with the range of maximum and minimum heterosis of -0.5 for a cross of 107×76 to -19.5% for 106×86, respectively. Thus, it appeared that the earliest male parent 86 had contributed for earliness, while the late male parent 76 contributed for lateness in comparison of better check heterosis. Among nine significant mid parent heterosis for DTF only four hybrids show significant negative mid parent heterosis while five of them are positively significant different from zero. In this case, negative heterosis was preferable whether it was significant or not for better hybrid vigour to flower earlier than of the better check. BPH ranged from -15.9 by a cross of 106 × 90 to 19.3% by the cross 107 x 86 which was the negative values are goodly selected for better hybrid performance to flower early as compare to that of better parent and the highest positive BPH was not preferable in this condition.
In general, lower value negative heterosis was preferable than higher value of positive heterosis for DTF and DTM. In another way higher value of positive heterosis was good for the rest traits such as GY, TSW, NPT, HL, HE, PW and the like. Heterosis in grain yield for male parent 67 x female line 107 was 0 (Zero). This indicates that as there was additive gene action, and this implies also there is no heterosis. For this case F1 hybrid should be the midpoint or average of both parents.
The per se performance of hybrids for grain yield and its components was in general connected to the heterotic effects. This brought out that selection of hybrids either on the basis of per se performance or on the basis of magnitude of heterotic effects would also be unfailing. This can be supported by yield advantage over OPV standard check. In this case 11 hybrid exhibited higher yield advantage over OPV check. These are across of 106 x 94, 106 x 90, 106 x 102, 107 x 99, 107 x 105, 106 x 95, 107 x 79, 106 x 81, 106 x 92, 107 x 81 and 107 x 94 have yield advantage of ranged from 42 to 75%.
Combining ability analysis
Analysis of variance for combining ability
Combining ability analysis was done following the significance value of genotypes from the general analysis of variance table in the fashion of line x Tester analysis method (Table 1). The nature and magnitude of estimates of genetic variance can deliver an idea about the relative role of fixable (explainable or cumulative) and non-fixable (unexplainable) gene effects in the inheritance of traits. This in turn can help us in identifying suitable parents for hybrid breeding as well as type of breeding method. The genetic variances were estimated from the analysis of variances for combining ability for all traits as suggested by Singh and Chaudhary (1977). The variations among the hybrids were further partitioned into genetic components attributable to GCA and SCA based on the method suggested by Kempthorne (1957). Joint analysis of variance over locations for combining ability indicated that variances due to females was significant for all traits except CHL and TSW, thereby revealing significant contribution of females towards combining ability for these traits. Mean squares due to males were also significant for all the studied traits except HE, and this shows the greatness of good parental selection for this hybrid cross formation which was found highly variable and this was an indication of getting good pollinator lines for hybrid breeding program (sorghum hybrid grain production). The single degree of freedom comparison for parent vs. hybrid, which indicates average heterosis, was significant for all the traits except for DTM and TSW; this clearly suggested considerable amount of average heterosis in the hybrids (Table 4). This also reflected the presence of adequate genetic variability in the experimental material. Similar findings has been reported for average heterosis by comparing parent vs. hybrid in single degree of freedom for 50 hybrids derived from ten female and five male sorghum lines (Kumar et al., 2017).
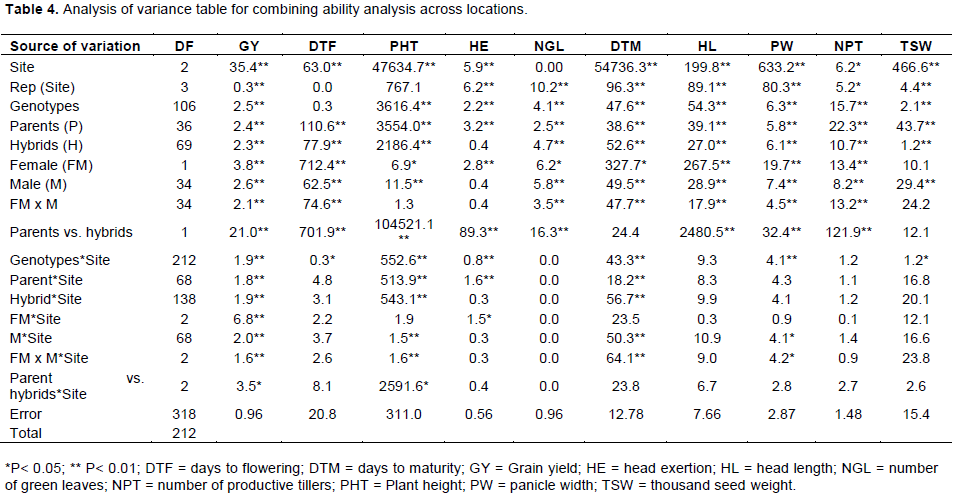
Combining ability analysis indicated significant GCA mean squares for all measured traits except CHL and TSW for females and HE for males. All traits except PHT, HE, SC and TSW for the female * male interaction were significantly different. In other way, those traits with mean squares due to female * male interaction were significant indicates the presence of genetic variability for SCA among the crosses. These observations further support comparisons of parents vs. hybrids for all traits in a single degree of freedom. Comparing parents with hybrids all measured traits except DTM and TSW was significantly hybrids with site is non-significant except GY and PHT.
The interaction of GCA of females (mean square of females) and site was significantly different only for GY and HE. Similarly, the interaction of site with GCA of males and female x male * site interaction is significantly different for only GY, PHT, DTM and PW and this indicates susceptibility for SCA among the hybrids of the testing sites for these studied traits.
Proportional contribution GCA of females and SCA of males and male*female interaction
In general, the proportional contribution of females, males and their interaction to the total variance showed that males played an important role in the traits indicating predominant male lines influence for these traits (Table 5). It also suggests that in the hybrid sorghum breeding more efforts should be paid to the selection of parental lines.
The highest contributions for grain yield were males and the lowest were the female lines. The proportional contribution of females, males and their interactions (Female x Male) to total variances showed that males played an important role in the traits such as GY, PHT, HE, NGL, CHL, DTM, HL, PW, TSW and SC indicating predominant male influence for these traits (Table 5); and the male to female interaction showed important role for only number of productive tillers. The smaller contribution of interactions of the female x male than males, indicating higher estimates of variances due to general combining ability that is additive gene action among the males used.
