Information on the combining ability and heterotic pattern of elite inbred lines is essential to maximize their use in hybrid maize development. This study was conducted to determine combining ability and heterotic pattern of locally developed maize inbred lines for grain yield and related traits. Seventeen inbred lines (10 female inbred lines and 7 tester inbred lines) were used to generate 70 single cross hybrids using line by tester crossing scheme. The resulting 70 cross progenies plus two standard checks arranged in 8×9 alpha lattice design replicated twice were planted at three mid-altitude sub-humid testing sites in Ethiopia (Bako, Hawassa and Pawe) in 2011 main cropping season. The combined analysis of variance for yield and other related traits showed highly significant differences among genotypes, crosses, female inbred lines (General combining ability, GCA), tester inbred lines (GCA), line x tester (Specific combining ability, SCA); and the interactions of these source of variation with the environment for all traits studied except for ear aspect (EA) and grain yield (GY) in female inbred lines (GCA), EA in inbred line testers (GCA) and for days to anthesis (AD) in line x tester (SCA) x environment. The significance of both GCA (lines and testers) and SCA of LxT for AD, days to silking (DS), plant height (PH), ear height (EH), EA and GY showed that both additive and non-additive gene actions are important in controlling these traits. Furthermore, the proportion of GCA sum of squares were greater than the SCA sum of squares for AD, DS, PH, EH, and EA indicating the predominance of additive gene actions in controlling these traits. For GY, the ratio of GCA to SCA sum of squares was near to unity indicating both additive and non-additive gene actions were equally important. This study identified inbred lines that can make good cross combination for more than one trait. L1 was found to be good combiner for lower values of AD, DS, PH and EH indicating that this line could be used in improving maize for earliness and short stature. L4 was ideal parent for reducing AD and DS. L3 was found to be good combiner for GY and other related traits. In addition, lines were grouped into heterotic group A, B or AB based on SCA. Based on its per se performance and combining ability, L3 was proposed to be used as a tester in heterotic group B. This study also validated T5 remain to be used as a tester in heterotic group A. Based on the SCA of crosses, heterosis and per se performance of the parents, five best cross combinations were identified for possible release or for use as parents of three way hybrids. Further verification of the stability of the selected hybrids and the new proposed tester across more locations needs to be done.
Maize (Zea mays L.), together with wheat (Triticum aesivum L.) and rice (Oryza sativa L.) is one of the three most important cereal crops that feed two - third of the world population (Ji et al., 2013). In Sub - Saharan Africa (SSA), maize is a major staple cereal food crop with the significance comparable to rice in South East Asia and wheat in the Middle East. Maize being the highest yielding cereal crop in the world is of significant importance for countries like Ethiopia, where future food supply would be a great challenge for the rapidly increasing population. Ethiopia currently produces more maize than any other crop (McCann, 2005; Abate et al., 2015; FAOSTAT, 2015). In 2014, maize was grown on about 2.1 million hectares, resulting in total annual production of 7.2 million tons, with an average of 3.42 tons per hectare (FAOSTAT, 2015). It is a primary crop in majority of farming systems and staple food of the rural population in much of the mid-altitude sub-humid agro-ecology of the country. Maize stands first in total production and productivity and, second in area coverage (20.27%) next to tef [Eragrostis tef (Zucc) Trotter] (30.66%) of all cereal crops cultivated in Ethiopia (FAOSTAT, 2015).
The hybrid development in Ethiopia has been highly effective in increasing maize yields since the commercialization of the hybrids in the country. The national average grain yield increased from about 1.6 ton ha−1 in 1990 (Abate et al., 2015; Worku et al., 2002; Mosisa et al., 2011) to 3.42 tons ha−1 in 2014 (FAOSTAT, 2015). Increased yields are in part due to improved agronomic practices and increased inputs, but increased yields could not have been realized without genetic improvements (Abate et al., 2015). However, the current national productivity average per hectare (3.42 tons ha-1) (FAOSTAT, 2015), is considerably low as compared to developed countries. The national average grain yield per hectare of some of the countries, for example, is 10.68, 10.73, 9.36, 5.18, and 6.0 tons ha-1 respectively for USA, Canada, Germany, Brazil and China (FAOSTAT, 2015). This emphasizes that further genetic improvement efforts have to be made to increase maize productivity in the country.
