Full Length Research Paper

ABSTRACT
Eighteen rice genotypes from Sudan and South Sudan and two genotypes (FR13A tolerant and IR42 sensitive checks) from the International Rice Research Institute were screened for complete submergence at seedling stage in a protected house. Most of the genotypes showed zero survival, except genotypes [NBGA (2.08%), Banban (3.92%), Masury1 (5.56%), Pipanfary Red2 (11.11%) and Sommboy (14.58%)]. Sensitive genotypes registered 2 to 3 folds reduction in root length. Mean soluble sugar concentration in roots increased by 41% and decreased by 57.3 and 80.5% in stem and leaves, respectively. Mean starch concentration was reduced by 13.8% in roots and 77.7% in stems; whereas it increased by 115% in leaves for all genotypes. Genotypes with some degree of tolerance exhibited higher soluble sugar and lower starch concentrations. Submergence reduced chlorophyll a, chlorophyll b and total chlorophyll (a+b) by 83.0, 83.3 and 82.7%, respectively. Genotypes that showed survival after submergence maintained higher values of chlorophyll a, b and (a+b) as compared to other genotypes. Genotypes tolerant to submergence such as Pipanfary Red2 and Sommboy had higher soluble sugar and starch concentrations and chlorophyll content and could be developed to be grown under rain-fed lowland flooded areas of Sudan and South Sudan.
Key words: rice; submergence; seedling stage; starch; soluble sugar; chlorophyll.
INTRODUCTION
Rice (Oryza sativa L.) production in Sudan has a great potential due to the availability of land, water resources as well as a large diversity in agro-climatic zones. Low land rice has been cultivated as the major activity and a key source of employment for rural population at traditional areas of its production (White Nile state in Sudan and low land swamps in South Sudan). In these areas the crop is cultivated as irrigated and flood ecosystem (Gasim et al., 2018). Low yields of 0.98 to 1.2 t ha-1 were produced. One of the major problems associated with this low yield is crop establishment as farmers use direct seeding, a simplified cultivation mode widely used all over the world (Sun et al., 2019). The seeds and young plants can be damaged by drought conditions before flood arrives in addition to weed competition.
Sudden flooding, as a result of global climate change, where large volume of water enters the field in a short time can lead to a high rate of seedling death because they face low oxygen dependent metabolic activity for proper utilization of carbohydrates (Su et al., 2022), and consequently low yield. Moreover, the depth and time of water in which the crop is grown and water conditions such as turbulence (Debarbata and Jijnasa, 2021; Chen et al., 2018), traditional varieties together with low inputs supply, poor drainage, poor maintenance of developed lowland swamps and poor soil management, are among the most important factors for low rice yield (Osman et al., 2012).
Tolerant rice genotypes seedlings normally have more total sugars as compared to seedlings of sensitive genotypes, which are used during submergence to supply energy for growth and maintenance metabolism. Thus initially higher content of soluble and insoluble sugars can be regarded as an advantage for submergence tolerance (Sayani et al., 2017).
Enhanced tolerance of flooding during germination and early seedling growth could help improve crop establishment and promote more widespread adoption of direct seeding. Not much research on submergence screening tolerance has been done on low land rice in Sudan. The objective of this research was to screen some Sudanese rice genotypes in response to complete submergence to develop submergence tolerant high yielding cultivars for sustainable traditional lowland rice production in Sudan.
MATERIALS AND METHODS
Planting material, experiment lay out and screening for complete submergence seedling tolerance
Eighteen Sudanese and South Sudanese rice genotypes seven from Aweil, South Sudan (NBGS1, NBGS2, NBGS3, NBGA, BG400-1, BG90-2, Banban) and eleven from Sudan (Masury 1, Masury 2, Pipanfary, Pipanfary Red 1, Pipanfary Red 2, JaingArri Red, Sommboy, Comarwa, Painjla, Tagmizedo, BactingArri,), which were never tested for submergence tolerance, as well as two cultivars from International Rice Research Institute (IRRI), namely FR13A as tolerant cultivar and IR42 intolerant checks, were used. The experiment was conducted in a protected house in a concrete tank at IRRI.