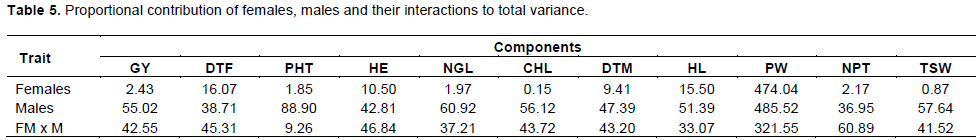
Contribution of interactions of female x male was higher than males for DTF, NPT, indicating higher estimates of SCA variances for interaction. For the same case male lines play important role for days to flowering in this cross being female and male interaction contributing more percentage than the rest (Females and males) did. Similarly, contribution of interactions of female x male was higher than female lines for all traits, indicating higher estimates of SCA variances for interaction. Again, the proportional contribution of males was observed to be higher than that of female x male interactions thus lower estimates of SCA variances.
In general, the proportional contribution of males, females and their interaction to the total variance showed that males played an important role towards the traits indicating predominant male lines influence for traits (Table 5). It also suggested that in the sorghum hybrid breeding more efforts should be paid to the selection of male lines.
General combining ability effects (GCA) for females and males inbred lines
The general combining ability effect of females and males for all measured traits were presented in Table 6. Significant positive and negative GCA effects were observed for all traits. For two Females CHL, NGL and TSW were not found significant and PHT not significant for ATX-623 (female line 106) and the rest significant for both females. Among 35 male lines 12 of them were non-significant for GY. Similarly, for DTF both the negative and positive values showed negative values were selected even if the values were non-significant. In this study; 27 male lines were non-significant for their GCA, but, 17 males were negatively combiner and the rest were positively combiner. The results showed negative and significant GCA effects for days to flowering in the female line (106) and this line induce earliness for the cross which it involves. Male parents including (80, 82 and 92), suggesting the contribution of these parents for earliness in crosses they were involved. Meanwhile, positive and significant GCA effects were observed in 75, 76, 78, 79 and 103 for the same trait. These parents greatly induced lateness in their crosses. The former parents were low general combiners (that is, significant and negative GCA effects). Thirty-two of the crosses expressed negative heterosis over mid parent involving at least one of these parents. The male parent, 86, 78, 76 and 85 greatly induced lateness in their crosses. Out of the two crosses involving male line 80, one of them (female line 106 x male line 80) exhibited significantly negative heterosis over mid and better parent. The lowest significant negative GCA effect (-12.1) was exhibited by male line 80, whereas the highest positive GCA (19.7) was observed in male line 86.
GCA for shorter plant stature (-42.6) was exhibited by male line 75 and the longer one (25.76) was exhibited by male line 105. In the case of GY, the range of maximum and minimum GCA was 1.13 and -0.93 respectively. In this case, male line 75 introduce dwarfness (shortness) for the crosses; it was involved and male line 105 was induced for tallness.