Information on the combining ability of parental inbred lines is very important for determining breeding strategies, classifying the parental inbred lines, defining heterotic groups, and predicting future hybrid performance (Xingming et al., 2004; Legesse et al., 2009). Classifying inbred lines into heterotic groups is critical to determine the potential usefulness of the inbred lines for the development of high yielding hybrids and synthetic varieties. Therefore, knowledge on the heterotic groups of inbred lines is important before they can be deployed in variety development.
Understanding the genetic basis for hybrid performance and identifying parental inbred lines that form superior hybrids is crucial in designing appropriate breeding strategies. Further advancement in the yield of maize requires certain information regarding the nature of combining ability of the parents available for use in the breeding program as well as the nature of gene action involved in expression of both quantitative and qualitative traits of economic importance.
General combining ability (GCA) and specific combining ability (SCA) effects are important indicators of the potential value of inbred lines in hybrid combinations and in grouping materials into heterotic groups. The use of heterotic groups, when aided with good testers in a breeding program can result in the production of high yielding hybrids. Testers of hybrid value or heterosis between parental inbred lines can increase the efficiency of hybrid breeding programs (Legesse et al., 2009). Estimates of GCA and SCA will help to devise breeding and selection strategies. The use of GCA and SCA estimates has proven effective in maize breeding program to identify superior hybrids, find the best parent for hybrid formation, and choose material for new heterotic groups (Hallauer and Miranda, 1958). Although some genetic studies have been conducted in maize on commercial inbred lines (Dagne, 2002; Dagne et al., 2010; Hadji, 2004), it is a continuous and an endless process that the information on combining ability and heterotic pattern of the newly developed inbred lines needs to be determined. Therefore, the objectives of this study were (a) to determine the combining ability of new mid-altitude sub-humid tropical maize inbred lines (b) to categorize the inbred lines into different heterotic groups based on SCA and heterosis for grain yield and (c) to identify single crosses which have better agronomic traits than the commercial checks.
Description of experimental sites
The study was conducted at three locations in the mid-altitude sub-humid agro ecology of Ethiopia; namely, Bako, Hawassa and Pawe Agricultural Research Centers in the main cropping season of 2011. Bako Agricultural Research Center (BARC) is located at 9°6’N latitude and 37°09'E longitude, 255 km west of Addis Ababa at an altitude of 1650 m above sea level (masl). The soil type at BARC is characterized by reddish brown clay (nitosol) with pH (H2O) of 6.0 (Yadessa et al., 2005). The site receives an average annual rainfall of 1245 mm and the mean minimum and maximum air temperatures are 13.3 and 27.9°C, respectively. Pawe Agricultural Research Center (PARC) is located at 11°12’N latitude and 36°20’E longitude at an elevation of 1150 masl. The annual precipitation of the testing site ranges from 1338.7 to 2005.7 mm with a mean of 1603.5 mm. The mean annual maximum and minimum temperature of the area are 38.54 and 16.36°C, respectively. Hawassa Agricultural Research Center (HARC) is situated at 7°08’N latitude and 38°48'E, 275 km south of Addis Ababa at an altitude of 1700 masl. The soil at HARC is volcanic in nature. The site receives average annual rainfall of 1110 mm and the mean minimum and maximum air temperatures are 13 and 27.4°C, respectively.
Experimental materials
Seventy single cross hybrids were generated from crosses of 10 white grained locally developed tropical mid-altitude sub-humid maize inbred lines used as females (L) and seven white grained inbred lines (five from CIMMYT and two from the National Program) used as males/testers (T), using line by tester mating scheme at Bako Research center in 2010 (Table 1). Among the male parents, T1 (from heterotic group B) and T2 (from heterotic group A) were locally developed inbred lines by the national maize breeding program having good combining ability. The remaining five male parents (T3, T4, T5, T6 and T7) were also CIMMYT inbred lines having defined heterotic groups.
Experimental design and trial management
In 2011, the 70 F1 single cross progenies along with two standard hybrid checks (BH540, single cross hybrid and BH543, three-way cross hybrid) were planted at Bako, Hawassa and Pawe. The experiment was planted in an alpha-lattice (8×9) design (Yadessa et al., 2005), with two replicates. Each entry was planted in a two-row plot of 5.1 m long and 0.75 m apart with a distance of 0.3 m between plants within a row.