The seeds were first soaked in water then the floating ones were removed, to assure the germination of the whole healthy seeds. They were incubated in the oven at 33°C for 24 h. Water was removed from the seeds and seeds were left for pre-germination at 33°C for 48 h.
The following day, the well pre-germinated seeds were selected and sown 0.5 cm below the soil surface in trays. The soil was mixed with 3 g Solophos fertilizer, 3 g Muriate of Potash and 6 g Ammonium Sulfate before sowing. Three rows per accession and 20 seeds per row were sown based on the experiment layout, using randomized complete block design with three replications, and covered with another layer of sieved garden soil. The pre-germinated seeds were left to grow under normal conditions for 21 days with regular watering using tab water and continuous weeding. After 21 days of normal growth, the number of seedlings of the first row was counted for seedling survival and the second row was used for sampling of chlorophyll (leaves) and carbohydrates (leaves, stems and roots) for the determination of soluble sugar and starch content. The third row was used for shoot and root length. Thereafter, the entire setup of seedling trays for submergence were submerged in the concrete tank at around 11:00 to 2:00 pm so as to allow the plants to photosynthesize before the treatment and the water depth was 80 cm above the soil surface.
The floodwater temperatures and other floodwater conditions (dissolved O2, light penetration and pH) were monitored daily in the morning (7:00 to 8:00) and in the afternoon (13:00 to 14:00) at various depths during submergence days using oxygen meter, light meter and pH meter, respectively. The seedlings were submerged for 11 days, and then de-submerged in the 12th day in the afternoon, when light intensity was low, and shoot and root lengths were measured. Samples for chlorophyll and carbohydrates determination were harvested as before submergence, for the submerged seedlings and the control.
Then the seedlings for assessing survival were left for another 21 days growth recovery under normal conditions with normal watering. Thereafter, the survived seedlings per entry were counted and the survival percentage was determined as follows:
.png)
Analysis of soluble sugar
Subsamples of about 200 mg each of the plant material (roots, stems and leaves) were freeze dried in liquid nitrogen, ground and extracted using 80 % ethanol (v/v). The sugar extract was cooled to room temperature and root sample extracts were diluted to 15 mL because the concentration of sugar in the root is low, while stem and leaf sample extracts were diluted to 25 mL. The residues were dried at 70°C for 24 h prior to starch assay.
For colorimetric determination of soluble sugar, 0.3 mL of the sugar extract was added to the test tube and mixed with 3.0 mL anthrone reagent followed by measurement of absorbance read at 620 nm (Fales, 1951).
Determination of starch
The residue previously dried at 70°C, which remained after soluble sugar extraction, was oven dried and used for starch analysis using the method described by Setter and Laureles (1996). The starch was then boiled for 3 h while vortex was mixed thoroughly every 20 min. The root samples were diluted to 15mL, while stem and leaf samples were diluted to 25mL. The starch hydrolysate (standard) was diluted to 50mL with distilled water. Absorbance was read at 450 nm against a blank sample (reference) after the procedure of Kunst et al. (1988).
Determination of chlorophyll
The samples of one leaf from each accession were first flash-frozen in liquid nitrogen (N2) immediately after bagging and stored at -80°C. Then, they were fully freeze-dried for five days. Chlorophyll concentration was determined following the method of Mackinney (1941) in acetone extracts. Readings were carried out using spectrophotometer machine (SHIMADZU CORP UV 1800) and absorbance was recorded at four wavelengths of 470, 645, 646 and 663 nm. The concentration of chlorophyll a, chlorophyll b and total chlorophyll (a+b) were then determined based on the equation outlined in Lichtenthaler and Wellburn (1983).
Statistical analysis
Computer program (R-STAR, 2013) was used for statistical analysis of the data. The data were analyzed according to the standard statistical procedure as described by Gomez and Gomez (1984) for complete randomized block design.