The effects of GCA for male lines revealed that 19 male lines were positively combiner and the rest were negatively combiner. The highest positive GCA value was male line 94 followed by male line 81 and 90 and the lowest negative GCA value was exhibited by male line 74, 88 and 86. Moreover, male line 94 induced high yielding gene effect for the crosses in which it was involved while male line 74 induced low yielding gene effect for those crosses made by male line 74. Among the thirty-five male lines, male line 94, 81, 90 and 92 could be selected for good general combiner for high yield.
Plant height is one of the determinant factors of yield and now a days breeding is to improve grain yield in the same way with biomass. On the contrarily plant height and grain yield is negatively correlated. That means both traits can’t improve at the same time. In this study, 12 males were negatively combiner for plant height and the rest 23 males were positively combiner. The 12 male lines could be selected for reducing plant height while the 23 male lines could be selected for increasing plant height. The highest positive GCA effects were contributed by male lines 105, 104, 84 and 82 respectively. Even though, male lines 75, 80, 93 and 102 were the lowest negative combiner for plant height. Among the highest positive combiner male lines, 81, 102, 99, 94, 90, 105, 104 and 84 were positive significant combiner for grain yield and male line 72, 71, 78, 82, 89 and 91 were significant negative combiner for grain yield.
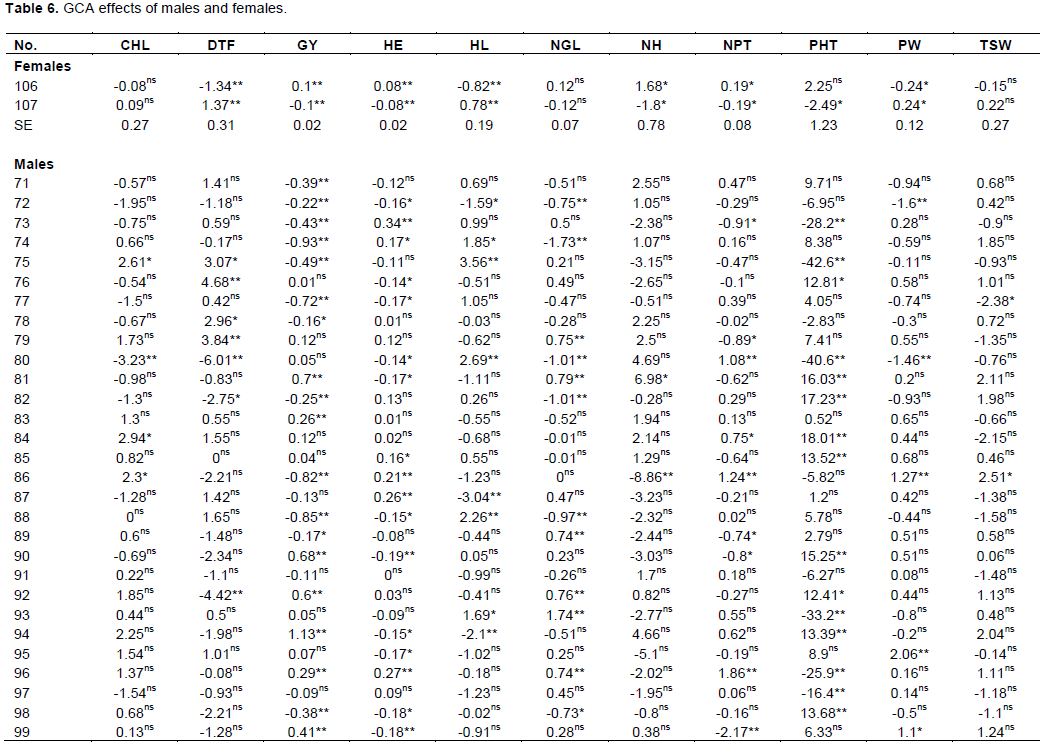
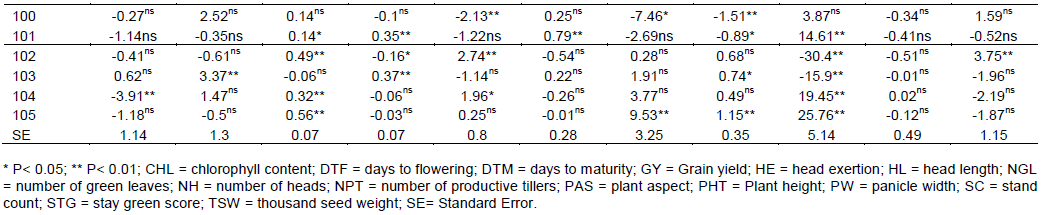
For the increment performance of hybrids compared to their inbred parents plant height is one of the driving factors for the wider utilization in sorghum production (Mindaye, 2015). The plant height is much better for the need of biomass especially in forage breeding. 20 inbred male parents were significant for the plant height and the rest were not significant, particularly, among 12 of them exhibited negatively GCA effects for the hybrid output. These parents with significant positive GCA effects were good combiners in increasing tallness, while those with significant negative GCA effects were good combiners in decreasing plant height and may be desirable in areas of lodging problem is facing.
The results revealed that male parents were grouped under negatively and positively combiner in the case of days to flowering. Effects of general combining ability analysis revealed that for days to 50% flowering, 16 male parents were positively combined, and the rest 19 males were negatively combined. Only 9 male inbred lines exhibited significant combined effects for days to flowering, and this is an indication that, those with significant negative GCA effects were good combiners in decreasing days to flowering that means to exploit hybrids for their early flowering and those with positively combined effects were better for hybrid exploitation of increasing photo period before flowering. Moreover, parents with positively combined effects greatly induced lateness in their crosses and those with negative effects induce earliness to their respective F1 hybrids.
Panicle length and panicle width were positively correlated with grain yield increment. Male parents exhibited positive combined effects and the rest 20 male lines were negatively combined. So, improving panicle width through heterosis can help to exploit the hybrid grain yield.