The trials were hand planted with two seeds per hill, which later were thinned to one seed per hill to get a total plant population of 44,444 per hectare. Fertilizers were applied as Di-ammonium Phosphate (DAP) and urea at the rates, kg ha-1 of 100 N and 100 P2O5 at Bako, 110 N and 46 P2O5 at Hawassa and 75 N and 70 P2O5 at Pawe. All recommended rates of P2O5 were applied at the time of planting while N was applied in split, half at planting and the remaining half at 35 to 40 days after planting. Pre-emergence herbicide, Primagram-gold 660 SC was applied at the rate of 3lha-1 at planting to control weeds.
Data collection and analysis
Data were collected on grain yield (GY) (adjusted to 12.5 grain moisture and expressed as tons ha-1), plant (PH) and ear height (EH) (cm), days to 50% anthesis (DA) and silking (DS), and ear aspect (EA) (1-5 scale) on plot basis at each of the three locations.
The data obtained for different traits from field measurements were organized and analyzed using SAS statistical package (SAS, 2009). The significance of each trait in each location, ANOVA for combining ability for individual locations and across locations was determined.
Analysis of variance per location was conducted with the PROC MIXED procedure in (SAS, 2009) where entries were treated as fixed factor; while location, replication and incomplete blocks within replication were considered as random factors. Entry means adjusted for block effects generated from individual location analysis according to lattice design (Cochran and Cox, 1957) were used to perform the combined analysis using the PROC GLM procedure in SAS (2009). Then, the data were subjected the F test and the significance at 5 and 1% probability levels was evaluated.
For traits that showed significant differences among crosses, further analysis was performed following the procedures of Singh and Chaudhary (1979) and Dabholkar (1999) to partition the mean square due to crosses into lines, testers and line by tester(LxT) effects using SAS computer program (SAS, 2009) for individual location and combined across locations. The mean squares for genotypes, crosses, checks and location were tested against the mean squares for their corresponding interaction with location as error term while their interaction with location were tested against their corresponding pooled error. Since means (over replications) were used for combined analysis of variance, estimates of pooled error mean squares were calculated following the procedure of Dabholkar (1999).
Estimation of combining ability effects
Genotypic means of individual locations were used for the determination of GCA and SCA. The two standard checks were excluded while analyzing combining abilities. The GCA effects of lines (L) and testers (T), the SCA effect of LxT, and their interactions with the environment were determined following the method stated by Kempthorne (1957), assuming the following model.
Y = µ + gi + gj + Sij +ek + (ge)ik +(ge)jk +(se)ijk +d ,
Where Yijk= the performance of the hybrid, made with ith female and jth male, in the kth location, μ = the overall mean, gi = the effect of the ith female, gj = the effect of the jth male, sij = the interaction of the ith female with the jth male, ek = the effect of the kth environment, (ge)ik = the interaction of the gi and ek, (ge)jk = the interaction of the gj and ek, (se)ijk = the interaction of sij and ek.
Significance of GCA and SCA effects were performed computing the standard error for lines, testers and crosses and then tested against t-test by taking the degree of freedom of pooled error mean square (Singh and Chaudhary, 1979; Dabholkar, 1999; Sharma, 2003). The proportional contributions of lines (GCAL), testers (GCAT), and their interaction (SCALxT) to the sum square of crosses were calculated as the ratio between sum of squares of each component and the cross sum of squares as given by Sharma (2003) and Dabholkar (1999).
Orientation of lines into heterotic group A and B depended on the direction of the specific combining ability such that lines exhibiting positive SCA with tester A were allocated to the opposite heterotic group B, and vice versa, whereas lines displaying positive SCA to both were designated as AB group.
Analysis of variance of Individual location
The mean squares of analysis of variance for different characters at Bako, Hawassa and Pawe are presented in Table 2. At Bako, significant differences were observed among the genotypes in grain yield and other traits (viz. DA, DS, PH, EH, EA). Likewise, the genotypes showed significant differences in their reaction to different foliar diseases such as grey leaf spot (GLS) and Turcicum leaf blight (TLB) (data not shown). Similarly, significant differences in DA, DS, PH, EH, EA, and grain yield among the genotypes were observed at Hawassa. The genotypes were also significantly different in their reaction to common leaf rust (CLR) at this site (data not shown). At Pawe, significant differences were observed among the genotypes in DA, DS, PH, EH, EA, and grain yield. There were no foliar diseases observed in the experimental field during the cropping season at Pawe.