RESULTS AND DISCUSSION
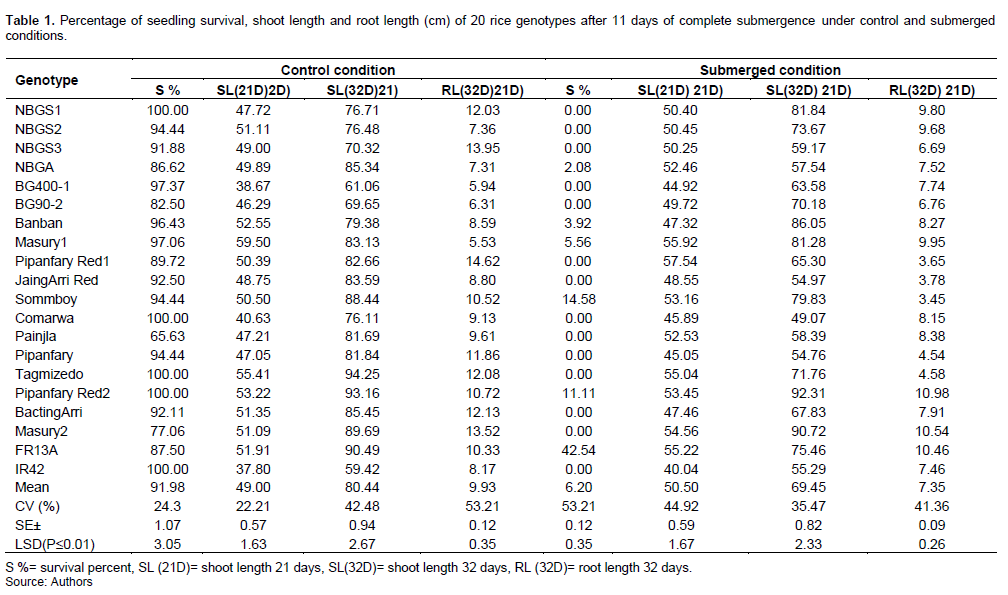
Screening for complete submergence seedling tolerance of 20 rice genotypes under control and complete submerged conditions
Submergence is a major environmental constraint to rice production throughout the world resulting in substantial yield losses and approximately one-fourth of the global rice crops are grown in lowlands that are prone to seasonal flooding (Debarbata and Jijnasa, 2021).
In the present study, statistical analysis for complete submergence tolerance of 20 rice genotypes showed highly significant (P≥0.01) differences between the genotypes in shoot length and root length measured after 21 days and 32 days under control and submerged conditions; while the result for percent seedling survival showed highly significant differences between the genotypes under submerged condition (Table 1). Complete submergence tolerant (FR13A) and sensitive genotypes (IR42 plus the rest of genotypes) had similar seedling survival under control condition; whereas under submerged condition the seedlings survival was greatly reduced by 96.2% for both tolerant and intolerant genotypes; most of the intolerant genotypes approached zero survival, except for genotypes NBGA (2.08%), Banban (3.92%), Masury1 (5.56%), Pipanfary Red2 (11.11%), and Sommboy (14.58%) in comparison to FR13A (42.54%). This suggests their tolerance under rain-fed lowland flooded areas.
Similar results were reported by Su et al. (2022) who found more than 78.1% survival in the rice seeds of strongly submergence-tolerant rice genotype as compared to only 56% survival for submergence-sensitive genotypes when treated by long-term submergence. This high reduction in genotypes survival under complete submergence might be due to the fact that plant survival is affected by depth and extent of submergence (Rachmawati et al., 2019).
Under control condition, the highest (59.50cm) shoot length measured 21 days after seeding was revealed by the genotype Masury1, which was greater than that of FR13A (51.91cm); whereas the least (38.67 cm) shoot length was obtained for the genotype BG400-1 which was more or less the same as that of IR42 (37.80cm). Moreover, the highest (94.25cm) shoot length measured 32 days after seedling growth under control condition was revealed by the genotype Tagmizedo, and was greater than that of FR13A (90.49cm) and the lowest (61.06 cm) shoot length was exhibited by the genotype BG400-1, which was greater than that of IR42 (59.42cm). The highest (14.62cm) root length measured 32 days after seedling growth in control condition was attained by the genotype Pipanfary Red1 longer than FR13A (10.33cm) and the lowest (5.53cm) root length was obtained for Masury1 less than that of IR42 (8.17cm) (Table 1).