In order to get high yielder genetic material for sorghum, panicle width should have wide diameter (width), and among the current study genetic materials, 19 male parents were found positively combined with the respective female lines and the remaining 16 males showed negative general combining ability effects. Male line 95, 86, 99 and 85 could be selected for good positive combiner for wide panicle width. Contrarily, male lines 72, 80, 71 and 82 were the narrowest and poor combiner to increase panicle width for better yield improvement.
Specific combining ability (SCA) effects
The results for SCA estimates detected both negative and positive values for a male line crossed with the two females with equal SCA values in magnitude and opposite in direction but varied among traits that were studied. This may be due to the two females, which had equal combining ability in magnitude but opposite in direction. Similar work was reported in the case of maize using 16 female lines and 2 male lines and the SCA effects was equal in magnitude but opposite in sign (Ejigu et al., 2017).
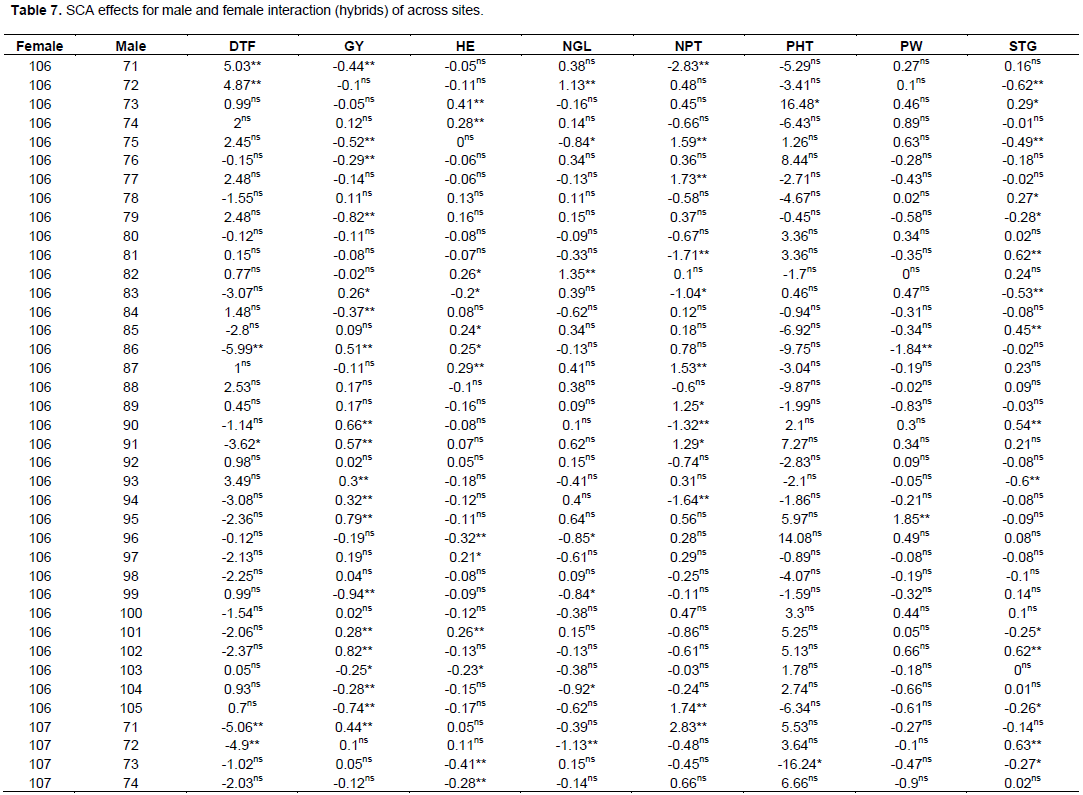
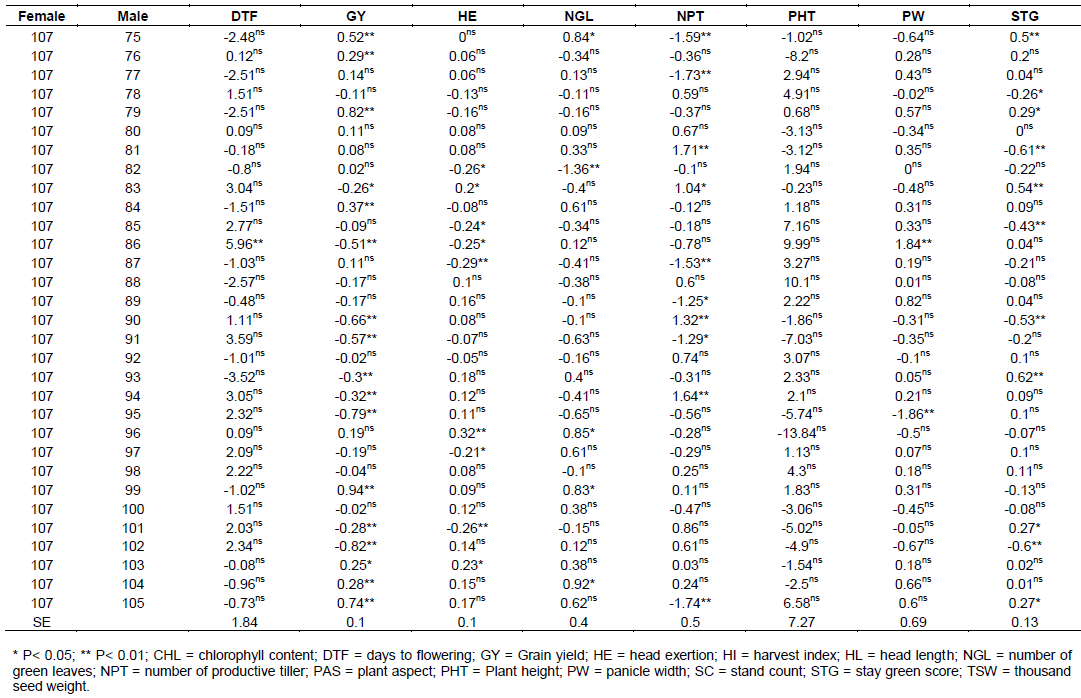
The estimate of SCA effects for the 70 different hybrids in respect of the traits studied are presented in Table 7. Hybrids evaluated in this study revealed considerable variation in SCA effects in all yield and yield-related traits. It was observed that some crosses involved good general combined parents which produced hybrids, with poor specific combining ability for a given trait example yield, indicating parents with high GCA effects might not always give hybrids with high SCA effects. The possible explanation is that both females used in the hybrid may have the same gene controlling effect to the trait(s) studied and not able to take advantage of any additive gene action. Regarding days to flowering, only six hybrid combinations were found highly significant (p<0.01) and one hybrid cross was found significant (p<0.05) probability level. These male parents induced earliness for the crosses where they were involved. But, among these significant hybrids for SCA effects only four (106 x 86, 106 x 91, 107 x 71 and 107 x 72) hybrids were negatively combiner and the rest were positively combiner for this trait.
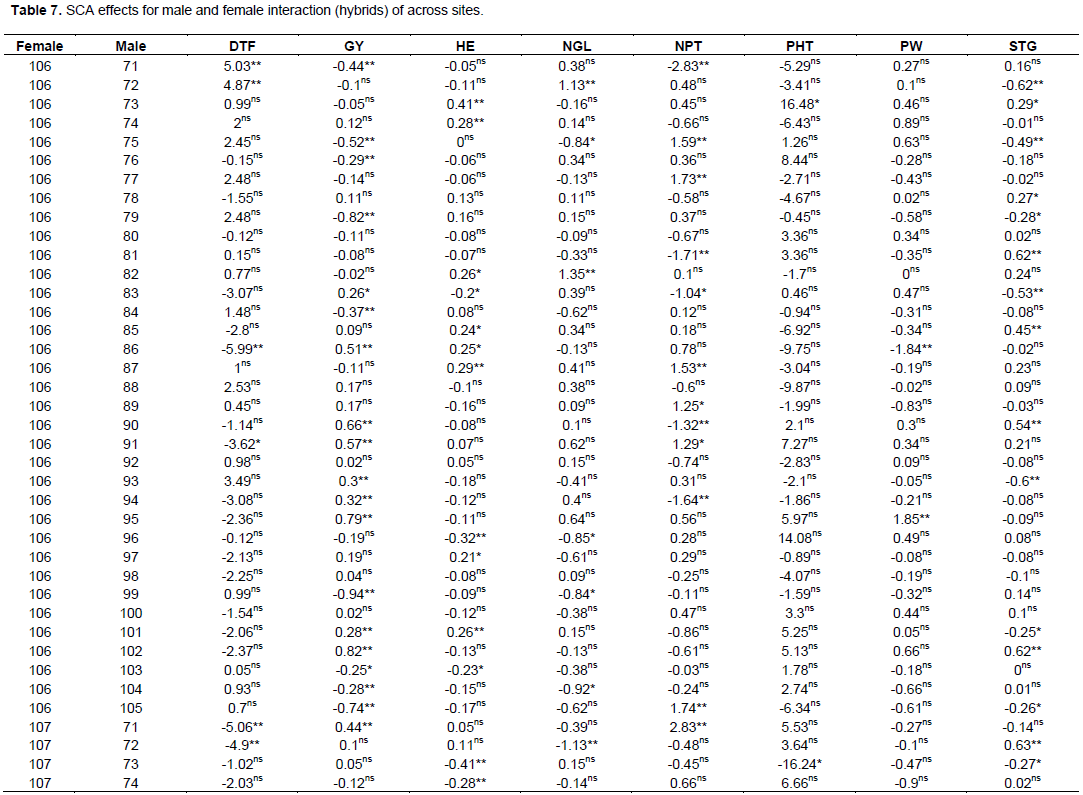
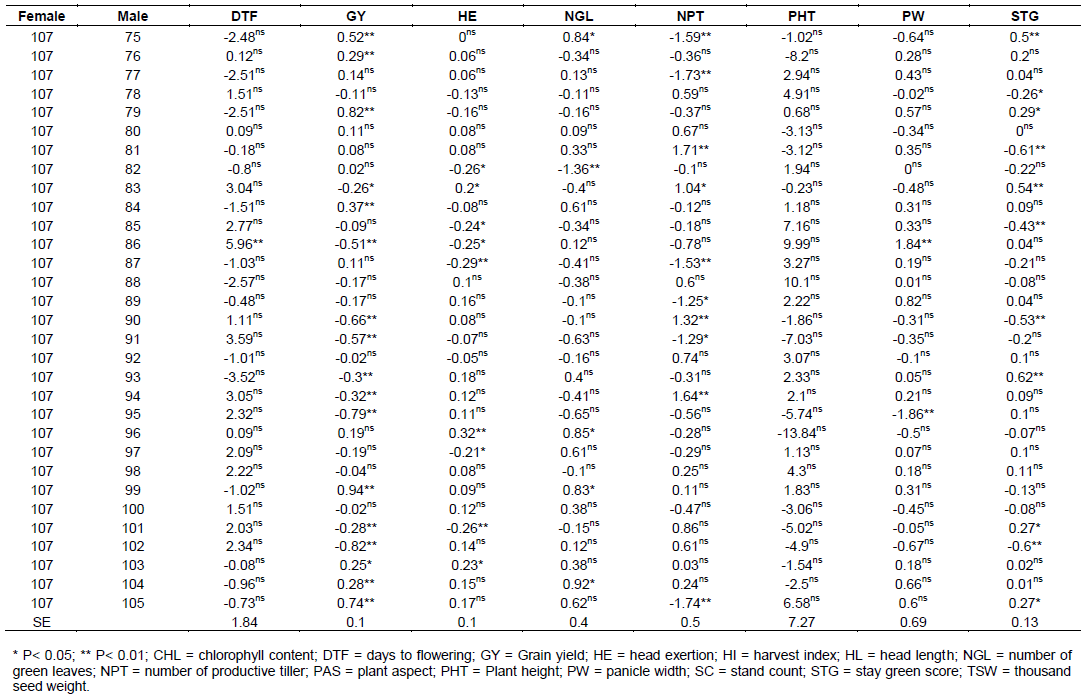
For days to maturity, only five cross combinations were found to be significant and among these only two were found highly significant (p<0.01) and three of them were significant (p<0.05) probability level. SCA estimates for days to 50% flowering and days to physiological maturity showed both negative and positive SCA effects. Negative SCA estimates for these traits indicated that the crosses took fewer days to 50% flowering and physiological maturity. On the contrary, crosses, which had positive SCA estimates for days to 50% flowering and physiological maturity indicated hybrids with delay in days to 50% flowering and maturity. The current investigation was similar to previous reports in maize using 16 female lines with 2 male lines (Ejigu et al., 2017).
Crosses of female 106 and 107 with male 71, 75, 76, 79, 84, 86, 90, 91, 93, 94, 95, 99, 101, 102, 104 and 105 was highly significant (p<0.01), followed by cross of the same female with male lines 83 and 103 which showed significant at (p<0.05) probability level for grain yield (GY). The range was from -0.94 to 0.94 for cross of male line 99 with 106 and 107 female lines, respectively.