While there were significant differences among the genotypes in six of the traits under investigation (viz., DA, DS, PH, EH, EA and GY) at each of the three locations, there were either no significant differences among the genotypes or there were significant differences among the genotypes at most at two environments in the remaining traits under study (data not shown). The combined analysis of variance across environments and combining ability analysis were, therefore, performed only for the traits that were significantly different among the genotypes at each of the three environments.
Combined analysis of variance
The combined analysis of variance for grain yield and other related traits showed highly significant differences (P<0.01) among genotypes, crosses, lines (L), testers (T), L x T; and the interactions of these source of variation with the location for all traits studied except for ear aspect and grain yield in lines, ear aspect in testers and for days to anthesis in line x tester x environment (Table 3). Similarly, significant differences were observed in checks x location and crosses vs check x location for grain yield. The result of the combined analysis of variance across the three environments revealed that there were significant differences among the genotypes for days to anthesis days to silking, plant height, ear height, ear aspect and grain yield. Various scientists (Asefa, 2004; Zeleke and Nepir, 2007; Kanyamasoro et al., 2012) have also reported similar findings in other groups of inbred lines they studied.
The significance of the mean squares of GCA (of lines and testers) and SCA of L x T for the traits under investigation indicated the importance of both additive and non-additive gene effects for these traits. This is consistent with the findings of other authors (Meseka et al., 2006; Alam et al., 2008; Wegary et al., 2011). The combined ANOVA of combining ability showed that the GCA sum of squares of lines were greater than the GCA sum of squares of testers for all traits except ear aspect. Similarly, the GCA/SCA ratio being greater than unity for days to AD, DS, PH and EH, indicated that additive gene effects were more important for these traits (Table 3) in this group of inbred lines. For grain yield, the GCA to SCA sum of square ratio being slightly near to unity indicated that both additive and non-additive genetic effects were equally important in the inheritance of this trait. Significant mean squares (P<0.01) of environments for DA, DS, PH, EH, EA and GY revealed that the responses of the genotypes across the three environments were different for these traits. The significance of mean squares due to genotypes (P<0.01) for the traits studied indicated the existence of genetic variability among the genotypes.
The significance of the interaction of GCA of parents (lines and testers) with the environment and SCA of the crosses with the environment revealed that the GCA effects of the parents and SCA of the crosses over the test environments were different. Mean values of six traits (viz. DA, DS, PH, EH, EA and GY) averaged across three environments for top 10 entries and GY advantage over the best check are presented in Table 4.
DA ranged from 69.83 to 79.33 days with a mean of 74.94 days. Similarly, DS ranged from 73.00 to 81.67 with a mean of 77.85. The lowest mean values for both DA and DS were observed in cross L4xT2, while the highest mean values for both the traits were observed in the cross L6xT6. As a result, L4xT2 could be preferred for early maturity.
For PH, the crosses ranged from 190.5 to 277.17 cm with a mean of 243.5 cm. Similarly, the crosses ranged from 89.33 to 160.67 cm for EH while the mean was124.90 cm. The highest mean values for PH and EH were observed in the cross L3xT5; while the lowest mean values for the same traits (not included in Table 4) were noted for the cross L10xT6 making this cross more preferable for tolerance to lodging. The entries ranged from 1.33 to 2.92 for their EA with a mean of 2.06. The highest and lowest mean values for ear aspect were observed in crosses L4xT1 and L9xT5 (not included in Table 4), respectively. Because the smaller values of EA are desirable, the cross L9xT5 was preferred for this trait. In GY, the entries ranged from 5.74 to 10.77 ton ha-1 with a mean of 8.68 ton ha-1. The two checks used in this experiment, BH543 and BH540, had mean grain yield values of 7.70 and 8.99 ton ha-1, respectively. The Mean grain yield of ten entries exceeded the mean grain yield of the best check (BH540 in this experiment) by over 10% is presented in Table 4. Further testing of these hybrids is needed to see their yield stability across more environments.
General combining ability of female inbred lines
The GCA effects of the 10 female inbred lines for GY and related traits are presented in Table 5. Among the 10 inbred lines, only L3 was with significantly positive mean GCA effects for GY. Therefore, this inbred line is good combiner for improving GY. In contrast, L5 had significantly negative GCA effects for GY, indicating, that this inbred line was not good combiner within this group of inbred lines and may not be exploited for GY improvement. In line with the current study, several authors reported either positive or negative significant GCA effects of inbred lines for GY in other group of inbred lines (Legesse et al., 2009; Dagne, 2002; Zeleke and Nepir, 2007; Ahmad and Saleem, 2003; Worku et al., 2009; Mosa, 2010). In contrary to this, Asefa et al. (2008) found non-significant GCA effects for GY.