Under the submerged condition, shoot length measured after 21 days seedling growth was increased by 3.06%. The highest (55.92 cm) shoot length was exhibited by the genotype Masury1, which was more or less the same (55.22cm) as FR13A and the lowest (44.92 cm) shoot length was registered by the genotype BG400, which was less (40.04cm) than that of IR42 (Table 1). These results are confirmed by Pramanik et al. (2016) who found that most of the rice genotypes showed enhanced shoot elongation in response to submergence. Also Nurrahma et al. (2021a) reported significant increase in shoot length by more than 34.3% after 6 days of submergence than the control and 40.9% by the end of the 6-day recovery period and explained that shoot elongation of these genotypes represents transient submergence intolerance. Studying the emergence rate of plants, Su et al. (2022) concluded that submergence-tolerant rice will rapidly elongate in a submerged environment and by that it becomes exposed to oxygen outside the water where the supply of oxygen and the production of carbohydrates can be maintained to improve plant survival. That is why escape strategy in rice is shown by rapid underwater shoot elongation especially in deep water lowland rice where vigorous stem extension prevents total submergence with increased depth of flood water and keep the upper leaves above water surface for normal gas exchange, and thereby survive submergence (Wu et al., 2018; Kuanar et al., 2019).
On the other hand, the shoot length measured after 11 days of complete submergence was reduced by 13.7%. Tolerant check FR13A as well as genotypes that showed some survival degree such as NBGA, Sommboy and Pipanfary Red2 reduced their shoot elongation as tolerant genotypes, were found to conserve energy during flooding for maintenance and survival processes as strategy in response to short term inundation (Pramanik et al., 2016). Likewise, the highest (10.98cm and 10.54cm) root lengths measured after 11 days of complete submergence was achieved by the genotypes Pipanfary Red2 and Masury2, respectively, which were higher (7.46cm) than that of FR13A and the lowest (3.45 cm) root length was obtained by Sommboy lower (7.46 cm) than that obtained for IR42 (Table 1). From the results, it is clear that root elongation followed an opposite pattern to that of shoots, whereby the accessions that showed tolerance registered higher root lengths as compared to intolerant ones. It is interesting to note that all genotypes that showed zero percent survival under submergence, such as Tagmezido, Pipanfary and Pipanfary Red1 registered a 2 to 3 folds reduction in root length under submerged conditions. This significant reduction in root length of most genotypes may be due to complete submergence, which greatly reduces the amount of oxygen that reaches roots via arenchyma thereby limiting root development.
On the other hand the tolerant check FR13A and genotypes with some degree of survival (e.g., Pipanfary Red2, Banban, Masury1 and NBGA) either had similar root lengths under both controlled and submerged conditions or increased their root length values when submerged. Increased root length with plant survival after submergence might explain the ability of the tolerant genotypes to maintain carbohydrate supply and aeration to support growth and functioning of the roots. Under submerged conditions root system elongation support the upright rooting of seedlings and the higher root volume forms a larger root surface area, thereby increasing the amount of absorbed oxygen and nutrients underwater (Su et al., 2022).
Variation in carbohydrate concentration in seedlings of 20 rice genotypes under control and complete submerged conditions
Statistical analysis, under both control and submerged conditions, showed highly significant (P≥0.01) differences between the genotypes for soluble sugar and starch concentration in roots, stems and leaves at all sampling dates (Tables 2 and 3). Mean soluble sugar concentrations in roots, and leaves measured after 32 days of seedling growth under control conditions were lower by 16.6, and 5.8%, respectively, than those measured 21 days of seedling growth, while that of stems was more or less the same. This reduction observed among the genotypes indicates that soluble sugar decreases with increasing time of seedling growth (Table 2). On the other hand, mean starch in stems and in leaves measured after 32 days seedling growth was reduced by 77 and 22%, respectively, than that measured 21 days seedling growth, with exception of some genotypes, whereas mean starch concentration in roots was increased by 3.6%. Highest increase was exhibited by some genotypes such as Pipanfary Red2, FR13A (Table 2).