For grain yield among all 70 crosses, a cross of same gene controlling effect to the trait(s) studied and not able to take advantage of any additive gene action. Regarding days to flowering, only six hybrid combinations were found highly significant (p<0.01) and one hybrid cross was found significant (p<0.05) probability level. These male parents induced earliness for the crosses where they were involved. But, among these significant hybrids for SCA effects only four (106 x 86, 106 x 91, 107 x 71 and 107 x 72) hybrids were negatively combiner and the rest were positively combiner for this trait.
For days to maturity, only five cross combinations were found to be significant and among these only two were found highly significant (p<0.01) and three of them were significant (p<0.05) probability level. SCA estimates for days to 50% flowering and days to physiological maturity showed both negative and positive SCA effects. Negative SCA estimates for these traits indicated that the crosses took fewer days to 50% flowering and physiological maturity. On the contrary, crosses, which had positive SCA estimates for days to 50% flowering and physiological maturity indicated hybrids with delay in days to 50% flowering and maturity. The current investigation was similar to previous reports in maize using 16 female lines with 2 male lines ((Ejigu et al., 2017).
Crosses of female 106 and 107 with male 71, 75, 76, 79, 84, 86, 90, 91, 93, 94, 95, 99, 101, 102, 104 and 105 was highly significant (p<0.01), followed by cross of the same female with male lines 83 and 103 which showed significant at (p<0.05) probability level for grain yield (GY). The range was from -0.94 to 0.94 for cross of male line 99 with 106 and 107 female lines, respectively.
For grain yield among all 70 crosses, a cross of 107x99, 106x102, 107x79, 106x95, 107x105, 106x90, 106x91, 107x75 and 106x86 revealed the highest positive and highly significant SCA effects. On the other hand, crosses such as, 107x77, 106x88, 106x89 and other few have non-significant positive SCA effects. These positive significant and non-significant SAC affected parents indicates that inbred lines involved in these crosses were genetically divergent, and hence could be regarded to be from different heterotic groups. Cross of 106 x 75, 107 x 91, 107 x 90, 106 x 105, 107x95, 106x79, 107x102, 106x99 and some of others showed lowest significant negative SCA effects for this trait, indicating that these crosses were poor in specific combiners for grain yield. Among all crosses that showed the highest significant positive SCA effects, a cross of 107 x 79 and 107 x 75 were a combination of poor female with poor male line in their respected GCA effects. On the contrary, only 2 hybrids 106 x 102 and 106 x 90 were found from a cross combination of good general combiner of female and male. The rest were a cross of poor and good parental general combiner. This showed that, the crosses performed better than what would be expected from the GCA effects of their respective parents. Therefore, these crosses could be selected for their specific combining ability for higher grain yield. When high yielding specific combinations are desired, especially in hybrid sorghum development, SCA effects could help in the selection of parental material for maximum exploitation of heterosis.