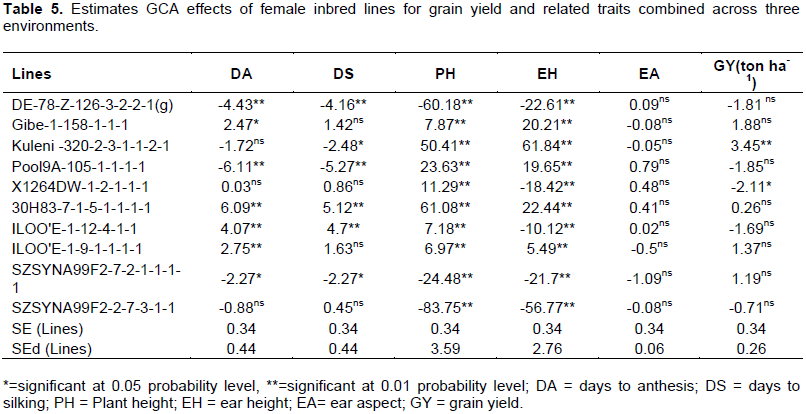
The results displayed that L2, L6, L7 and L8 had significantly positive GCA effects for DA, whereas L1, L4 and L9 had significantly negative GCA effects for this trait. The remaining three of the 10 female inbred lines had no significant GCA effects for DA. For DS, L6 and L7 had significant positive GCA effects, whereas L1, L3, L4 and L9 had significantly negative GCA effects, and the remaining four of the 10 female inbred lines had positive but non - significant GCA effects. It has been noticeable that the frequency and distribution of rain, and length of the rainy season particularly in the mid altitude sub-humid agro-ecology of Ethiopia where it used to be for more than five consecutive months is becoming less than four months (data not shown) as the past few years were good examples. This is believed to be attributed to the global climate change. This phenomenon, therefore, emphasizes the importance of early to mid-maturing varieties than late maturing varieties in the mid altitude sub-humid agro-ecology of Ethiopia. As a result, due to the importance of relatively early maturing and lower values of DA and DS, inbred lines which had significantly negative GCA effects were considered as good combiners for the improvement of these traits. These results are in general agreement with the findings of many authors (Dagne, 2002; Ahmad and Saleem, 2003; Mosa, 2010; Bello and Olaoye, 2009).
Seven of the 10 female inbred lines (viz., L2, L3, L4, L5, L6, L7 and L8) had significantly positive GCA effects for PH while the remaining three lines (L1, L9 and L10) had significantly negative GCA effects for the same trait (Table 5). While five of the 10 inbred lines (viz. L2, L3, L4, L6 and L8) had significantly positive GCA effects for EH, the remaining five inbred lines (viz. L1, L5, L7, L9 and L10) had significantly negative GCA effects for this trait. Again, due to the reason that the lower plant height and ear height, the more the plant is expected to be resistant / tolerant to lodging, inbred lines which had significantly negative GCA for these traits were considered to be desirable for the improvement of PH and EH. Similar findings have been reported by several authors (Hadji, 2004; Mosa, 2010; Rahman et al., 2010).
General combining ability of tester inbred lines
The GCA effects of the seven tester inbred lines for GY and other five agronomic traits are presented in Table 6. Two of the seven male inbred lines, namely; T5 and T7 had significantly positive GCA effects for GY, whereas T3 had significantly negative GCA effects for the same trait. Consequently, T5 and T7 were considered good combiners for the improvement of GY within this group of inbred lines included in this study. The GCA effects of the remaining four tester inbred lines were not significant for grain yield, all with the same negative sign with different magnitudes.