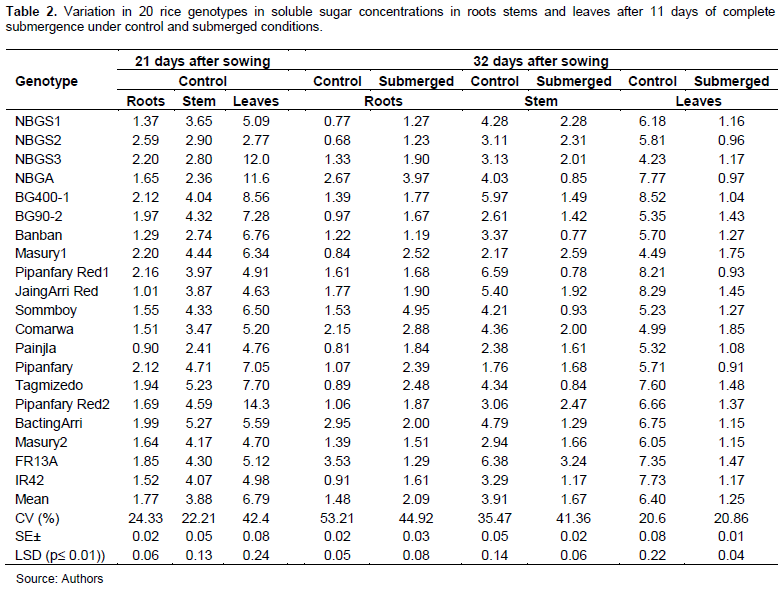
Under complete submergence, mean soluble sugar concentration in roots was increased by 41%. Regarding individual genotypes, BactingArri showed 32.2% decrease in mean soluble sugar concentration, while Sommboy registered the highest (69%) increase in mean soluble sugar among genotypes followed by Masury1 (66.3%) and the tolerant check FR13A (63.5%). Mean soluble sugar in stems was reduced under complete submergence by 57.3%.
However, Masury1 achieved an increase by (16.1%). The greatest (88.2%) reduction in stem soluble sugar concentration was registered for Pipanfary Red1 and the least (4.5%) was recorded for Pipanfary. Mean soluble sugar in leaves was reduced by 80.5%, highest (88.7%) reduction in mean soluble sugar concentration was registered for Pipanfary Red1 and least (61.0%) reduction was observed for BactingArri (Table 2). The results suggest the effects of flooding on conversion of soluble sugars into starch. These findings are in agreement with those of Samanta et al. (2020) who reported on the notable reduction in the soluble sugar content in plants under submerged condition in comparison to those under control conditions and that tolerant genotypes showed less reduction in their sugar content than sensitive ones.
Mean starch concentration in roots was reduced by 13.8% under complete submerged condition than under controlled condition. It is clear from these results that soluble sugar concentrations were higher than starch concentrations in roots, stems and leaves under controlled and submerged conditions as well as the higher increase in soluble sugar might indicate higher rates of solubilization of structural carbohydrates supply during submergence (Samanta et al., 2020).
Mean starch concentration in stems was reduced by 77.7%. However, Pipanfary, registered an increase of 5%, the highest (81.4%) reduction was recorded for IR42 and least (37.7) reduction was by Painjla. Mean starch concentration in leaves was increased by 115% for all genotypes with Pipanfary Red2 registering the highest (236%) increase and IR42 the least (57.9%) increase (Table 3). High rates of soluble sugars are necessary for elongation during submergence period and insoluble sugar is used for maintenance of structural build-up during recovery period, thus initially higher content of soluble and insoluble sugars can be regarded as an advantage for submergence tolerance (Sayani et al., 2017).