Crosses with positive and higher SCA values are desirable for the improvement of productivity of sorghum hybrid grain yield by exploiting maximum heterosis. On the contrary, crosses with negative SCA values are undesirable for sorghum grain yield. However, to get the best SCA, results may not necessarily be from crosses between two good general combiners. From this study, it was found that the combination of a parent with negative and positive GCA value resulted in a hybrid with highly significant positive SCA values in some cases. For example, the combination of a parent with negative GCA female parent and positive GCA male parents gave positive SCA values. Such as across of female line 107 (-0.1) with male line 76 (0.01), male 79 (0.12), male line 84 (0.41), male line 99 (0.41), male line 104 (0.32) and male line 105 (0.56), resulted in positive SCA values of 0.29, 0.82, 0.37, 0.94, 0.28 and 0.74 respectively. The combination of parents with positive and negative GCA values resulting in positive SCA values is also reported from previous investigators (Ejigu, 2017). On the other hand, there were combinations of both parents with positive GCA values, which, resulted in hybrids with negative SCA values, such as female line 106 x male line 76, female line 106 x male line 80, female line 106 x male line 81, female line 106 x male line 84, female line 106 x male line 96 and female line 106 x male line 99, female line 106 x male line 104 and female line 106 x Male line 105. The reverse is also true when two parents with negative GCA values were crossed and gave hybrids with positive SCA values such as across of female line 107 with male lines 71, 72, 73, 75, 77, 82, 87 and 103. Therefore, from this study it was found out that the good specific combiners for different traits involved parents with high x high, high x low, low x low general combining ability evaluations.
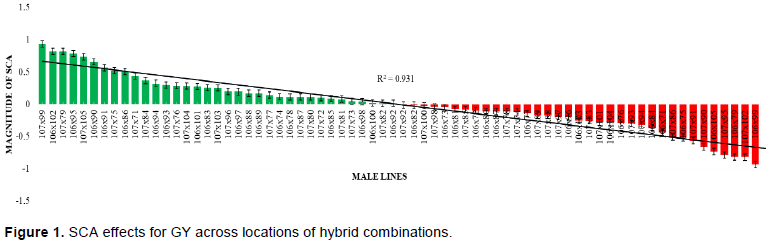
Among the hybrids, 35 showed positive effects for SCA and the rest showed negative SCA effects. Based on their SCA magnitude the three highest ranks from the top list was a cross of female line 107 with male line 99, 106 x 102 and 107 x 79 respectively. On the other hand, there were also three lowest ranked from the 70 hybrids based on their SCA effects. Those crosses were 106 x 79, 107 x 102 and 107 x 99 and these were the negative reverse of the above former mentioned crosses (Figure 1).
SCA of grain yield derived from female line 106 exhibited positive SCA effect when crossed with male lines 74, 78, 83, 85, 86, 88, 89, 90, 91, 92, 93, 94, 95, 97, 98, 100, 101 and 102. However, hybrid derived from female line 107 exhibited negative SCA effect for those above-mentioned male lines but, equal in magnitude with SCA effects of hybrids derived from female lines 106.
Similarly, days to 50% flowering and plant height also exhibited the same SCA effects. The negative SCA effect is preferred for early maturing traits and this happens due to the rule of the summation of SCA effect is zero since two female lines were used to derive the hybrids.
Gene effects and estimates of variance components for combining ability
Variances of male, females and their interaction (female * male)
Estimates of variance indicated the higher SCA variances than the GCA variances for all the studied traits show that predominance of non-additive gene action in the inheritance of these traits and vice versa. The variance of females was found smaller than that of the male lines for all studied traits except NPT which was higher for females than males. The total variance components of parents were contributed by the male lines for cross (hybrid) variances. That is the total hybrid (male * female interaction) variances is its due to male’s variation. So, it can be said that selection for male parents for this hybrid production was successful and the next parental selection for hybrid sorghum grain production should take an account of parents for their variance which can put contribution towards the hybrid production (Table 8).
Variances of combining ability effect
Estimates of gca variance (σ2gca), SCA variance(σ2sca) and unity ratio (Table 8). The estimates of variance due to combining ability revealed that σ2gca was lower than σ2sca for all the traits except plant height and number of heads. However, the ratio of σ2gca / σ2sca revealed the preponderance of dominance gene action for all traits except for plant height and number of heads, where additive gene action was more with σ2gca /σ2sca ratio being more than unity.
Generally, the SCA variances (σ2sca) were higher than GCA variance (σ2gca) for almost all the traits except plant height. The ratio of σ2gca/σ2sca was less than one for almost all the considered traits except plant height indicating prevalence of non-additive gene action (dominance and epistasis). Similar investigation was reported for forage sorghum hybrids (Dehinwal et al., 2017). The magnitude of GCA/SCA variance ratio for plant height was specifically sizable, indicating the predominance of additive gene action; however, the specific effects were also highly significant, suggesting the involvement of non-additive effects in controlling this trait. For days to 50% flowering the preponderance of SCA effects (dominance gene effect) was higher than the additive gene effects and this indicates that hybrids are earlier than their parental effect. Similar investigation was done for 7 lines and 8 testers of 28 forage hybrids in relation to days to 50% flowering (Mohammed, 2009). For all traits, SCA variances found higher preponderance of additive gene action heterosis breeding will be effective. Contrarily, for those traits such as plant height and number of heads’ GCA, variances are higher than SCA variances (Table 3).