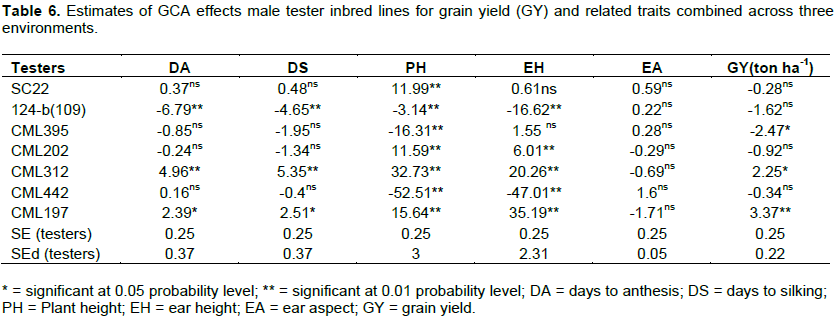
The GCA effects of two of the seven male tester inbred lines (viz. T5 and T7) were significantly positive for both DA and DS while that of T2 was significantly negative for these traits. Therefore, T2 is preferable for the improvement of these traits. The GCA effects of the remaining four of the seven tester inbred lines for both AD and DS were not significant either positively or negatively. For PH, four tester inbred lines namely; T1, T4, T5 and T7 had significantly positive GCA effects whereas T2, T3 and T6 had significantly negative GCA effects. Three of the seven tester inbred lines (viz. T4, T5, and T7) had significantly positive GCA effects for EH, while two tester inbred lines (viz. T2 and T6) had significantly negative GCA effects for the trait. The GCA effect of none of the tester inbred lines was significant for EA (Table 6).
Specific combining ability of crosses
Maximum and minimum estimates of specific combining ability effects (SCA) of the 70 crosses across three locations for GY and other five related traits are presented in Figure 1. Two of the 10 female inbred lines (L1 and L8) had significantly positive SCA effects when crossed to T1 for GY implying that there was significant positive interaction of genes between the two parents for this trait. Such gene interactions lead to the expression of heterosis, which can be exploited in the development of hybrid varieties. Similarly, the crosses of L4xT3 and L5xT2 had significantly positive SCA effects for GY indicating the importance of non-additive gene action in these cross combinations (data not shown).
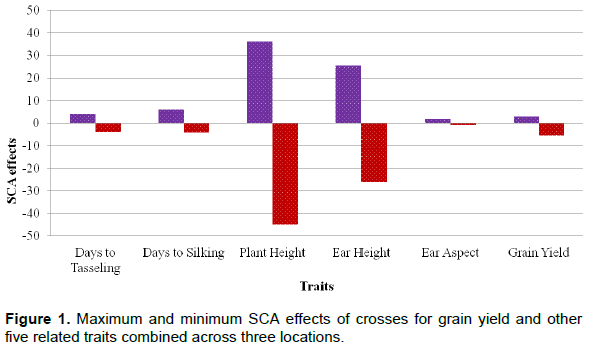
For AD, the crosses L1xT7, L4xT4, L5xT5, L5xT6, L6x T6, L7xT2, L7xT3, L9xT1 and L10x T3 had significantly positive SCA effects, and, hence, were not good cross combinations for this trait, as early maturing varieties are desired. Conversely, the crosses L1 x T6, L2 x T5, L3 x T2, L4 x T2, L5x T2, L5 x T3, L6 x T1 and L6 xT4 had significantly negative SCA effects for AD indicating that these crosses were good cross combinations for early maturity. The crosses L1xT7, L4xT4, L4xT7, L5x T5, L5xT6, L6xT6, L7xT3, L8xT5, L10xT2 had significant positive SCA effects for DS indicating that these cross combinations were not desirable crosses for early maturity. The crosses L1xT6, L2xT5, L4xT2, L4xT6, L5x T2, L5xT3, L7x T7, L8xT7, L9x T7 and L10 x T6 had significant negative SCA effects for DS. These crosses were considered good cross combinations for early maturity (data not shown).
Fifty-eight of the 70 crosses had significant SCA effects for PH, and out of that 30 were with positively significant and 28 were negatively significant SCA effects (data not shown). The crosses L1xT7, L4xT4, L8xT3, L3xT2 and L9x T1 were with the highest significant negative SCA effects for PH and were considered the best cross combination for being likely to be tolerant to lodging. Likewise, 31 of the 70 crosses had significant negative SCA effects for EH indicating that these cross combinations may be exploited for tolerance to lodging. None of the crosses had significant SCA effects for EA. Crosses L1xT7 and L8xT1had the highest positive and negative SCA effects for EH, respectively. The crosses that had negative values of SCA for EA were considered desirable as the higher values of EA score indicates bad ear characteristics.