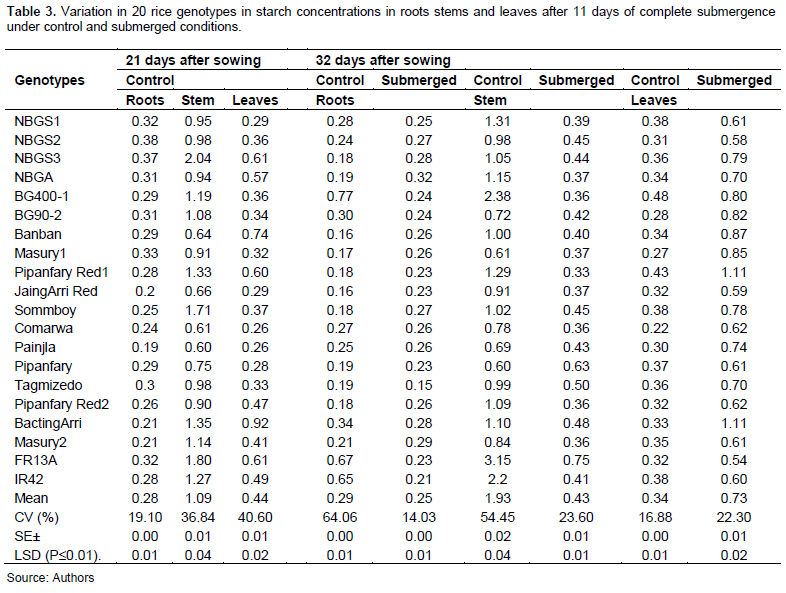
Under submerged condition, the increase in mean soluble sugar concentration in roots and reduction in mean starch concentration, suggest that the genotypes used some stored starch for the growth of roots under complete submerged condition, in which tolerant genotypes, revealed lower soluble sugar and starch concentrations.
The reduction in mean soluble sugar concentration and starch concentration in the stems indicate that tolerant genotypes obtained higher soluble sugar and starch concentrations than the intolerant ones as the reduction of photosynthetic function caused by changes in plant photosynthetic pigments under adverse conditions was found to affect the production of carbohydrates and plants survival (Su et al., 2022).
In leaves, the high reduction in mean soluble sugar concentration and high increase in mean starch concentrations for all genotypes, might indicate that the genotypes conserved their starch to be used by the plant for recovery after de-submergence. The higher reserved carbohydrate in the tolerant varieties might assist in a faster restart of their normal growth after desubmergence (Pramanik et al., 2016).
Data in Table 4 show highly significant (P≥0.01) differences between the genotypes for chlorophyll a, b and total chlorophyll (a+b) content under control and submerged conditions. These results are in contrast with those of Panda and Sarkar (2014) who reported non-significant effects of chlorophyll concentrations under control condition.
Under control condition, chlorophyll concentrations in plant leaves were increased with time among the genotypes. Chlorophyll a, chlorophyll b and total chlorophyll (a+b) concentrations measured after 32 days of seedling growth were higher by 15.8, 29.7 and 20.1%, respectively, than those measured after 21days of seedling growth, with exception of some genotypes (Table 4). The highest chlorophyll a and total chlorophyll (a+b) concentrations were observed for genotype BG90-2, whereas the highest chlorophyll b concentration was registered by genotype NBGS2 and the lowest chlorophyll a, chlorophyll b and total chlorophyll (a+b) concentrations, were obtained for the tolerant cultivar FR13A.