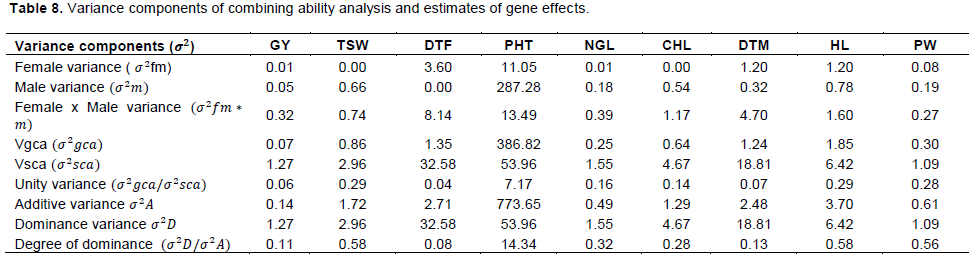
The degree of dominance (σ2D/σ2A) was found greater than one for all the traits except plant height, indicating the over dominance behavior of interacting alleles. Since over dominance geneaction is involved for inheritance of grain yield, heterosis breeding would be the most effective approach to improve the trait. The significance of mean square for female x male provides a direct test of significance of dominance variance, σ2D while significance of σ2A is provided by significance of females and males mean squares (Nduwumuremyi et al., 2013) (Table 8). The results revealed that for plant height, stay green score, and other few, the additive genetic effects were more pronounced than non-additive effects, and the general combining ability variance was higher than specific combining ability. This result suggests that the inheritance of these traits was mainly controlled by additive genes and selection of parents should be more important in breeding practice.
Estimates of gene action
Detail estimates of additive variance (σ2A), dominance variance (σ2D) and average degree of dominance (σ2D /σ2A) on the basis of pooled analysis over sites (Table 8). The estimates for components of genetic variance, that is, σ2A and σ2D were computed from the variance of combining ability calculated on the basis of the covariance of half sib and full sib as suggested by Singh and Chaudhary (1977). Both additive and non-additive gene actions are expected to be important in the expression of the studied traits, with the preponderance of additive gene actions for plant height and stay green and non-additive actions for all traits except plant height and stay green (Table 8).
Estimation of components of variance (additive variance from GCA and dominance variance from SCA variance), and subsequent estimation of average degree of dominance (σ2D/σ2A) was more than unity (one) for most of the studied traits during pooled analysis. This further indicates the preponderance of dominance gene action for most of the traits except plant height, number of heads and stand count where additive gene action seems to be more important (Figure 2) and this agrees with similar findings in forage sorghum (Kumar et al., 2017). For those traits like PHT, NH, SC and STG with degree of dominance was found greater than unity (one) in both the additive and non-additive gene actions which were responsible for inheritance of grain yield and its components in sorghum hybrid grain production.
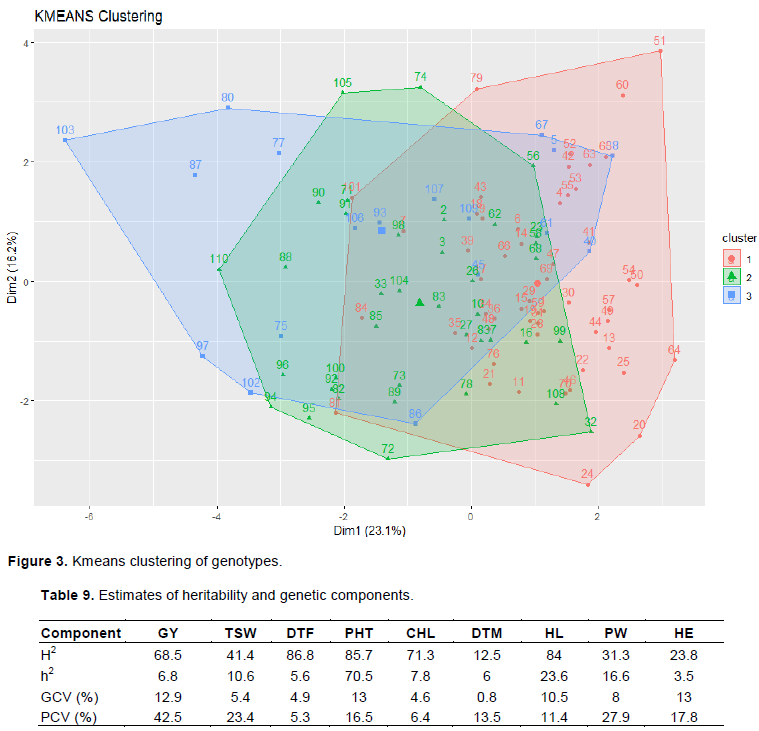
Clustering analysis of experimental genotypes
Estimation of cluster likelihood was calculated based on five independent runs (R software [Version 5.1]) for a variable number of clusters, from K = 2 to 10. K = 3 was chosen due to their low variation of probability values and repetitive clustering. Clustering analysis based on the genetic dissimilarity grouped the 110 sorghum genotypes into 3 distinct clusters. It was observed that cluster I had maximum number of genotypes followed by cluster II and III with 59, 29 and 22 genotypes respectively.
In cluster III, all genotypes are parental genotypes except genotype 45 and 61. In cluster I, most of the grouped genotypes are also parents and some are hybrids (crosses). Both female lines are grouped under cluster III and this indicates that both female lines are similar in their performance. In this case, the dissimilarity range is found between 16.2 to 23.1%. Cluster III had 22 genotypes reflecting narrow genetic diversity amongst them. The narrow genetic diversity may be attributed to similarity in the base material from which they have been evolved. Therefore, it can conclude that parental lines were more variable than their hybrids derived from them since hybrids are concentrated more in cluster I while parents are distributed in clusters II and III (Figure 3).
Heritability and variances of genetic components
The extent to which variation in yield components are responsible for differences in yield among various cultivars, depends on heritable and non-heritable components. While coefficients of variation measure the magnitude of variability present in a population estimates of heritability. The estimates of PCV and GCV were classified as low (< 10%), medium (10-20%), and high (> 20%) (Sivasubramanian and Madhavamenon, 1973). The results revealed considerable phenotypic and genotypic variances among all the genotypes for the traits under consideration for this study. In most traits a large portion of the phenotypic variance was accounted for the genetic component and the contributions of genetic variance to phenotypic variance were found high (Table 9). The estimates of PCV were high for all studied traits ranging from 5.3 of days of flowering to 64.1 of number of productive tillers except for number of productive tillers and number of green leaves which are equal in their GCV and PCV. The PCV values were higher than GCV values for all the traits can reflect the influence of environment on the expression of traits. High PCV values were recorded for GY, PW, NPT and TSW while DTF and NGL were found low. For the case of GCV only for NP is found high value while for GY, PHT, HL, NH and NGL were found medium. All the other traits showed moderate PCV and GCV values. For those traits like grain yield, panicle width, head length, number of productive tiller and number of green leaves in which genotypic variance is higher than error variance, genetic variance is more important, and selection can be done if possible. Similarly, for trait which is GCV is higher indicates that there were low environmental effects on these traits.