Heterotic orientation
Grain yields of 70 crosses averaged over three environments and estimates of SCA effects of these crosses are presented in Table 7. Among the male inbred lines, T1 and T2 are locally developed inbred lines by the national maize breeding program having good combining ability; and have been used as testers in maize breeding for mid-altitude sub-humid agro-ecology of Ethiopia. Either one or both of these inbred lines are present in pedigrees of two of the commercial hybrids used as standard checks in this study (T1 in pedigree of BH540, and both T1 and T2 in pedigree of BH543). SC22 was developed from SC5522 (heterotic group B) while 124-b (109) was developed from Kitale Synthetic II (heterotic group A) (Tolesa et al., 1993). The testers T3, T4, T5, T6 and T7 are CIMMYT inbred lines having known heterotic groups with T3 and T4 being in heterotic group ‘B’ whereas, T5, T6 and T7 are in heterotic group ‘A’ (Tolesa et al., 1993). Four of these (viz. T3, T4, T5 and T6) have been used as testers in CIMMYT and other national and international maize breeding programs in the tropics. The heterotic grouping of the locally developed inbred lines in the present study was, therefore, based on using these CIMMYT established testers. Based on the assumption that SCA and heterosis of two inbred lines from different heterotic groups is greater than those from the same group, the heterotic orientation of the 10 inbred lines studied in this study will be useful for further breeding work in the national maize breeding program.
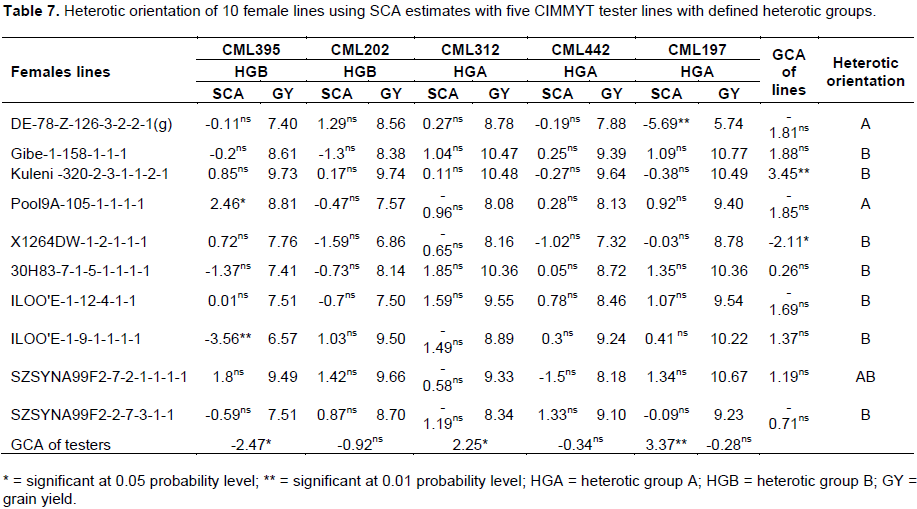
The inbred line L1 had no significant SCA effects for GY when crossed with T3, T4, T5 and T6, except with T7 where it had significant negative SCA effect (Table 7). This inbred line also gave the lowest GY (5.74 ton ha-1) when crossed to T7. This, therefore, indicated that L1 and T7 were highly likely to be in the same heterotic group. As a result, L1 was assigned to heterotic group A. Although L2 had no significant SCA effects when crossed with testers of both heterotic group A and B, it gave high heterosis when crossed with testers of heterotic group A (Table 7). Consequently, L2 was postulated to be in heterotic group B. L3 was assigned to heterotic group B because it gave relatively higher heterosis when crossed with testers from heterotic group A (10.48 ton ha-1 with T5 and 10.49 ton ha-1 with T7); and it had no significant SCA effects when crossed with testers from both heterotic groups.