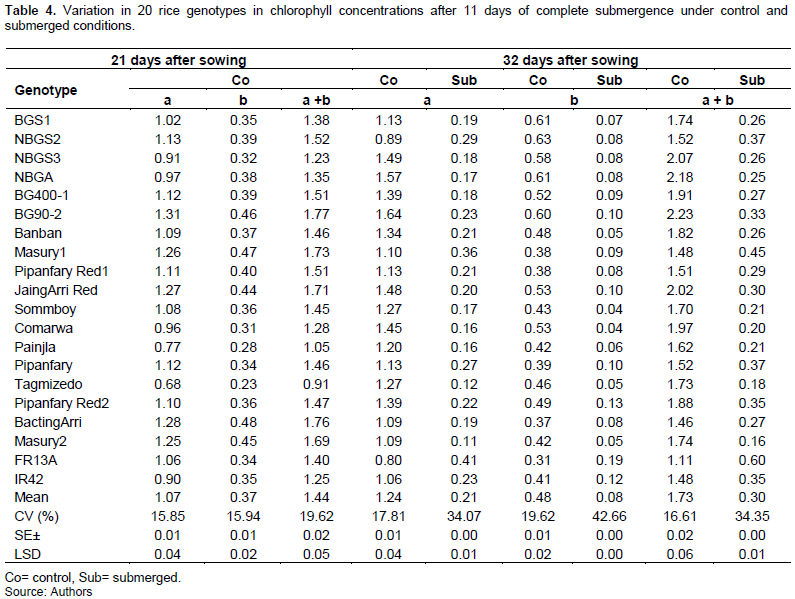
Submergence reduced chlorophyll a, chlorophyll b and total chlorophyll (a+b) concentrations among the genotypes by 83, 83.3 and 82.7% respectively. This reduction had most effect on intolerant genotypes as compared to the tolerant ones. Similar findings were reported by Nurrahma et al. (2021b) who found that submergence lowered concentrations of chlorophyll by72.4% versus control in a sensitive genotype as compared to only 14.4% reduction in a tolerant genotype.
Tolerant genotypes are able to maintain vital processes, principally photosynthesis, minimizing the detrimental effects of submergence stress (Nurrahma et al., 2021b).
In this study, the highest chlorophyll a, chlorophyll b and total chlorophyll (a+b) concentrations, were achieved by the tolerant check FR13A and the lowest chlorophyll a was shown by Masury2.
While, the lowest chlorophyll b was obtained for accessions Sommboy and Comarwa, and lowest total chlorophyll concentration (a+b), was registered by accession Masury2 (Table 4).
Leaf photosynthesis is one of the earliest plant responses to submergence, which resulted in significant reduction of chlorophyll a, chlorophyll b and total chlorophyll (a+b) after 11 days submergence in both sensitive and tolerant genotypes, in which the intolerant genotypes were the most affected ones. On the other hand, the tolerant genotype FR13A and the Sudanese genotypes (Masury 1, Pipanfary Red1) which showed survival after submergence for 11 days, maintained significantly higher values of chlorophyll a, chlorophyll b and total chlorophyll (a+b). This shows that genotypes with higher chlorophyll concentrations during and after submergence have the highest survival. Su et al. (2022) reported that chlorophyll is the medium for absorption, transformation and light energy transmission in plants, therefore under submergence plants with higher chlorophyll content have better chances of carbohydrates production and plants survival and concluded that the level of chlorophyll content can be used to evaluate the potential of dry matter production and plant survival.
CONCLUSION
From the results of this study, it could be concluded that submergence tolerant genotypes such as Pipanfary Red2 and Sommboy had higher soluble sugar and starch concentrations and chlorophyll content, required for plant survival under stress and recovery afterwards. These genotypes could be developed to be grown under rain-fed lowland flooded areas of Sudan and South Sudan.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
Authors thankfully acknowledge the support of IRRI technicians for the technical assistance. Thanks are also extended to the Agricultural Research Corporation, Kosti, Sudan and Ministry of Agriculture, Bahr Elgabal State, South Sudan for providing the germplasm.