Heritability estimates (broad-sense) for yield and its components were done following Singh and Chaudhary (1979). They were categorized using the criteria of Robinson and Comstock (1949): 0-30% = low; 31-60% = moderate; > 60% = high. High heritability estimate was recorded for all traits while for days to maturity was found low and for panicle width, thousand seed weight and number of heads were found moderately heritable. This indicates that these traits are highly heritable, and it would give the best picture of genetic advance to be expected from selection.
Heterotic grouping
Two heterotic grouping methods were used to assign male parents into different groups, including SCA effects for grain yield (Pswarayi and Vivek, 2008) and heterotic grouping based on GCA of multiple traits (HGCAMT) method (Badu-Apraku et al., 2013). Dendrogram was constructed for the groupings based on HGCAMT. Using Ward’s method based on Euclidean distances obtained from GCA for all traits. Classification by these two methods showed similar but not identical trends. As of two female lines (106 and 107) were used in this cross, the principle of SCA effects method is as follows: Male parents showing negative SCA effects when crossed with 106 and exhibiting positive SCA effects with 107 were classified into heterotic Group A. Male lines showing positive SCA effects with 107 and negative effects with 106 were assigned into heterotic Group B, similar result was reported by Akata et al. (2017) for heterotic grouping of 19 male lines crossed with two female lines into 4 groups based on their SCA effects. The SCA effects of grain yield method classified those sorghum elite lines into two heterotic groups (Table 10). Based on this heterotic group, 16 elite lines were found under group A with negative SCA effects for hybrids derived from female line 106 and the rest 19 elite lines were laid under group B which had positive SCA effects for cross of female line 107. Grouping of inbred lines based on their GCA effects of multiple traits should give a better, more probable and practical heterotic group of the lines since GCA dealings the additive gene effects for each trait. Dendrogram based on HGCAMT method grouped sorghum inbred lines into two heterotic groups (Figure 4). In group I, twenty inbred lines were grouped together and in group II fifteen lines were found. Inbred lines of both groups had negative and positive significant GCA effect for grain yield and flowering time; while almost all group I individuals were lateness lines. The highest positive highly significant five lines were being found under group I while the other lowest found in group II for grain yield.
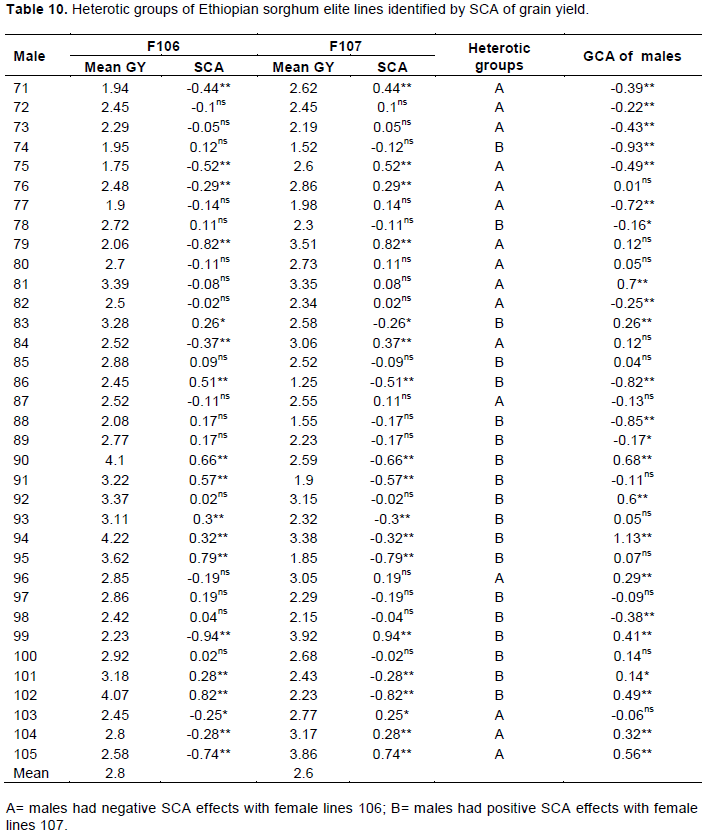
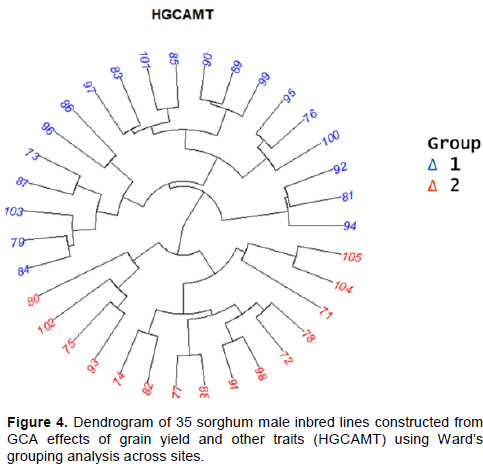
Eight of lines were grouped under group II in both of grouping methods while other eight lines also were grouped under group I use these two grouping methods. The HGCAMT method provides information related to plant performance and general combining ability of multiple traits and can be supported by correlation pattern between per se performance and general combining ability. Furthermore, while HGCAMT tended to put most of the inbred lines with positive GCA effect for grain yield into group I and the ones with negative values into group II, the SCA method puts most of the inbred lines in to group A.
Examination of the results of the classification of the sorghum inbreds based on the SCA effects of yield and HGCAMT, fifteen inbreds were grouped similarly by both grouping methods (SCA and HGCAMT). It is striking that the assortment of the sorghum inbreds in an earlier study using the SCA method (Akata et al., 2017) and HGCAMT method used by (Badu-Apraku et al., 2013) showed close correspondence with the classification by the HGCAMT and SCA methods used in the present study.