L4 had no significant SCA effects when crossed to T4, T5, T6, and T7. The inbred line L4 when crossed with T3 exhibited the highest significant positive SCA effect (2.46**). This revealed that L4 and T3 could be in different heterotic groups and, thus, L4 was assigned to heterotic group A. The inbred lines L5, L6 and L7 had no significant SCA effects when crossed with testers from both heterotic groups. Nevertheless, due to the higher heterosis observed when L5, L6 and L7 were crossed to heterotic group A than to heterotic group B; these lines may be assigned to heterotic group B, but need further investigation. The inbred line L8 displayed highly significant negative SCA effect when crossed to T3 and showed no significant SCA effects with the rest testers. This indicated that L8 and T3 were in the same heterotic group and, thus, L8 was assigned to heterotic group B. There were no testers whose cross combination with L9 had significant SCA effects. However, all crosses of L9 with testers from both heterotic groups gave high heterosis (ranged from 8.18 tons ha-1 to 10.67 tons ha-1). Consequently, L9 was postulated to be in AB heterotic group. Despite the absence of cross combination that had significant SCA effect when crossed with L10, this inbred line had the highest negative SCA value when crossed to T3. In addition, this cross combination had the lowest mean grain yield (7.51 tons ha-1) compared to the cross of L10 with the rest of the testers indicating that these two inbred lines (L10 and T3) might be in the same heterotic group.
This study identified four new inbred lines and one tester inbred line (L1, L3, L4, L9 and T2) that were good combiners for reducing days to anthesis (DA) and day to silking (DS). Owing to the seasonal rainfall distribution and its inadequacy over much of the mid-altitude sub-humid agro-ecology of Ethiopia, rainfall is the overriding factor determining maize production in the potential maize producing areas of the country. The main rainy season being frequently not exceeding 3 to 4 months imposes limitations on the permissible delay in planting and early cessation before maturity ultimately causing substantial grain yield reductions. The inbred lines having significant negative GCA for DA and DS identified in the present study could, therefore, be used as parents for breeding maize for earliness in the mid–altitude sub-humid maize growing agro-ecology. Also, the three female inbred lines and three male inbred line testers (viz. L1, L9, L10, T2, T3 and T6) having significant negative GCA effects for PH and EH identified in the present study can also be used as parents for breeding maize for short stature to reduce the grain yield loss caused by lodging.
Likewise, the study identified 10 best cross combinations (viz. L2xT7, L9xT7, L8xT1, L3xT7, L3xT5, L2xT5, L6xT5, L6xT7, L3xT1 and L8xT7) that exceeded the best check (BH540 in this study) by over 10% in mean GY. Out of the 10 cross combinations, five had T7 as a common parent and T7 is a commercial line released as male parent of BH543. This inbred line has been used as a tester in the mid-altitude maize growing agro-ecology of Ethiopia having positive GCA as exemplified in the present study. However, it is no longer used as a tester due to its poor per se performance, especially in areas where turcicum leaf blight is important (data not shown). Consequently, the crosses that were selected based on their higher mean grain yield and have T7 as parent in their pedigrees have less value for commercial use. Of the remaining five crosses identified based on their heterosis over the best standard check, three (L3xT5, L2xT5, L6xT5) had T5 as a parent in their pedigrees. The high significant positive GCA effect for GY observed in the present study coupled with the good per se performance (data not shown) of this tester (T5), confirms that it can be used as parent in hybrid maize breeding for commercial release or for further breeding activities. Besides, the current study validated that T5 could remain to be used as a tester (in hetrotic group A) in mid- altitude maize breeding program until better tester is identified. Because L2, L3 and L6 were the elite inbred lines in their per se performance (data for per se performance not shown for all inbred lies), the crosses L3xT5, L2xT5, L6xT5 were considered the promising single cross hybrids for possible release after further verification of their performance across more environments, or for use as a single cross parents for predicting three-way cross hybrids. The inbred line T3 had the highest significant positive GCA effect for grain yield besides its good line per se performance. As a result, this inbred line was proposed to be used as a tester in heterotic group B. The rest two cross combinations (L3xT1 and L8xT1) selected based on their heterosis over the standard check having T1 as a common parent in their pedigrees, were also considered promising because both lines (L3 and L8) and the tester had good per se performance. In general, five promising single cross hybrids (L3xT5, L2xT5, L6xT5, T3xT1 and L8xT1) were identified based on their GY and their respective parental inbred lines’ per se performance.
As a further step, verifying the utility of the proposed new tester (L3) in early generation testing of lines needs to be conducted in order to give more information on the stability of its combining ability, and relate its performance to the previously used testers. In addition, further evaluation of the selected hybrids under more testing locations in order to get more information on Genotype x environment interactions needs to be done. The five single cross hybrids identified based on their GY and parents’ per se performance in this study, should be used to decide on either to use them for possible release or for predicting the performance of new three-way cross hybrids. The information obtained from this study on the heterotic pattern and combining ability of the inbred lines for different traits would be useful in planning hybrid maize breeding and predicting the hybrid performance.