REFERENCES
|
Chen FX, Tang YP, Xie YJ (2018). Research Advances of Rice Mechanical Direct-Seeding Technology in China. China Rice 24(4):9-15. |
|
|
Debarbata P, Jijnasa B (2021). Flooding Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Science 28(1):43-67. |
|
|
Fales FW (1951). "The assimilation and degradation of carbohydrates by yeast cells". Journal of Biological Chemistry 193:113-124. |
|
|
Gasim S, Abuanja I, Dafalla M, Abdalla A (2018). Screening for sodium-salt tolerance at the seedling stage of rice (Oryza sativa L.) genotypes from Sudan and South Sudan. Journal of Agricultural Science and Technology B 8(2018) 341-351. |
|
|
Gomez KA, Gomez AA (1984). Statistical Procedures for Agricultural Research. 2nd edition. John Willey and sons, New York. |
|
|
Kuanar SR, Molla KA, Chattopadhyay K (2019). Introgression of Sub1 (SUB1) QTL in mega rice cultivars increases ethylene production to the detriment of grain-filling under stagnant flooding. Science Report (9):18567. |
|
|
Kunst A, Draeger B, Ziegenhorn J, Bergemeyer HU (1988). "Colorimetric methods with glucose oxidase and peroxidase. Methods of enzymatic analysis". Metabolites I. Carbohydrates 6:178-185. Verlag-Chemie, Weinheim, Germany. |
|
|
Lichtenthaler HK, Wellburn AR (1983). "Determinations of total carotenoids and chlorophylls a and b in leaf extracts in different solvents". Biochemistry and Social Transaction 11:591-592. |
|
|
Mackinney G (1941). Absorption of light by chlorophyll solutions. Journal of Biological Chemistry 140(2):315-322. |
|
|
Nurrahma AHI, Yabuta S, Junaedi A, Sakagami JI (2021). Analysis of non-structural carbohydrate in relation with shoot elongation of rice under complete submergence. Sustainability 13:670. |
|
|
Nurrahma AHI, Yabuta S, Junaedi A, Sakagami JI (2021). Characterizing the photosynthetic ability of the submergence-tolerant rice variety of Inpari30 via maximum quantum yield performance during transient flooding stress and recovery. Australian Journal of Crop Science 15(1):107-113. |
|
|
Pramanik MHR, Shelley IJ, Adhikary D, Islam MO (2016). Carbohydrate reserve and aerenchyma formation enhance submergence tolerance in rice. Progressive Agriculture 27(3):256-264. |
|
|
Rachmawati D, Maryani MM, Kusumadewi S, Rahayu F (2019). Survival and root structure changes of rice seedlings in different cultivars under submergence condition. Biodiversitas 20(10):3011-3017. |
|
|
R-STAR (2013)."Statistical Tools for Agricultural Research".Version 1.1. International Rice Research Institute (IRRI), Los Baños, Philippines. |
|
|
Samanta P, Ganie SA, Chakraborty A, Dey N (2020). Study on regulation of carbohydrate usage in a heterogeneous rice population under submergence. Journal of Plant Biochemistry and Biotechnology 30(1):138-146. |
|
|
Sayani G, Kumar KR, Anupam P, Narottam D (2017). Study of selected biochemical parameters related to submergence tolerance in rice (Oryza sativa L.) with special reference to land races and wild species. Research Journal of Chemistry and Environment 21(11):29-38. |
|
|
Osman KA, Mustafa AM, Ali F, Zheng Y, Qui F (2012). Genetic variability for yield and related attributes of upland rice genotypes in semi-arid zone (Sudan). African Journal of Agricultural Research 7(33):4613-4619. |
|
|
Setter T L, Laureles CV (1996)."The beneficial effect of reduced elongation growth on submergence tolerance of rice". Journal of Experimental Botany 47(10):1551-1559. |
|
|
Su X, Wu H, Xiang J, Zhan JJ, Wang JQ, Li XM, Wei YH, Dai HF, Chen HZ (2022). Evaluation of submergence tolerance of different rice genotypes at seedling emergence stage under water direct seeding. Open Access Library Journal 9(5):1-15. |
|
|
Sun K, Li DX, Yang J (2019). Genome-wide association analysis for rice submergence seedling rate. Scientia Agricultura Sinica 52(3):6-19. |
|
|
Wu H, Xiang J, Chen HZ, Zhang YP, Zhang YK, Zhu DF (2018). Effects of exogenous growth regulators on plant elongation and carbohydrate consumption of rice seedlings under submergence. Chinese Journal of Applied Ecology 29(1):149-157. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


